
Abstract
Network activation triggers a significant energy metabolism increase in both neurons and astrocytes. Questions of the primary neuronal energy substrate (e.g., glucose vs. lactate) as well as the relative contributions of glycolysis and oxidative phosphorylation and their cellular origin (neurons vs. astrocytes) are still a matter of debates. Using simultaneous measurements of electrophysiological and metabolic parameters during synaptic stimulation in hippocampal slices from mature mice, we show that neurons and astrocytes use both glycolysis and oxidative phosphorylation to meet their energy demands. Supplementation or replacement of glucose in artificial cerebrospinal fluid (ACSF) with pyruvate or lactate strongly modifies parameters related to network activity-triggered energy metabolism. These effects are not induced by changes in ATP content, pHi, [Ca2+]i or accumulation of reactive oxygen species. Our results suggest that during network activation, a significant fraction of NAD(P)H response (its overshoot phase) corresponds to glycolysis and the changes in cytosolic NAD(P)H and mitochondrial FAD are coupled. Our data do not support the hypothesis of a preferential utilization of astrocyte-released lactate by neurons during network activation in slices—instead, we show that during such activity glucose is an effective energy substrate for both neurons and astrocytes.
INTRODUCTION
High cellular energy demands during network activation are met by upregulation of cytosolic glycolysis and mitochondrial oxidative phosphorylation. Mitochondrial metabolism provides most of the ATP but glycolysis is also enhanced and may contribute to the energy production.1, 2, 3, 4 For elucidation of the cellular basis of neuroenergetics, measurements of metabolic signals including the oxygen utilization and NAD(P)H/FAD autofluorescence provide valuable information for connecting energy metabolism with neuronal activity. NADH (reduced form) is fluorescent when excited with UV light whereas NAD+ is not, leading to a decrease in observed fluorescence as a result of NADH oxidation. In contrast, FAD (oxidized form) is fluorescent, so the oxidation of FADH2 to FAD causes an increase in fluorescence. The fluorescence of NADH cannot be separated from that of NADPH and their emission is measured in concert (NAD(P)H). NAD(P)H fluorescence represents a ‘mixed’ signal since this cofactor can be produced by both glycolysis and mitochondria, whereas FAD fluorescence is entirely mitochondrial.5, 6 Measurements of these parameters in combination with electrophysiological recordings have been used in many studies to monitor the energy status during neuronal activity in brain tissues.
Typically, NAD(P)H transients induced by synaptic stimulation have a characteristic biphasic waveform: the initial short dip is followed by a long-lasting overshoot. While there exists a common consensus that the initial decrease in fluorescence is due to mitochondrial function, the origin of the overshoot is as yet unclear. A radical solution to this problem has been proposed by Kaschiske
Another important issue concerns the preferred energy substrate for supporting cellular activity. Glucose is the main energy substrate supplied to the adult brain under normal conditions, although the exact supply pathways and cell types that consume glucose during network activation are not fully elucidated. Brain cells are capable of utilizing varying energy substrates in addition to glucose. 13 For instance, the ‘lactate shuttle’ hypothesis postulates that lactate, as the end product of astrocytic glycolysis triggered by glutamate uptake, is the main fuel for neurons during their activity. 14 Glucose is thus the essential substrate for energy generation throughout network activity, but its metabolism is segregated into two cell types—glycolysis in astrocytes and oxidation in neurons. The relevance of such a scenario, however, has been questioned (for review, refs. 15, 16, 17) and views on the major fuel source for activated neurons and the corresponding neuron–astrocyte interactions are controversial.
Therefore, the roles of glycolysis and oxidative phosphorylation in energy metabolism of neurons and astrocytes during network activation are still debatable. This is an issue of principal importance for furthering our understanding of cellular specificities of energy metabolism and their coupling with neuronal–astrocytic interactions. We addressed these questions in the present study, utilizing simultaneous electrophysiological and metabolic recordings in hippocampal slices from mature mice.
