
Abstract
Functional neuroimaging measures quantitative changes in neurophysiological parameters coupled to neuronal activity during observable behavior. These results have usually been interpreted by assuming that mental causation of behavior arises from the simultaneous actions of distinct psychological mechanisms or modules. However, reproducible localization of these modules in the brain using functional magnetic resonance imaging (MRI) and positron emission tomography (PET) imaging has been elusive other than for sensory systems. In this paper, we show that neuroenergetic studies using PET, calibrated functional magnetic resonance imaging (fMRI), 13C magnetic resonance spectroscopy, and electrical recordings do not support the standard approach, which identifies the location of mental modules from changes in brain activity. Of importance in reaching this conclusion is that changes in neuronal activities underlying the fMRI signal are many times smaller than the high ubiquitous, baseline neuronal activity, or energy in resting, awake humans. Furthermore, the incremental signal depends on the baseline activity contradicting theoretical assumptions about linearity and insertion of mental modules. To avoid these problems, while making use of these valuable results, we propose that neuroimaging should be used to identify observable brain activities that are necessary for a person's observable behavior rather than being used to seek hypothesized mental processes.
INTRODUCTION
Modern neuroimaging methods have significantly improved our understanding of brain function by allowing in vivo measurements of energetic and physiological parameters coupled to neuronal activity. Positron emission tomography (PET) has been used to measure the cerebral blood flow (CBF) and cerebral metabolic rates of oxygen and glucose consumption (CMRO2, CMRglc), whereas functional magnetic resonance imaging (fMRI) measures changes in blood oxygenation owing to changes in CBF and CMRO2. 13C magnetic resonance spectroscopy (MRS), in addition to evaluating the glucose and oxygen metabolic rates, has demonstrated that over 75% of total cortical energy consumption is coupled to glutamate and GABA (γ-amino butyric acid) neurotransmitter cycling. 1
At present, fMRI is the most widely used metabolic/physiologically based method for brain activity mapping in humans due to its high spatial resolution (down to 0.1 to 0.3mm in animal models and 1 to 3mm spatial resolution in human brain) and temporal resolution on the order of 1 second. In contrast, metabolic PET and MRS measurements provide spatial resolution on the order of 10mm and temporal resolution on the order of 2 to 60 minutes. Although recent functional neuroimaging methods provide quantitative or semi-quantitative measures of neurophysiological and neuroenergetic parameters, interpretations of these experiments largely are based on theories of mental processes derived from psychological studies of human behavior and cognition. 2 Here we show the impact of the empirical relationship found between brain energetics and neuronal activity on the interpretation of functional neuroimaging studies. We describe experimental disagreements with expectations of the prevailing cognitive neuroscience model of functional imaging, and offer experimental support for an alternative model of brain function described in a recent book by one of us. 3 The new imaging methods have shown that the neuronal activities underlying the fMRI signal are many times smaller than the high, ubiquitous baseline neuronal activity found in resting and awake humans and animals. Results show that the magnitudes of stimulus-evoked fMRI signals depend on the level of baseline neuronal activity. These findings challenge the traditional interpretation that the differenced blood oxygenation level -dependent (BOLD) signal maps the neuronal activity that supports hypothesized mental processes. Instead, we propose that neuroimaging studies should be used to identify brain activities that are necessary for a person's observable behavior. Necessary means that the person cannot perform the behavior without the activity (e.g., indicating to an investigator that he recognizes a projected face). In place of assuming innate modules that perform brain functions like memory, attention or cognition, we propose to identify brain activities that provide necessary support for the person's observed behavior. By not introducing unreliable psychological interpretations the reliable neurophysiological imaging results can support subsequent physical experiments.
Physiological studies dating back to the 1950 s by Kety and colleagues 4 showed that the active or resting brain, although only ∼2% of body weight accounts for ∼20% of whole-body glucose and oxygen consumption at rest. 5 Despite this disproportionate energy requirement, it was largely believed, before the 1980 s, that the energies devoted to brain function were small, 6 and that neuronal activity was essentially negligible at rest compared with the increase in activity measured during sensory stimulation or cognition 7 (we note an early exception to this view from Ingvar and Lassen9,10 who found very small changes, including decreases, in blood flow during stimulations and tasks 8 ). However, as described below, and more thoroughly in several reviews,11–19 a range of neuroenergetic studies over the past 16 years have shown that the majority of energy consumed in the resting awake mammalian cerebral cortex directly supports synaptic function. Resting levels of neuronal activity are one to two orders of magnitude greater than activity changes induced by sensory stimulation and externally driven cognitive tasks. This review discusses the implications of this high baseline activity on the standard interpretation of fMRI and describes our proposed alternative model of brain function.
RESTING AWAKE BRAIN ENERGY PRODUCTION PRIMARILY SUPPORTS NEURONAL ACTIVITY
Over 90% of neurons and synapses in the cerebral cortex are either GABAergic or glutamatergic.1,20 Figure 1 is a schematic of oxidative energy metabolism in a glutamatergic neuron and an astroglial cell along with the neurotransmitter cycle that couples neuronal and glial metabolism. Neuronal glutamate released via neurotransmission from presynaptic neurons is taken up by glial cells and converted to glutamine by glutamine synthetase, but glutamine may be also be synthesized de novo with pyruvate carboxylase. Glutamine synthesized by these two enzymes can either be involved in glutamate oxidation or help maintain nitrogen balance.21–23 13C MRS has measured the flow of 13C-labeled substrates (e.g., glucose, acetate) into neuronal pools of glutamate and GABA and glial pools of glutamine. In conjunction with metabolic modeling these results have been used to determine the rates of glucose oxidation in GABAergic astroglia and glutamatergic neurons (i.e., CMRglc(ox),N), as well as the rates of the glutamate/glutamine and GABA/glutamine cycles whose sum equals the total cycling, Vcycle. The rate of glucose oxidation is evaluated from the tricarboxylic acid (TCA) cycle flux.1,20
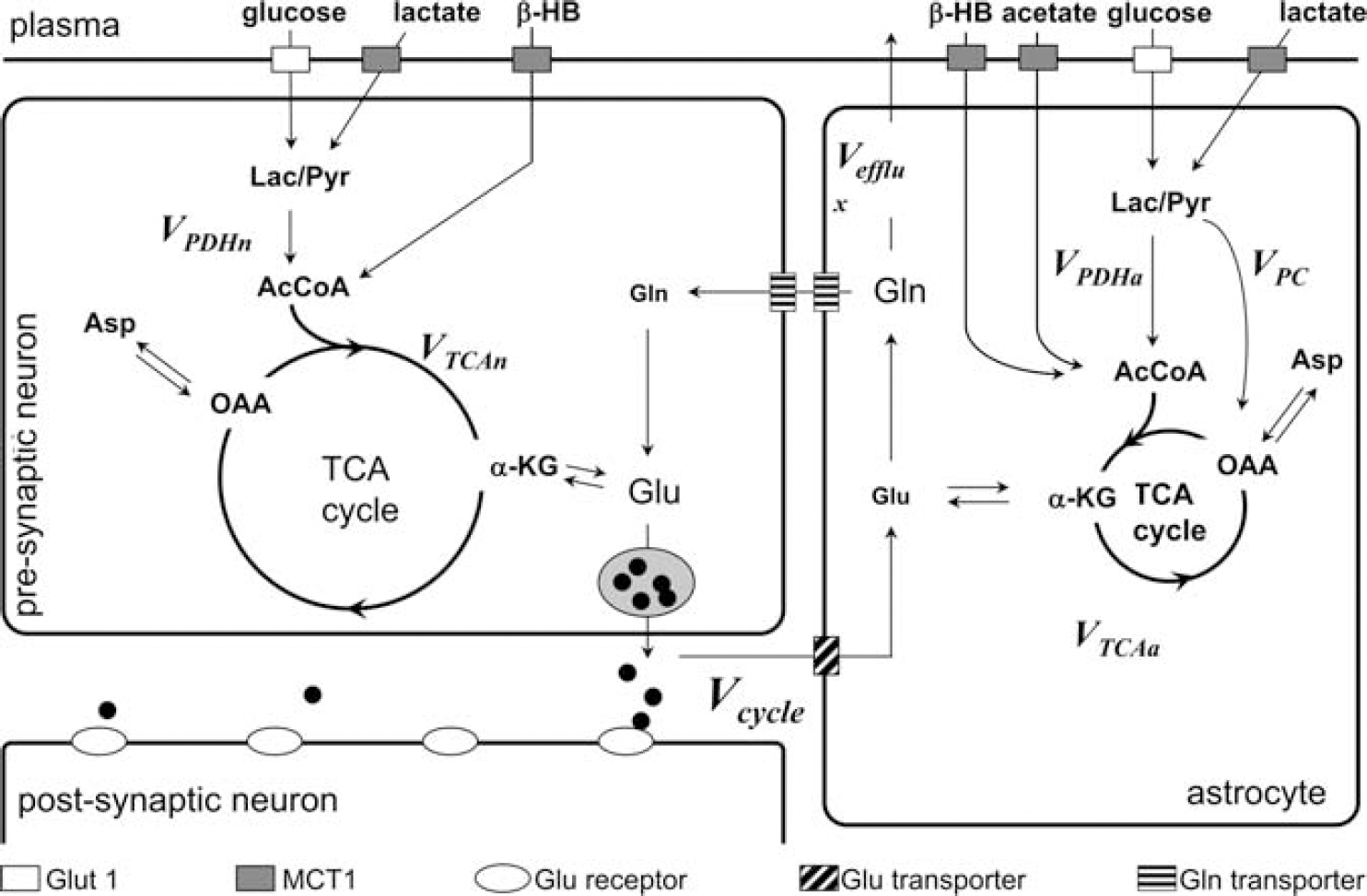
Metabolic pathways within glutamatergic neurons and surrounding astroglial cells. The diagram shows the glutamate (Glu)/ glutamine (Gln) cycle (Vcycle) between neurons and astroglial cells. Glucose and lactate enter the glial (VTCAa) and neuronal (VTCAn) tricarboxylic acid (TCA) cycles via pyruvate dehydrogenase (Vpdh), β-hydroxybutyrate (β-HB) is directly incorporated into the neuronal and astroglial TCA cycles, and acetate is near exclusively incorporated into the glial TCA cycle. Neuronal Glu that is released via neurotransmission is taken up by astroglial cells and converted by Gln synthetase to Gln at a rate equal to the Vcycle. In addition to neurotransmitter cycling, Gln may be synthesized de novo starting with the pyruvate carboxylase (PC) reaction (VPC). Gln synthesized via PC can replace neurotransmitter Glu oxidized in the astrocyte or elsewhere (and be recycled back to the neuron) or leave the brain (Vefflux) to remove ammonia and maintain the nitrogen balance.21–22 To measure the rates of these pathways, 13C-labeled substrates are used and the flow of 13C isotope into Glu and Gln is measured using 13C magnetic resonance spectroscopy (MRS). For detailed descriptions of how these pathways are tracked using 13C MRS, and how isotopically labeled substrates and rates are calculated by metabolic modeling, see details of citations within.1 AcCoA, acetyl-CoA; Asp, aspartate; Glut 1, glucose transporter 1; a-KG, a-ketoglutarate; Lac, lactate; MCT1, monocarboxylate transporter 1; OAA, oxaloacetate; Pyr, pyruvate. Reproduced with permission from Rothman et al. 1
13C Magnetic resonance spectroscopy measurements of the relationship between the glutamate/glutamine cycle and neuronal glucose oxidation
To determine the energetic cost of brain function13C MRS studies have been performed in rats over a wide range of activities from isoelectric pentobarbital anesthesia under which there is no cortical signaling to awake states with high levels of cortical signaling. The original results by Sibson et al 24 have been reproduced in several laboratories,25–38 demonstrating a tight correlation between Vcycle and CMRglc(ox),N (Figure 2; see Supplementary Table 1 for both rat24–39 and human23,40–52 results). This neurometabolic coupling in the rat somatosensory cortex spanning awake to deeply anesthetized conditions has several consequences. First, it demonstrates a linear relationship between neuronal activity, as measured by glutamatergic flux, and the rate of glucose oxidation. Second at isoelectricity when Vcycle falls to zero the remaining neuronal energy consumption falls to ∼15% of the total neuronal energy consumption in the awake state. Therefore, in the awake resting rodent brain ∼85% of the neuronal energy demand is devoted to events associated with neuronal signaling. Third the data show a stoichiometric relation wherein one equivalent of glucose oxidized is accompanied by one equivalent of neurotransmitter glutamate released and recycled. A recent in vivo 1H MRS study combined with electroencephalography (EEG) recordings in the rat cortex also showed a linear coupling between EEG activity and glucose consumption with a finite intercept during cortical isoelectricity. 53 Small increases in both glutamate and lactate have been observed with 1H MRS during sensory and motor stimuli in humans,54–56 as well as increases in Vcycle.56–58 Recently, a top-down energy budget for the rat cortex, relating layer-specific neuronal firing rates from awake to isoelectric conditions to glucose metabolism measured by 2-deoxyglucose autoradiography, suggests that linear coupling between neuronal firing rate and glucose metabolism in neurons has a finite intercept during cortical isoelectricity, 59 very similar to the 13C MRS results shown in Figure 2.
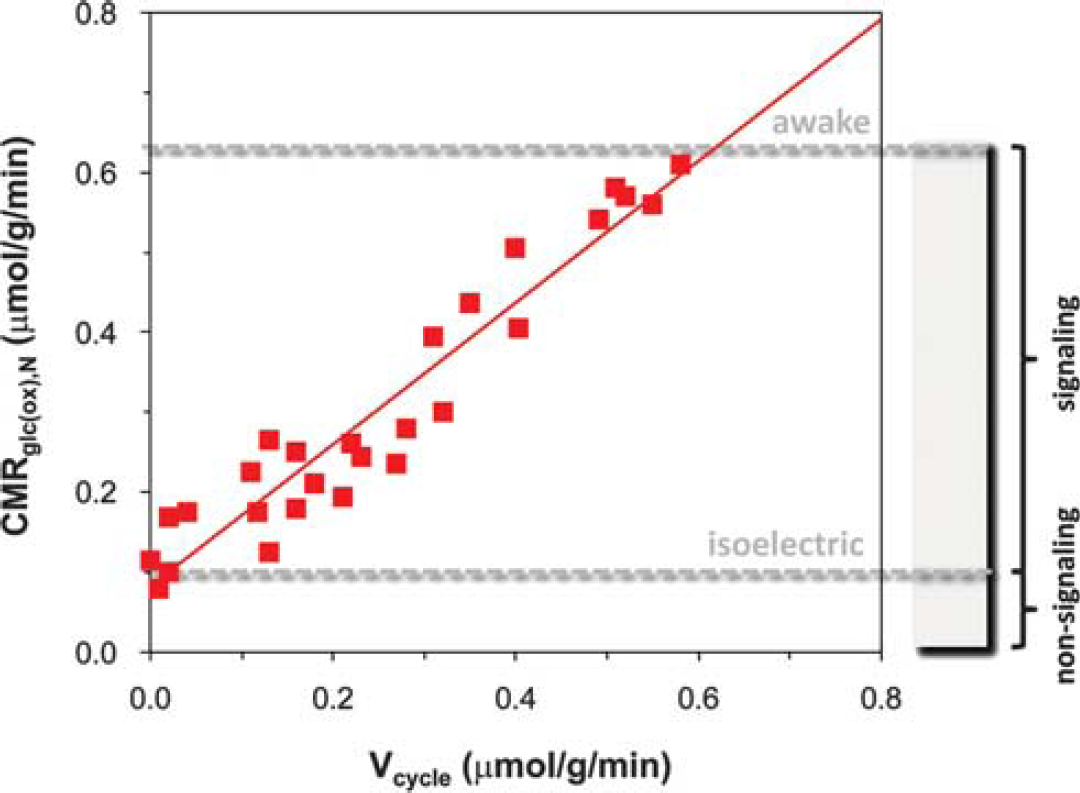
Relationship between neuronal activity (horizontal axis) and glucose metabolism (vertical axis) measured by magnetic resonance spectroscopy (MRS) in rat brain. Rates of glutamate neurotransmitter cycling (Vcycle, horizontal axis) and glucose oxidation in neurons (CMRglc(ox),N, vertical axis) measured by 13C MRS in rat brain. The 24 different data points represent a variety of behavioral states—different levels of anesthesia, sleep, seizure, awake, etc.,—from 14 independent studies.24–39 These 13C MRS results suggest that in the resting awake state ∼85% of neuronal energy consumption supports signaling events associated with neuronal activity, whereas ∼15% of neuronal energy consumption supports non-signaling functions. The red line is the best-fit linear regression of the rat data (i.e., CMRglc(ox),N=0.9 Vcycle + 0.1, R2=0.9). Modified from Hyder et al. 19
A similar relationship has been found between the GABAergic TCA cycle and GABA/glutamine cycling by 13C MRS in rat brain. 60 The rates of the TCA cycle and neurotransmitter cycling in both glutamatergic and GABAergic neurons increase in parallel as opposed to the common misconception that GABAergic activity increase leads to a decrease in glutamatergic activity (e.g., see first figure in Wieronska et al 61 ). These 13C MRS findings are consistent with recent electrophysiological recordings by McCormick and colleagues 62 that suggest balanced excitation and inhibition across a range of activity levels, where changes in total activity seem to be determined by small shifts in the excitatory/inhibitory balance. In the resting awake rodent and human cerebral cortex 13C MRS studies have shown that astroglia account for between 15 and 20% of total energy consumption (see Hyder et al 20 ). The slope of glial oxidative energy consumption versus neuronal activity, as measured by neurotransmitter cycling or by electrical recording, is several-fold lower than for glutamatergic and GABAergic neurons.25,26,42,46–48,63 We note that the measured 13C MRS values of glial energy consumption are several-fold higher than calculations from bottom-up energy budget models. 64 However, a recent paper has suggested that the low glial energy consumption calculated was likely due to the use of conductivity values for astrocytes from slices which are several fold lower than in vivo measurements. 59 With the non-functional energetics of both neurons and glia subtracted ∼70% to 80% of cortical gray matter energy consumption supports functional neuronal processes in cortical gray matter in the awake resting state.