MATERIALS AND METHODS
Tissue Slice Preparation
Brain slices were prepared from P19-P44 Swiss mice of both sexes. However, from 64 mice used in the experiments, only 5 mice were younger 30 days, and therefore we mention the animals as ‘mature’ in the text. All animal protocols conformed the INSERM guidelines on the use of laboratory animals and are approved by the Ethics Committee for Animal Experimentation of Marseille (#30-03102012). A mouse anesthetized with isoflurane was decapitated, the brain was rapidly removed from the skull and placed in the ice-cold artificial cerebrospinal fluid (ACSF). The ACSF solution consisted of (in mmol/L): NaCl 124, KCl 3.50, NaH2PO4 1.25, NaHCO3 25, CaCl2 2.00, MgCl2 1.30, and dextrose 10, pH 7.4. ACSF was aerated with 95% O2/5% CO2 gas mixture. Sagittal slices (350
Synaptic Stimulation and Field Potential Recordings
Shaffer collateral/commissural pathway was stimulated using the DS2A isolated stimulator (Digitimer Ltd, Hertfordshire, UK) with a bipolar metal electrode situated in the stratum radiatum of CA1 hippocampal region. To reduce the stimulated area in some experiments, we used a bipolar glass electrode (30
NAD(P)H and FAD Fluorescence Imaging
NADPH and NADH have similar optical properties; therefore, it is expected that NADPH may contribute to the total autofluorescence signal. Changes in NAD(P)H fluorescence in hippocampal slices were monitored using a 290–370 nm excitation filter and a 420 nm long-pass filter for the emission (Omega Optical, Brattleboro, VT, USA). In the case of FAD imaging, 500 to 570 nm emission was excited with 400 to 490 nm light. The light source was the Intensiligh C-HGFI illuminator (Nikon Instruments Europe B.V., Amsterdam, Netherland) equipped with a mercury arc lamp. Slices were epi-illuminated and imaged through a Nikon upright microscope (FN1, Eclipse; Nikon Instruments Europe B.V.) with × 4/0.10 Nikon Plan objective. Images were acquired using a linear, cooled 12-bit CCD camera (Sensicam; PCO AG, Kelheim, Germany) with a 640 × 480 digital spatial resolution. Because of a low level of fluorescence emission, NAD(P)H or FAD images were acquired every 600 to 800 ms as 8 × 8 binned images (effective spatial resolution of 80 × 60 pixels). The exposure time was adjusted to obtain a baseline fluorescence intensity between 2,000 and 3,000 optical intensity levels. Fluorescence intensity changes in stratum radiatum near sites of LFP and O2 recordings were measured in three to five regions of interest (ROIs) using the ImageJ software (NIH, USA). Data were expressed as the percentage changes in fluorescence over a baseline ((Δ
Oxygen Measurements
A Clark-style oxygen microelectrode (tip diameter 10
Intracellular pH and Ca2+ Imaging
pH and Ca2+ were measured in two separate experiment series using ratiometric imaging approach. For Ca2+ measurements, brain slices were loaded with Fura-2 AM (Molecular Probes, Life Technologies, St Aubin, France), and with SNARF-1 (Invitrogen, Life Technologies) for pH evaluation (see details in Supplementary Methods).
ATP Measurements
Brain slices from one animal were divided into two groups. One group was incubated for 40 minutes at 32°C in 5 mmol/L glucose-ACSF whereas another one in 10 mmol/L pyruvate-ACSF. ATP quantity was determined in each group using the Stay Brite ATP Bioluminiscence assay kit (BioVision Inc., Milpitas, CA, USA). ATP content in slices was normalized to total protein content determined using Bio-Rad protein assay (see Supplementary Methods).