Multiunit recordings of neuronal activity versus changes in energy consumption measured by calibrated functional magnetic resonance imaging
The relationship between energetics and function has also been evaluated by comparing absolute changes in energy metabolism using calibrated fMRI with measurements of ensemble electrical activity by multiunit or single-cell recording of an ensemble of neurons.65–67 These measurements compared sensory-evoked changes in regional energy metabolism and electrical signaling at different levels of anesthesia. Total regional electrical signaling was quantitated by measuring the firing rate of a representative ensemble of neurons within the voxels where the calibrated fMRI measurements were made.
Calibrated fMRI refers to methods with which quantitative changes in CMRO2 can be determined from multimodal fMRI measurements of BOLD signal (S), blood flow (CBF), and blood volume. These methods take advantage of the following biophysical model originally proposed by Ogawa and extended by others,68–72
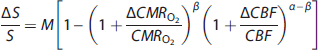
where α describes the cerebral blood volume-CBF coupling and is assumed to be 0.4 in human studies based on PET primate studies with hypercapnia challenge, 73 but is directly measurable in animals74,75 and also now in humans76,77 using intravascular MRI contrast agents based on superparamagnetic iron oxide nanoparticles; β describes the CMRO2-CBF coupling and is assumed to be 1.5 in human studies based on simulations of BOLD effect at 1.5T 78 but it may approach unity at higher magnetic field strengths; 79 and M represents the baseline BOLD signal and can be either modeled from hypercapnia challenged experimental data by examining the BOLD-CBF coupling under the assumption that CMRO2 does not change during the gas exposure 71 or directly measured from transverse relaxation measurements (of gradient- and spin-echo MRI) under well shimmed conditions.79,80
Values of ΔCMRO2/CMRO2 determined by calibrated fMRI65–67,81,82 were compared with the firing rates (v) from multiunit recordings of a representative neuronal ensemble, demonstrating a quantitative relationship between changes in energy, as ΔCMRO2, with changes in representative firing, as ΔV (Figure 3). The neuronal firing rates for a given condition (i.e., resting or stimulated) were represented in a histogram of the number of neurons in the somatosensory cortex at each neuron's average firing frequency.65,67 As the anesthetic dose increased (e.g., light versus deep α-chloralose), the histograms shifted to lower firing rate values and integrating over the histogram to determine average firing frequency showed a proportional decrease in v and CMRO2 (Figure 3A). Moreover, on sensory stimulation in each anesthetized state the histograms shifted to higher firing rates showing that both CMRO2 and v increased consistently (Figure 3B). In agreement with the earlier 13C MRS results (Figure 2), these calibrated fMRI and electrophysiology results (Figure 3) demonstrated a linear relationship between neuronal activities, as measured by multiunit firing of pyramidal neurons, and their oxidative demand showing that CMRO2 changes derived from calibrated fMRI are closely associated with glutamatergic function.72,83 Owing to the heterogeneity of neuronal responses within the volume resolution of fMRI it is important that a representative histogram of the entire neuronal response be obtained to relate it to energy consumption. For example, in the studies described above ∼60% of the population showed an increase in v, whereas other neurons either decreased or showed no change in firing rate. Measurement of a small number of neurons could easily miss the relationship between ensemble firing and energy consumption in a region.
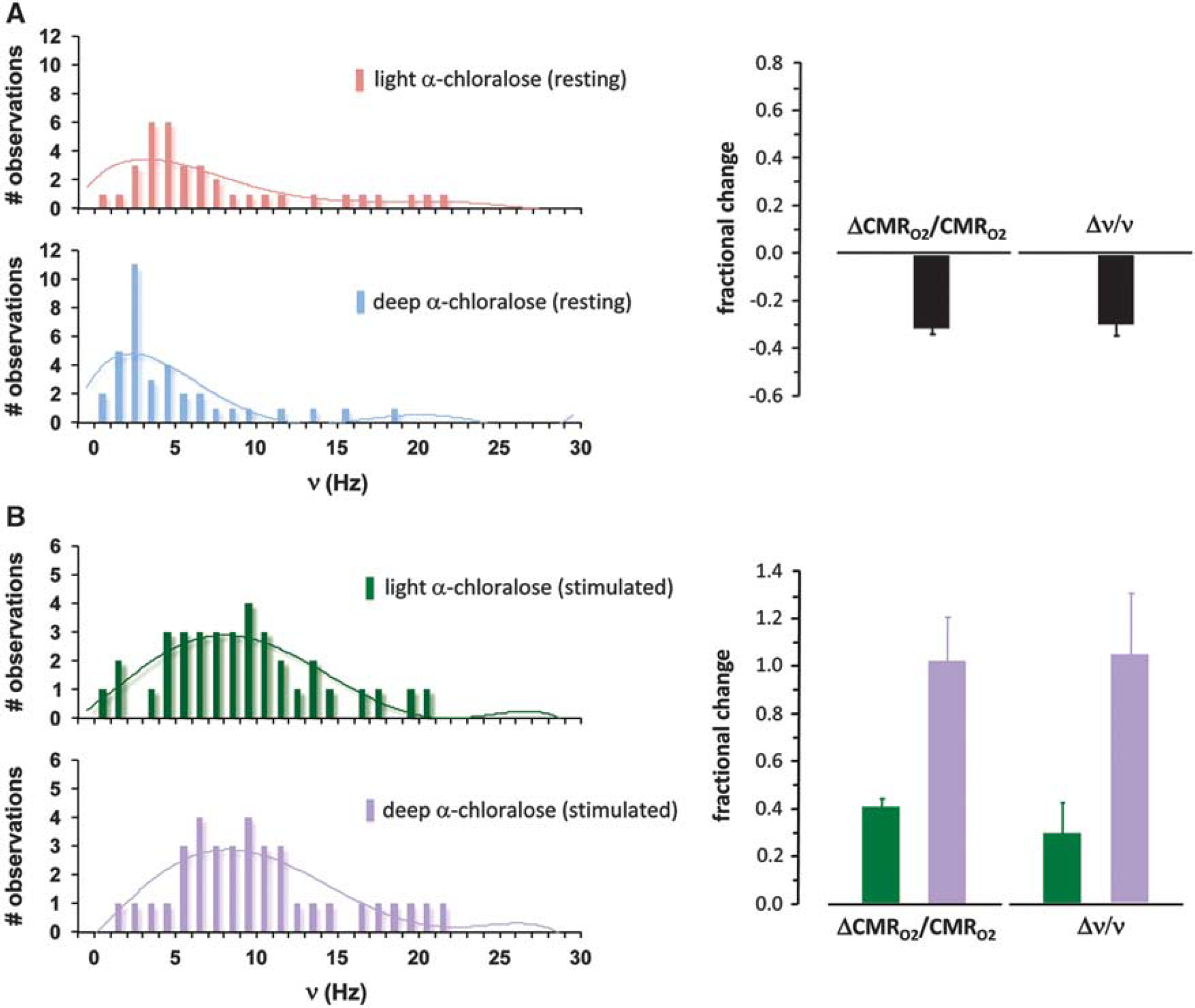
Relationship between fractional changes in localized oxidative energy demand (ΔCMRO2/CMRO2) and firing rate of an ensemble of pyramidal neurons (Δv/v) in the rat somatosensory cortex, measured respectively by calibrated functional magnetic resonance imaging fMRI and electrophysiology.
65
Histograms, which represent firing rates of a population of neurons, are shown for (
The relationship between measurements of BOLD and electrical activity has been controversial with different experiments finding variable correlations of BOLD signals with either multiunit activity or local field potential.65,84,85 We will not address this controversy here except to point out that the regional energy consumption derived from calibrated fMRI has consistently tracked well with both neurotransmitter cycling and multiunit activity-type electrical activity.65–67,86 The cases where the non-calibrated BOLD signal, which depends on the relative changes in CBF, CMRO2, and cerebral blood volume as indicated by equation (1), correlates better with local field potential or multiunit activity may be due to different spatial as well as temporal couplings between CBF and cerebral blood volume with neuronal activity.81,82,87
High energy demand for resting state electrical activity in the awake human cerebral cortex
As shown in Figure 4A, the ratio of Vcycle/CMRglc(ox)N measured in awake human occipital cortex is highly consistent with the findings in rodent somatosensory cortex, supporting a similar high baseline level of neuronal activity. The slightly lower value in the human studies is probably due to the measurements sampling a higher fraction of white matter (where relatively little glutamate neurotransmitter cycling takes place) due to the higher percentage of white matter in the human brain. 88 To assess whether there is a linear relationship between energy metabolism and neuronal activity as in the rat, a meta-analysis was performed of EEG and PET data from humans at various levels of activity, 19 under conditions ranging from awake89–92 to non-rapid eye movement sleep91–94 to different anesthetized states (i.e., halothane,95–97 sevoflurane,96,98,99 and propofol91,98,100,101) and to different vegetative states (i.e., acute102,103 and persistent104,105). Figure 4B shows a linear relationship between total EEG power estimated from the bispectral index (BIS) and CMRglc measured by fluorodeoxyglucose PET for the different conditions (Supplementary Table 2). The BIS versus CMRglc plot in Figure 4B has an intercept at ∼20% of the resting awake CMRglc value (i.e., when BIS approaches zero), showing that in the resting awake human there is a similar high fraction of energy consumption supporting signaling as in the rat brain (Figure 2).
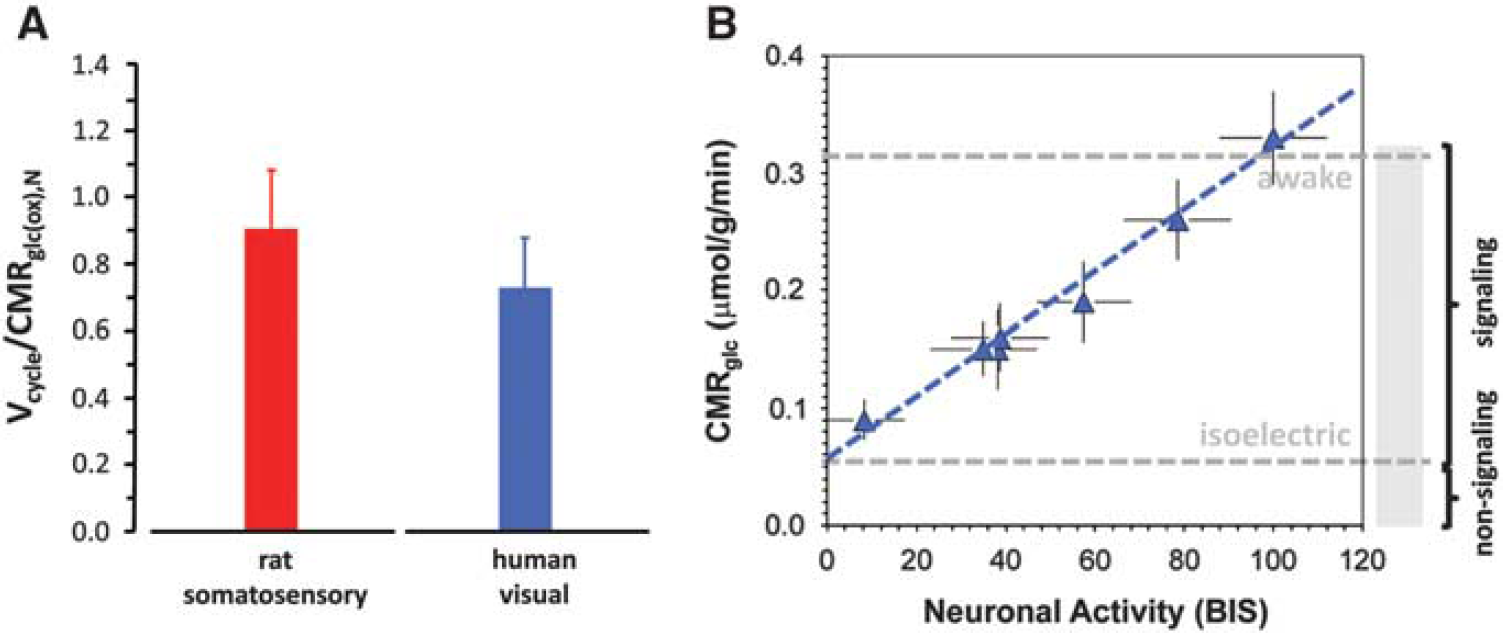
Summary of 13C magnetic resonance spectroscopy, positron emission tomography (PET) in μmol/g/min, and electroencephalogrphy (EEG) results from the human visual cortex. (
To examine whether the high resting state activity is present throughout the human cerebral cortex, a meta-analysis 19 of PET data, acquired under quiet and awake experimental settings, analyzed regional distributions of CMRO2 and CMRglc. 106–110 The results, presented as histograms of average CMRO2 or CMRglc for different anatomical brain regions, were examined to determine whether any anatomical regions or proposed brain networks, including the default mode network, 110 have different energy demands. Figure 5 shows part of the results of this meta-analysis, where the ratio of CMRO2/CMRglc was represented by the oxygento- glucose index. High oxygen-to-glucose index values are present throughout the cerebral cortex as well as in proposed resting state fMRI-based networks with variations rarely above 10% of the mean, with a similar uniformity of the component CMRO2 and CMRglc values. We anticipate that PET data when corrected for white matter within voxels would give an even smaller variation in metabolic demand. 19
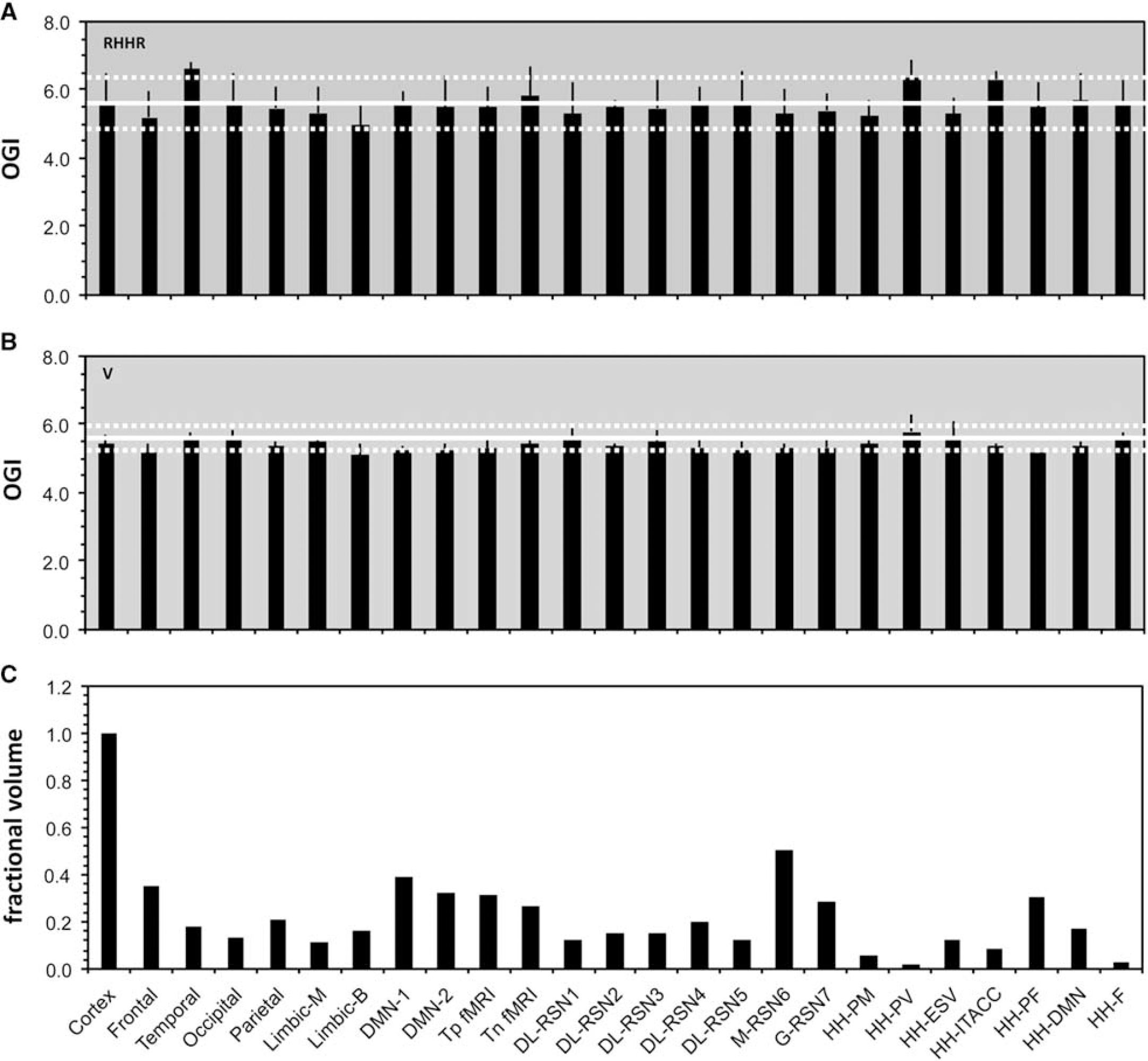
Variation of glucose oxidation across brain networks for the resting awake human brain, given by the ratio of rates for oxygen (CMRO2) and glucose (CMRglc) demand or the oxygen-to-glucose index (i.e., OGI =CMRO2/CMRglc). Oxygen-to-glucose index (OGI) comparison for (
THE RELATIVE MAGNITUDES OF STIMULATED AND RESTING STATE NEURONAL ACTIVITY
The relative magnitudes of increments in neuronal activities during a task relative to the baseline activity can be obtained from calibrated fMRI and PET studies, which have calculated ΔCMRO2 (or ΔCMRglc) under a wide range of cognitive tasks and sensory stimulations. The largest changes have been found in the primary visual and motor cortex with a maximum of 10% to 15% by PET111–113 and up to 30% with calibrated fMRI.83,114,115 This difference between PET and MRI is probably due to larger voxels with greater partial volume effects in the PET data. Much smaller increments are seen in secondary areas associated with cognitive activity, such as the frontal lobe, temporal lobe, and hippocampus where changes are at most 10% and more often 1% or less. Thus, in all cases the baseline activity, as estimated from the total energy consumption, is much larger than the induced activity.