Pharmacology
Drugs used were purchased from Sigma (Sigma-Aldrich Chimie S.a.r.l., Lyon, France) (
Statistical Analysis
Group measures were expressed as means±s.e.m. Statistical significance was assessed using Student's paired
RESULTS
In several studies on slices, it has been shown that the waveshape of NAD(P)H response to synaptic stimulation depends strongly on the availability of oxygen: an insufficient pO2 results in a decrease in oxidation phase and an increase in overshoot.10, 18, 19, 20 In brain slices, pO2 decreases with depth and cells located at different depths may contribute differentially to the acquired NAD(P)H signal, hindering its adequate interpretation. Therefore, we first had to confirm that the NAD(P)H recordings were not ‘contaminated’ by such artifactual conditions and that the NAD(P)H transient waveshapes are similar throughout the slice depth. We used a high superfusion rate (15 mL/min) together with solution access to the both sides of the slice to ensure adequate slice oxygenation18, 21, 22 and tested NAD(P)H signaling using a local Shaffer collaterals stimulation by a double-barrel glass electrode while simultaneously measuring pO2 at the same depth (Figure 1A). Note that the variability in electrodes' location during intrusion into the slice underlies the difference in response values in Figure 1A. For instance, a smaller NAD(P)H response at 300
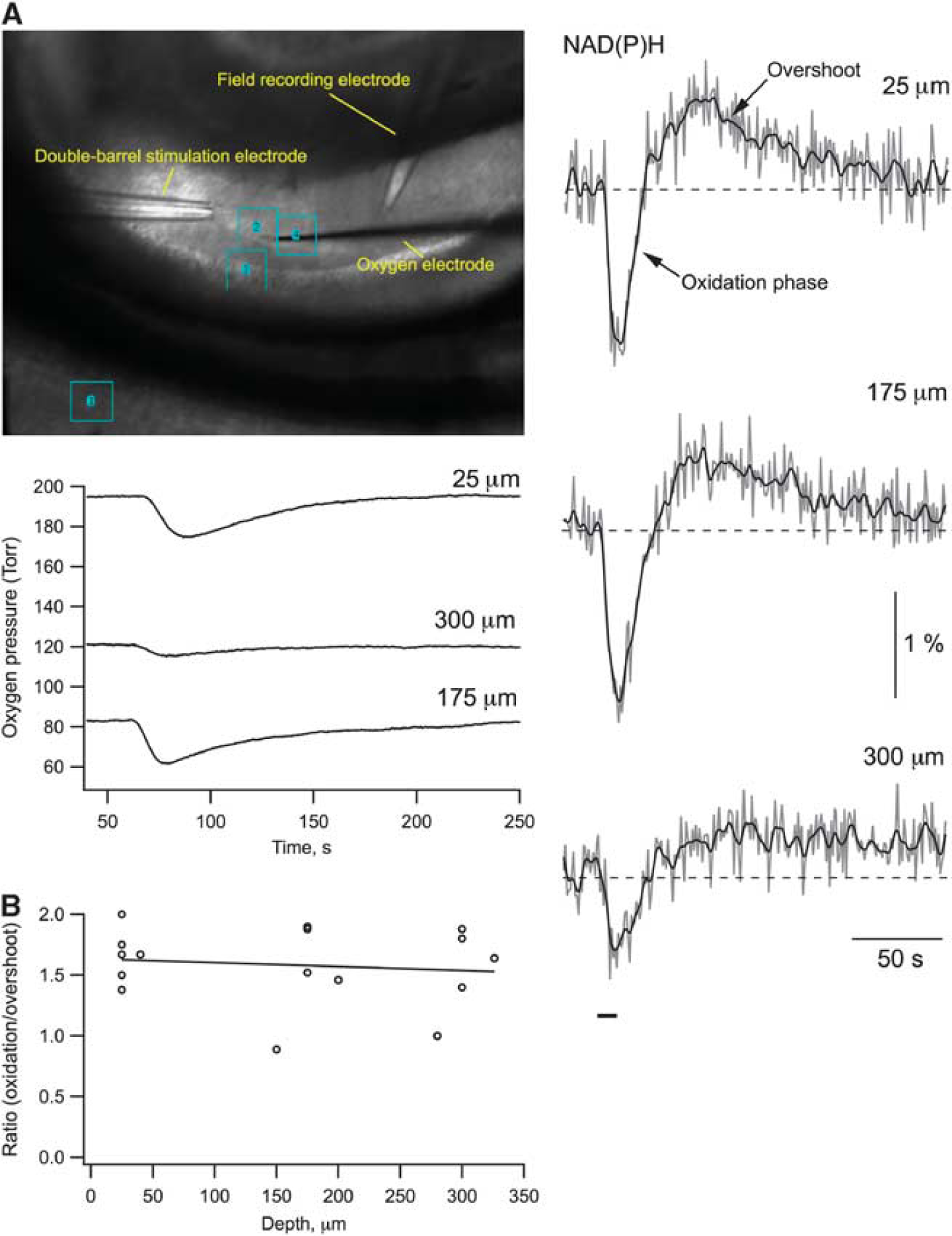
NAD(P)H transient waveshape is similar throughout the depth of the slice. (
Contribution of Neurons and Astrocytes to the Energy Production During Evoked Activity
Stimulation-triggered presynaptic neurotransmitter release from Shaffer collateral terminals activates energy metabolism in both CA1 neurons and astrocytes. In neurons, energy is mainly spent on recovering the transmembrane potential and ion gradients perturbed by the various ion channel-mediated currents, whereas in astrocytes energy is mostly spent on the glutamate and