The impact of baseline activity on the magnitude of functional magnetic resonance imaging signals
The conventional view of the baseline activity is that it does not interact with the activity induced by external stimuli. 116 However, the impact of baseline activity on the magnitude of induced activity during sensory stimulation was recently highlighted in a commentary by Hyder et al. 117 Figure 6 summarizes typical results from one of these studies,65,118 which shows that the stimulus evoked increase in energy demand (e.g., ΔCMRO2 measured by calibrated fMRI, 17O MRS, or PET) depends on the depth of anesthesia—the lower the baseline activity the larger the magnitude of the induced activity. On stimulation the total activity reached was, within experimental errors, close to equal.
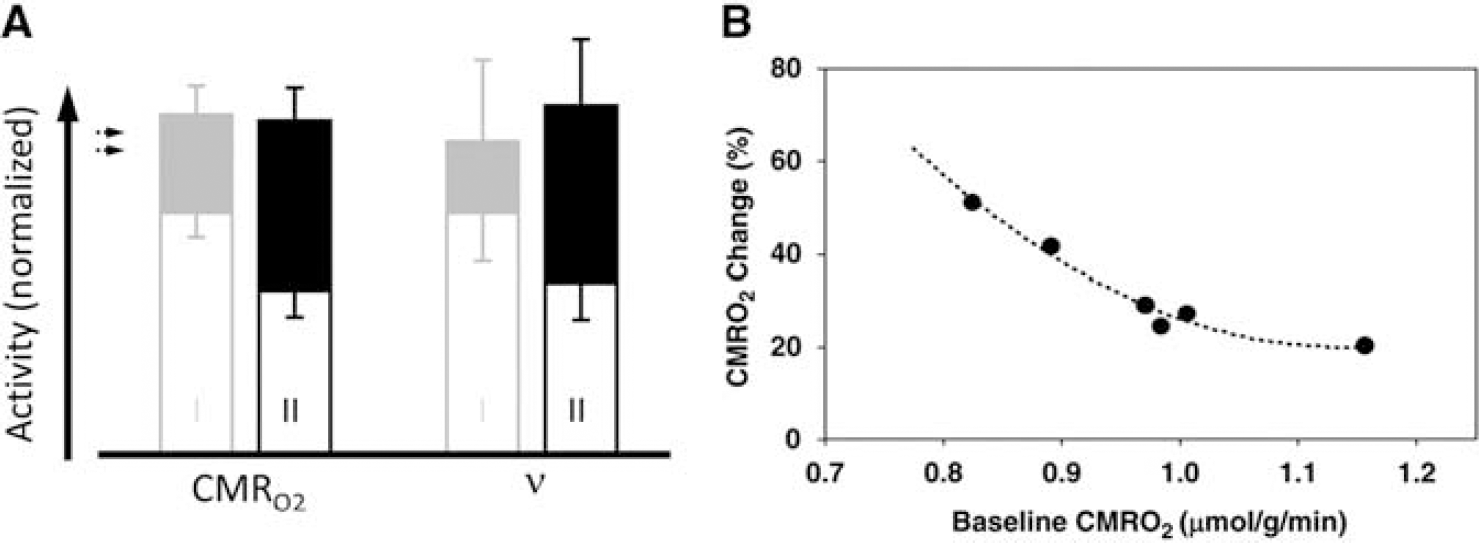
Relationship between the magnitudes of stimulated and resting activity, represented by measurements of oxidative energy demand (cerebal metabolic rate of oxygen consumption (CMRO2)) and firing rate of an ensemble of pyramidal neurons (v). (
Measuring how the baseline activity also influences conventional electrophysiological parameters Xu and colleagues 119 exposed a rat to olfactory stimuli at different levels of baseline activity induced by anesthesia and created a histogram of measured neuronal spike rates. The total activity and distribution of firing during olfactory stimulation were the same independent of the starting baseline state (i.e., similar to Figure 3B). As the differenced activities from the two baseline states were not the same, these results contradict the traditional interpretation that a fixed amount of induced neuronal activity supports sensory processing independent of baseline activity.13,16 The dependence of evoked responses on baseline activity has been found for a variety of behavioral states and neuroimaging techniques (e.g., fMRI, optical imaging, electrophysiology).67,120–125
IMPLICATIONS OF THE HIGH BASELINE ACTIVITY ON THE INTERPRETATION OF FUNCTIONAL MAGNETIC RESONANCE IMAGING
Given the prevailing view of the brain, as primarily an interactive organ that is active only when stimulated, consequences of the baseline activity are often disregarded.116,126 If the baseline activity were negligible the differenced fMRI signal would be a quantitative measure of the neuronal activity induced by a stimulus or task (Figure 7A), However, in the actual case of high baseline activity additional assumptions are required to assign incremental neuronal activity (Figure 7B). Here we describe how the standard assumptions used in psychological models of fMRI studies lead to misinterpretations when the baseline activity is ignored.
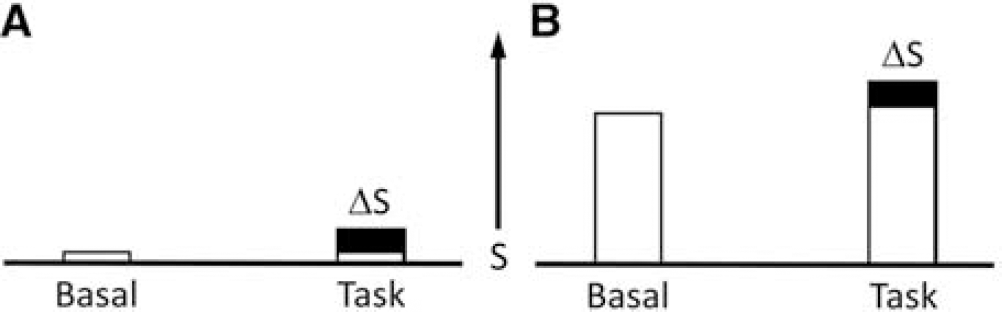
Schematic representation of the signal (S), quantitated as total energy devoted to neuronal activity, obtained in functional imaging differencing experiments. Positron emission tomography measurements of cerebral blood flow, cerebral metabolic rate of glucose consumption, and cerebral metabolic rate of oxygen consumption directly measure S and S+ΔS under both basal (i.e., no stimuli) and task (i.e., stimuli) conditions, although usually only the difference ΔS is used to localize and quantitate the task activation. The conventional analysis of functional magnetic resonance imaging is based on the same differencing approach, where the focus is mainly ΔS, either by differencing or by correlation. (
Definitions of terms
In the following sections of the paper which discuss the standard cognitive neuroscience interpretation of functional imaging and our alternate interpretation, we have used the terms brain activity, mental processes, mental modules (abbreviated to module) and behavior. We provide here definitions of these terms. Brain activity is any measure of neuronal signaling or communication within a brain region. Examples are neuronal firing rate, pre- and postsynaptic potentials, and neurotransmitter release. As described above, a large fraction of brain energy metabolism is directly coupled to the rate of neurotransmitter release and neuronal signaling (firing rate, synaptic potentials), so that fMRI and CMRglc PET measurements can be directly related to brain activity. A mental process is a brain state that corresponds to and gives rise to a behavior such as memorizing a list of words. Mental processes are usually given common names describing the behavior (or internal states/feelings) such as memory, emotion, speech, and craving. These mental processes are further assumed to consist of the simultaneous activity of a variety of distinct, modules. Modules are normally defined by theories from cognitive psychology, which studies the brain by treating it as an information processing system such as a computer. For example the mental process of working memory, in which a subject is asked to remember information and perform a mental manipulation of the information (e.g., being briefly shown a number and then writing it down backwards), has been decomposed into a range of modules. An early example is from Baddeley and Hitch 127 in which working memory was broken into multiple modules including a central executive, visuospatial scratch pad, and phonological loop and store. Modern theories have greatly extended the number of modules involved in this process but have largely not questioned that an orthogonal decomposition of the components can to some degree be performed. 128 A much more detailed analysis of modules and a critical look at their ontological validity has been presented by Fodor. 129 We define behavior operationally as the observable actions of a person. For example, in a fMRI study of memory in which a subject is asked to memorize a list of words the behavior would be the subject indicating the words that they remember. As discussed below, in our interpretation an internal mental process of ‘memory’ is not assumed or implied by this definition but the brain processes necessary for the person to perform the behavior that defines that process can be inferred from brain activities during the observed behavior.
The standard relation between signal changes and mental processes/modules in functional imaging
The conventional interpretation of functional imaging was largely developed by the pioneering studies of Posner and Raichle. 130 In an early study, using H2 15O PET, scans were performed on subjects at rest and compared with scans obtained during visual presentation of False Fonts, Letter Strings, Pseudo-words or Meaningful Words. 131 The most significant results were the unique incremental changes in the left hemisphere from Pseudo-words and Meaningful Word presentation relative to Letter Strings and False Fonts. It was assumed, based on the theory of insertion from cognitive neuroscience 131 that different modules were independently activated by the different stimuli. The theory of insertion assumes that to a first-order modules are independent so that a new module could be activated without modifying the existing activities.131 In the present example, the unique left hemisphere response was assigned to a cognitive module for semantic understanding not present in other tasks, while the neuronal activity associated with the visual processing modules were assumed to be the same for all presentations. This interpretation was generalized and made quantitative by Frackowiak et al132 who introduced statistical parametric mapping as a way to assess the statistical validity of signal changes in functional imaging studies. In their analysis it was assumed that mental processes (and their underlying modules) are independent, and within each image voxel there is a linear relation between the induced neuronal activity (supporting one or more modules) and the size of the imaging difference signal.
Using the Posner and Raichle study described above as an example, and following the statistical parametric mapping interpretation, the following relations (shown in equations (2) to (4) can be written for the Meaningful Word (Task 2) and the Letter String (Task 1) presentations. Mv is the mental (i.e., neuronal) activity supporting visual processing, Mc is the neuronal activity supporting semantic understanding of words, S1 and S2 are the signal magnitudes for Tasks 1 and 2. The term Sb is the baseline signal in the voxel, which generally was assumed not to be related to neuronal activity. In PET the baseline signal would be the CBF, CMRglc, or CMRO2, measured in the resting state. The increase in image signal within a voxel is related to the underlying neuronal activity supporting the mental processes by a constant k.
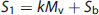
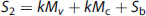
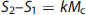
Provided the assumption of no neuronal activity in the baseline state is correct, the subtraction of S2 and S1 is seen as isolating the signal (proportional to neuronal activity) derived from the hypothesized modules supporting cognitive activity (kMc) (equation 4). In the general case a summation can be made describing the relationship of the increment in signal within each voxel to any number of underlying mental modules (Mj)
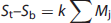
Ambiguity in assigning neuronal activity to mental processes/ modules caused by the high resting state neuronal activity
As described above, a key assumption in the standard model is that the neuronal activity (or equivalently the change in image signal) that supports a mental process can be determined by the difference signal. This assumption would be trivially true if neuronal activity in the resting state were negligible, which was implicitly assumed until the late 1990 s, based on the standard model of the brain as containing modules devoted to specific externally driven computational processes. However, as first pointed out by Shulman and Rothman 11 the experimental findings of high human and rodent baseline neuronal activity called into question the assumption that the magnitude of the neuronal activity supporting a mental process could be obtained from the difference in signal between tasks or between task and rest. As an example they gave two hypothetical tasks, one that increased energy consumption in a voxel by 2% and in a second by 1%. In the standard interpretation the first task (and the mental processes/modules supported by it) required twice the neuronal activity in the voxel as the second. However that conclusion depends on the assumption that the large baseline activity is not involved in supporting the task. If instead the total, as opposed to just the incremental, activity was used during the task then the conclusion would need to be that the first task required ∼1% more energy (and by inference neuronal activity) than the second (Figure 7). Another consequence of the high baseline activity discussed in the paper was that the absence of change in a voxel between conditions cannot be interpreted by itself as evidence that the neurons in the voxel are not supporting performance of the task (or an underlying mental process) as an equally good interpretation is that the total neuronal activity supporting the task is coincidentally the same in both conditions, within the signal-to-noise, leading to a false negative. As described above, this prediction has been supported by findings of large increases in areas of activation (and deactivation) when imaging sensitivity was improved.
The paradox of negative functional magnetic resonance imaging signals and explanation based on the high baseline activity
A paradox created by the standard model's assumption that mental processes were supported by increases in neuronal activity was first noticed by G.L. Shulman et al 133 who reported negative signal changes in some brain regions. These negative changes were originally missed because it was assumed in the data analysis that they would not exist due to an implicit assumption of negligible baseline neuronal activity. This paradox was resolved by Shulman and Rothman 11 who proposed that in these regions the neuronal activity during a task was simply less than the high baseline neuronal activity (and associated energetics) in the absence of stimulation.
Impact of resting state activity on the assignment of the incremental neuronal activity to a mental process
Several investigators, e.g., Morcom and Fletcher,
126
have argued, on theoretical psychological grounds, that the high baseline neuronal activity does not impact the interpretation of fMRI because it will be canceled by differencing. The proposal of Morcom and Fletcher, as applied to the comparison of the signal in a task (St) and baseline (Sb) state can be expressed as follows for each voxel
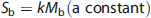
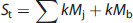
LINKING NEURONAL ACTIVITY TO BEHAVIOR WITHOUT ASSUMING MENTAL PROCESSES: AN ALTERNATE APPROACH FOR INTERPRETING NEUROIMAGING
As described above, the large resting state baseline activity creates a substantial ambiguity in the assignment of a change in neuronal activity measured by fMRI (or PET) to a hypothesized mental process/module. Based on the results of an fMRI experiment which failed to find a unique location for working memory in 1994, 134 Shulman 135 proposed that rather than the problem being methodological perhaps fMRI was not confirming psychological theories because the theories themselves were not describing how the brain works. It was suggested that a better way to study the brain is to let the experimental results drive the development of theoretical explanations. In 2009 in a paper 14 and 2013 in a book, 3 these ideas were extended into an alternate paradigm for how the brain supports behavior. In studies based on this newer paradigm brain activity is directly related to measured behavior as opposed to the standard approach of assuming intervening mental processes.
Schematically the standard model, proposed in Cognitive Neuroscience, can be described as
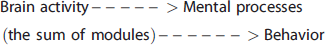
Alternatively, we propose that to interpret brain activity by functional imaging it is more useful to directly look at the relation between brain activity and behavior

In the first model, the experimental goal depends on knowing the nature of the presumed modules and then measuring how they are activated and combined into a mental process that supports an observed behavior. In this view, once one has identified how these modules interact during the behavior one has understood how they have been combined to cause the behavior which is ‘tantamount to a theory of the structure of the mind’. 129
In the second model (equation (9)), hypothesized mental processes are not included in the pathway from necessary brain activities to behaviors. Behavior can be identified to an acceptably objective degree and brain activities can be identified that are necessary for the individual to recognize stimuli and to perform the indicated tasks. In this approach, brain activities follow, support and are necessary for the person's behavior, reflecting the person's experience, but they do not cause the behavior. Furthermore, it is not assumed, as in the standard model, that brain activities can be separated into mental processes that correspond to everyday descriptions of behaviors/internal states such as memory, intent, and emotion.
The standard Cognitive Neuroscience model attempts to resemble the reasoning of classical physics where given the state of the object (in this case an activated mental process or combination of modules) there are rules from which the effect (a behavior) will follow. However this application differs significantly from classical physics where both the identification of phenomena and the rules for predicting their future have been well established experimentally. Rather the hypothesized modules are obtained largely from theories developed to explain human behavior from a psychological or information theory perspective. 129 In our alternate proposal, resembling the usual reasoning in biology, a phenomenon, the behavior, is identified and is understood by its properties, including, in this case, the neuronal mechanisms that are necessary for it to occur in the human. With accumulating measurements of the properties, our understanding of the phenomenon grows, eventually leading to an understanding which may completely differ from the mental processes assumed today.
The experimental background for these models of brain functioning can be found in studies of the visual systems of animal models, which empirically assigned specific visual processes to anatomic regions in the visual cortex. Functions were assigned by exposing animals to specific visual stimuli (e.g., rotating pinwheels, colors, checker boards), and identifying localized non-overlapping brain regions that responded through either an increase in their neuronal firing rate or the glucose metabolism. 136 Over time, a lexicon was developed for mapping specific columns to specific components of a visual presentation (e.g., edges, color, rotation), thereby identifying brain regions such as specialized columns as modules of visual activity. These studies are often cited as the basis for higher order cognitive modules, as in Posner and Raichle, 131 but this extension suffers because although brain responses to sensory stimuli have a strong empirical basis, the responses to higher-order stimuli such as would be engaged in a cognitive task do not. In today's fMRI studies when using the standard model to look at higher order behaviors (e.g., remembering, intending, or being conscious) underlying mental processes and modules are assumed to have been well established by prior psychological experiments and the role of the brain imager is merely to determine where in the brain they are located. However, in contrast to the reproducible, well-localized brain responses that have been found to sensory stimuli consistently localized activations associated with modules assumed to underlie mental processes supporting complex behavioral states have not been found. 2
APPLICATION TO IMAGING STUDIES OF THE STATE OF CONSCIOUSNESS
The model described above was used to study the behavioral state of consciousness through a meta-analysis of PET studies measuring CMRglc or CMRO2 in volunteers at different levels of anesthesia. 14 The state of consciousness was defined behaviorally as the capability to respond to simple cognitive questions, which is the standard that anesthesiologists use in assessing if a patient is conscious or unconscious. Similar definitions have been made by others including the intransitive consciousness of Bennett and Hacker 137 , who pointed out that this assignment of a behavioral state was objective, something that could be accepted by all observers.
The energetic basis for the loss of consciousness has been linked to overt behavioral indices. 14 In the studies reviewed, PET images of CMRglc (or CMRO2) were obtained in subjects before and during anesthesia 138 and then compared with their behavioral responses. Subjects at different levels of anesthesia answered a series of questions and were considered to be in the state of consciousness by their answers. 139
Figure 8A shows a plot of the probability of a subject not being able to respond (i.e., behaviorally no longer in the state of consciousness) versus anesthesia dosage. At a dose of about 0.6% sevoflurane approximately half of the subjects were no longer in the state of consciousness. 139 As there are situations where people may be conscious but cannot respond (e.g., locked in syndrome) but may have a variable degree of consciousness, we limit the definition of unresponsiveness to people who have full expressive faculties.
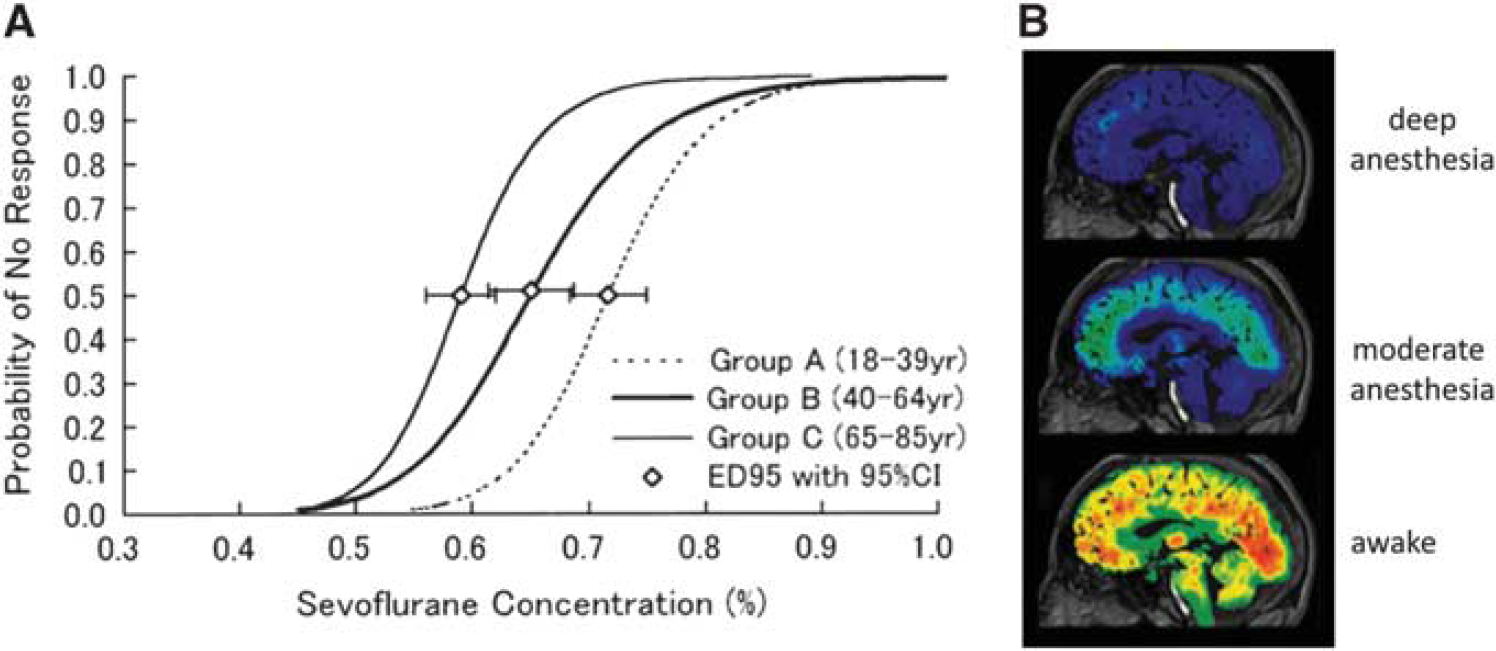
Loss of consciousness with anesthesia, as assessed by (
Figure 8B shows PET data from a single subject who was exposed to different levels of anesthesia from the awake state. 138 Different anesthetic depths, increasing from bottom to top, reduced CMRglc uniformly throughout the brain. These findings suggest that the conscious state depends on total cerebral energy. As seen in Figure 8 in the non-conscious, deeply anesthetized state (similar to that achieved during surgery) the average reduction in cortical CMRglc or CMRO2 (and by inference the neuronal activity) was on the order of 40% to 50% throughout the cortex, whereas differences between regions were less than 10% of the average decrease.
The importance of the total brain energy for maintaining the state of consciousness was noted previously, but not assigned as relevant for consciousness due to the focus on small differences in the percent reduction of glucose consumption between regions, consistent with the standard model. 140 In similar PET studies over a range of anesthetics and vegetative states, the standard model was applied to locate the neuronal basis of the mental processes of consciousness, on the assumption that such processes can be localized.90,97,141–144 Images taken in the awake state before anesthesia and in the unconscious state, during anesthesia or in vegetative state, were subtracted. The regions with the largest percentage drop in CMRglc or CMRO2 were then assigned to the mental process of consciousness. However the differences were very small compared with the overall drop in CMRglc or CMRO2 throughout the brain. With the assignment from neuroenergetic studies of these global decreases to neuronal activity the ability to assign a region as selectively shut down becomes ambiguous.
High cortical energy consumption supports the state of consciousness
We have proposed 14 that high cortical energy consumption measured in all the cortical regions is a brain property necessary to support the person in the state of consciousness. This high energy is not sufficient to support the state of consciousness, as there can be high brain activity and energy consumption in other states such as seizure, without the person being in the behavioral state of consciousness. A similar support has been found in PET studies of sleep where the reductions in energy metabolism in stage 3 and 4 sleep are also relatively uniform91,92 and not dissimilar to what has been reported for anesthesia. The results do not rule out the possibility of a smaller region in the brain having a more critical role but if so its neuronal activity is not clearly separable from the activity of the rest of the brain.
The high cortical energy consumption fulfills the role envisioned for neuronal correlates enabling consciousness described by Koch. 145 Although suggesting that high global brain activity satisfies the role for an enabling condition, we disagree with Koch's view that, ‘brain activity is both necessary and sufficient for biological sentience’ 145 . Rather, we propose that an understanding of the conscious state should build on neurophysiological properties that are necessary for, but not sufficient for, the person's ability to have complex behavioral functions.
We note briefly that the high level of baseline neuronal activity has been incorporated into an alternate model developed by Raichle 146 . In this model the state of consciousness is maintained by a uniquely high neuronal activity specifically in the default mode network, which as discussed above is not observed (Figure 5), because no specific brain network possesses higher energy demand than the global energy demand. 19
The different magnitudes of baseline energies in the awake and anesthetized states, provide a physical basis for Llinas's views that ‘sensory cues earn representation via their impact on the preexisting functional disposition of the brain’. 147 Llinas, from many electrophysiological studies, proposed that the 40 Hz electrical signals and their synchronicity 148 recorded on the scalp, measure consciousness, and bind together separate brain activities. Crick and Koch 149 extended this idea to propose that the 40 Hz oscillations bind together the different inputs to thereby create awareness. Although its coherence may or may not have a role in binding together brain activities, its increased magnitude in the conscious state is most likely a consequence of the high overall baseline neuronal activity as shown in Figure 5. Further support for the linkage between coherent electrical activity as measured through gamma band EEG oscillations, higher cognitive functions, and brain energy metabolism have been more recently provided by several studies.150–153
MODULARITY AND ITS LACK IN BRAIN RESPONSES
Functional magnetic resonance imaging studies of humans have been able to identify separate visual and other sensory processing regions that are consistent with earlier atlases based on electrical and metabolic studies of animals compiled by Zeki. 154 However despite strenuous efforts to obtain modularity, unique brain regional responses that reproducibly map to higher order mental processes have not been clearly found. 155 Although individual studies usually assign mental processes to highly localized brain regions, there is considerable variation in the area mapped in different studies when a study is repeated on the same subject or a group of subjects. The lack of experimental reproducibility within and between subjects has been recognized 2 and attributed to various causes including fMRI signal to noise and the need for better concepts. 156 In addition, it has been shown that brain responses are context dependent. For example a series of electrophysiology and fMRI studies have found that the degree to which a subject pays attention to a stimulus can influence the brain response even in sensory regions such as the visual cortex. 157 There have been attempts to maintain the link between neuronal activity and specific mental modules by proposing internal connections between modules allowing them to interact non linearly.156,158
The paradox of well localized responses to sensory stimulation and less well-localized responses to cognitive studies may partially be due to the lower changes in signal induced by the latter. This point was shown dramatically recently in a study by Bandettini and colleagues 159 who found that the number of brain areas involved in a ‘simple visual stimulation plus attention control task’ increased with signal to noise to the point where at very high signal to noise a significant change in the fMRI signal was observed throughout the entire cerebral cortex. A similar observation was made by Xu et al 160 who found that increasing the imaging signalto- noise ratio increased the area of activation in olfactory tasks.
In the standard model, the fusiform gyrus was proposed, from fMRI, single-unit recording of neuronal responses, and from lesions in humans and in nonhuman primates to be an innate, specialized region responsible for facial recognition and was given the name, the Fusiform Face Area. The innate nature of the Fusiform Face Area was challenged in the late 1990's by experiments that showed how the Fusiform Face Area responded to training and prior experience of the subject. 161 In one of these studies experimenters showed subjects images of Greebles, a class of novel head-like objects and compared the fMRI signal obtained from right side up Greebles to upside down Greebles. In untrained human subjects, there was only a small difference signal between the response to right side up and upside down Greebles. However, after the same human subjects were trained to identify right side up Greebles, the difference signal between right side up and inverted Greebles was large and similar to the normal response to faces in fMRI experiments.
The ability to recruit the Fusiform Face Area led to a reformulation of its modular definition and it is now largely assigned to ‘expertise’ in the mental process of visual object recognition as opposed to being face recognition specific. A similar reformulation proposing this change, e.g., that the brain can recruit different modules after practice, was made by Petersen et al 162 who showed that when a subject performs novel verbal or motor tasks and then practices the tasks the induced brain activity shifts from frontal to parietal regions. Although the results of the Greebles and related experiments on recognition expertise have been incorporated into models of modular function in this way, we propose that the results are more consistent with a model in which the brain response depends on the understanding that the person obtains by experience. 3 Expertise in Greeble recognition has not brought expertise in recognizing birds or cars. That depends on experience as opposed to activation of an intrinsic module. Although a person's knowledge of sensory stimuli is usually very reproducible from common experiences extending back to his earliest years, 136 his knowledge of higher order concepts varies with his individual experience and therefore with its context. With an inconsistent understanding of higher order concepts, the brain response to them will not be reproducible, as has been found experimentally (e.g., see McGonigle et al 163 and Otzenberger et al 164 ).
AN ALTERNATE MODEL TO EXPLAIN MODULARITY AND NONMODULARITY IN FMRI STUDIES
In this section, we describe an alternate model to interpret functional imaging studies, which does not assume that hypothesized mental processes provide necessary and sufficient causes of behavior. The model was developed from consideration of experimental studies described above and from philosophical considerations that are touched on below but described in more detail in a recent book. 3
The first point of this model is that ‘only of a living human being and what resembles (behaves like) a living human being can one say: it has sensations; it sees; is blind; hears; is deaf; is conscious or unconscious’. 165 This fundamentally differs from the claim in cognitive neuroscience that the brain causes the person's behavior. Rather in this model it is the person's behavior that is an observable starting point and experiments can find brain activities necessary for that behavior. This point, originally made by Wittgenstein was developed by Bennett and Hacker 137 who claimed that it was incorrect to assign a property of the organism to a part of it, such as the brain, a logical failing which is called the mereological fallacy. In following this metaphysical point our starting point for investigating brain function is the observation of the person's behavior.
The second premise of the model is that although empirical brain activity can be found that is necessary for the person to have a certain behavior that activity is not sufficient to cause the behavior. Necessary mechanisms can develop an understanding of the behavior, building up what has been called a ‘Mosaic’ as more and more of the necessary properties are found. 166 From this viewpoint a healthy medial temporal lobe is necessary for a person to have Long Term Memory but this brain activity is not sufficient to explain the behavior. Another example, illustrated in imaging studies of the state of consciousness, is that high global baseline energy is needed for a person to be in the state of consciousness but is not sufficient for that to happen.
The third point is that observable behavior is supported by brain activity and that substituting a mental process/module as an intermediate between brain activity and behavior lacks the reliability offered by limiting our experimental parameters to observables of brain activity and behavior. This avoidance of mental processes may be difficult to accept because these generalizations are of considerable value in daily life and because understanding what is meant by them has been a principle goal of philosophical enquiry. Disregarding philosophical and psychological efforts to define generalizations behind behavior (e.g. substituting memory for the act of remembering) has been advocated most directly by Ludwig Wittgenstein, who criticized ‘ the contemptuous attitude towards the particular case’. He continued by regretting that people assume 167 ‘the idea that properties are ingredients of things which have the properties; e.g. that beauty is an ingredient of all beautiful things as alcohol is of beer and wine’. In agreement with this criticism our model identifies brain activities as properties of an observable behavior and disregards generalizations of mental processes claimed to define that state. This avoidance of mental processes/modules in the interpretation of fMRI data is the fundamental distinction between our model, equation (9), and the prevailing model of Cognitive Neuroscience, equation (8). The advantage of avoiding mental modules and sticking to observables is that we can continue to study brain properties of the state of consciousness by reliable physical measures.
In a fourth point, even in cases of apparent modularity such as in primary sensory systems, the reproducible brain responses are not innate but rather depend highly on a person's understanding gained from experience. Plasticity allows the brain responses to change with experience so as to facilitate the brain's support of the person's behavior. In this way brain activity helps the person to adapt to his environment and thereby to survive.
Fifth, the brain response is reproducible to the extent that the person reproducibly recognizes a stimulus. On the neuronal level, this reproducibility would be a similar distribution of neuronal firing rates every time the subject was exposed to the stimulus. This degree of reproducibility has been found in sensory tasks in animal models and in the consistent response observed in animal models after they have been highly trained to perform cognitive tasks such as working memory, a degree of training that fortunately is generally not allowed for human subjects.
A key philosophical basis of the model is that the meaning of a word describing a behavior or state is neither innate nor generally valid but rather is determined by the person's experience and understanding. 168 In Cognitive Neuroscience the belief that objective, innate mental processes correspond to definitions of concepts that encompass words like memory, intention, honor or love led to the expectation that functional imaging can identify the objective neuronal basis of these abstractions. However, in our alternate interpretation, there is no more reproducibility in the neuronal activity supporting these concepts than can be found in the person's understanding of these concepts in the first place. To the extent that the person has a reproducible understanding of what these concepts mean (or equivalently recognizes objects such as Greebles) the brain's neuronal response will be reproducible. As people have normally been trained during their early years to recognize sensory stimuli their brains will show similar patterns of neuronal activity when the same stimuli are presented, and when summed will give modular fMRI signals. However, as generalized behaviors like remembering, calculating or paying attention depend on their context and on the person's history, the neuronal activity needed to support them will vary and will not yield reproducible, modular fMRI signals. Physically reliable experiments can continue to improve our understanding of the brain properties necessary for behavior providing the results are not interpreted by such psychological generalizations.
Accepting the model of brain function proposed here, in place of the positivistic approach of cognitive neuroscience, returns the responsibility for defining higher order behaviors to the properties of the individual. A person's understanding is both individualistic and historical and the brain's neuronal activity, supporting that understanding, will be analogously unique. To the extent that a psychology can be integrated into the study of brain function it must replace the causal, general laws proposed by cognitive neuroscience with an understanding that reflects the individualistic and historical nature of the living organism. It has been noted by Shulman et al169,170 that Freudian psychology by attempting to provide a structure that could reconcile the individual's history with observations of behavior could contribute to the model of brain function that we have proposed. The physical information available from brain imaging requires a model of brain function that brings us to the interface where physical understanding must enter into dialogue with the individuality and history of living organisms. By abandoning uncertain psychological attributes we are able to build an understanding of the properties of behavior based on reliable neurophysiological measurements.
Footnotes
The authors declare no conflict of interest.
ACKNOWLEDGMENTS
We thank the National Institute of Health for their support through the following grants: R01 AG034953, R01 MH067528, P30 NS052519.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.