potassium uptake.2, 3, 9 To pharmacologically separate the postsynaptic neuronal fraction of the observed response, we used a mixture (cocktail) of postsynaptic receptor blockers consisting of NBQX (antagonist of AMPA receptors, 10
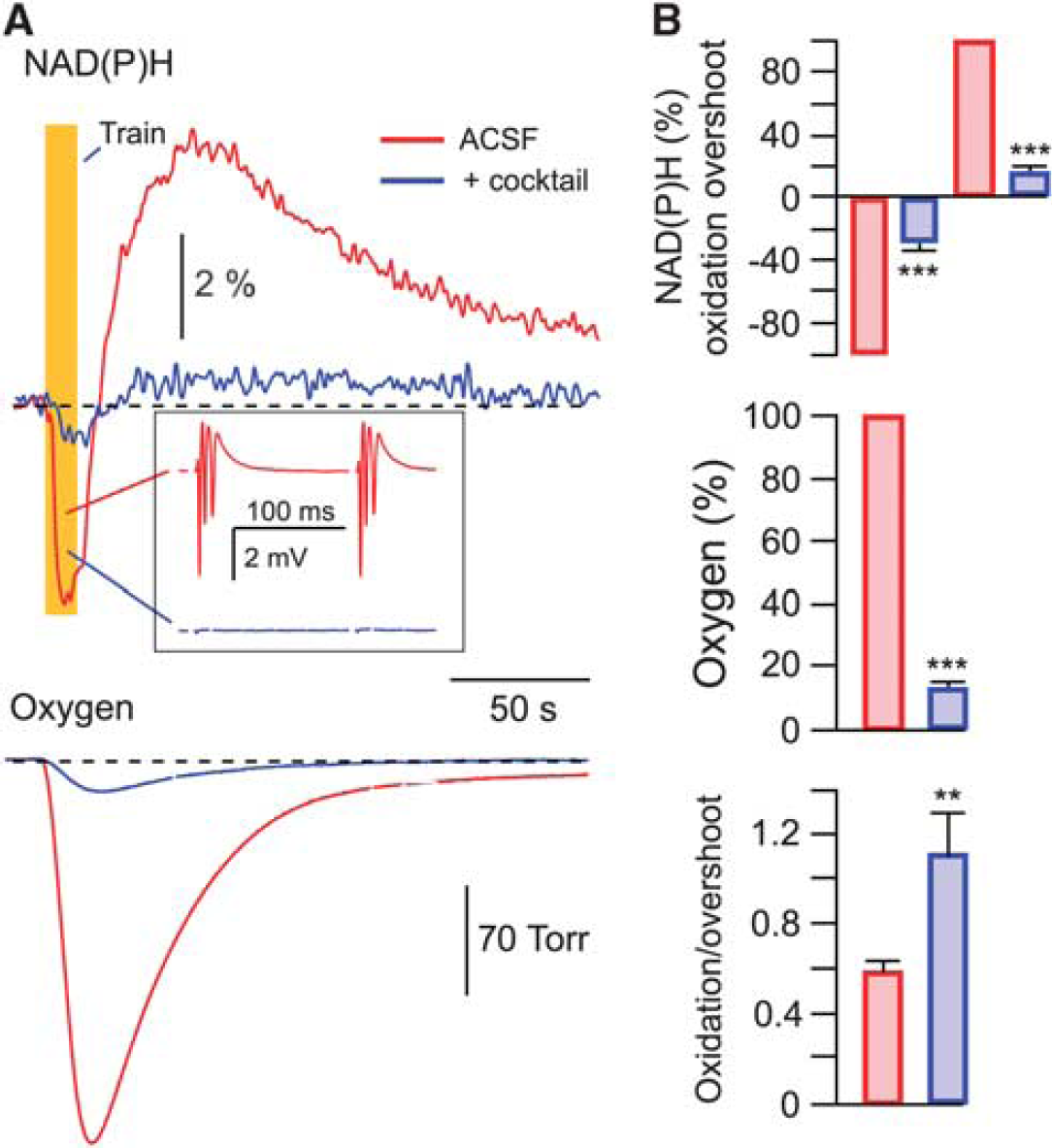
Neuronal and astrocytic metabolic signaling induced by synaptic stimulation. (
To reveal in detail the postsynaptic neuronal and astrocytic metabolic activities ‘uncontaminated’ by presynaptic components, we utilized a different type of stimulation—a 10-Hz train of short (10 ms) puffs of glutamate (10 mmol/L) directly onto the recorded region. Glutamate application can serve as metabolic stimulation due to induced neuronal electrical activity
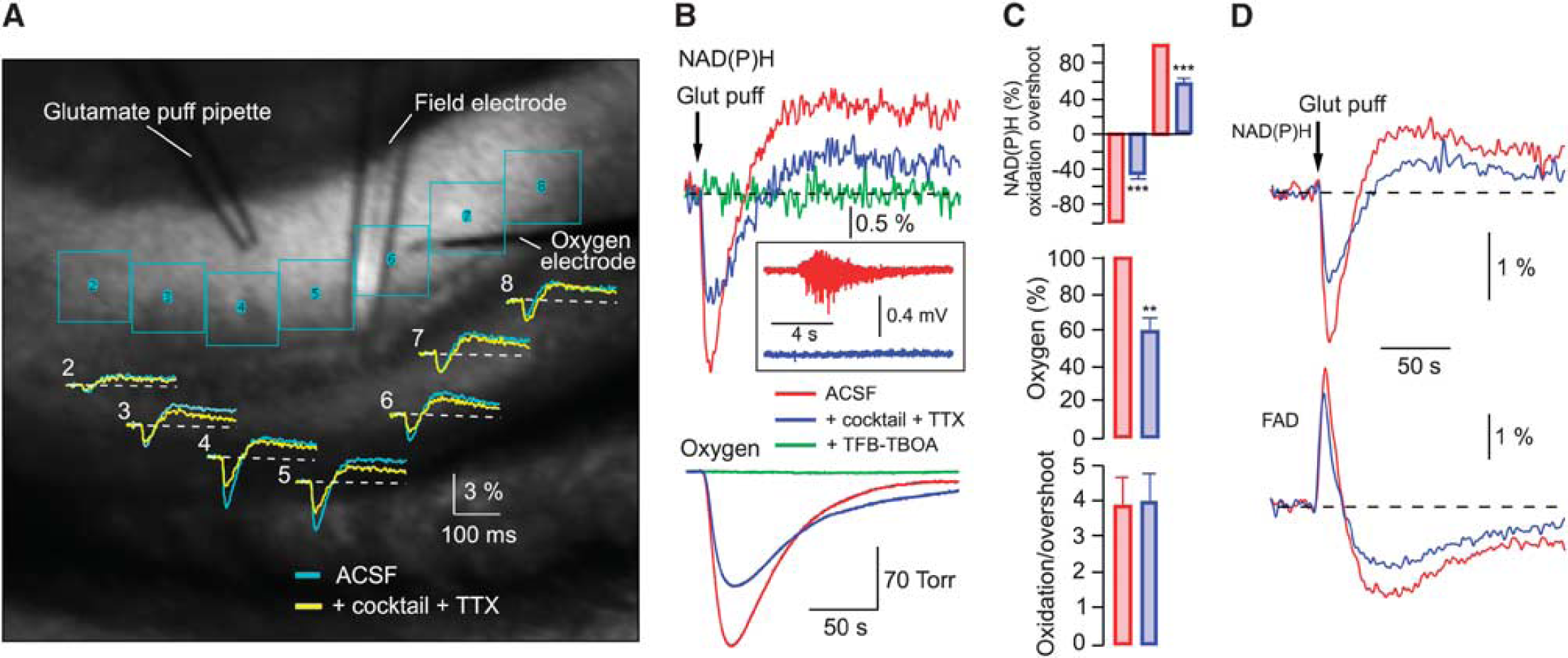
Neuronal and astrocytic metabolic signaling induced by the glutamate puff train stimulation. (
To further confirm this conclusion, we recorded the FAD autofluorescence originating from mitochondria.5, 6 In six experiments, we recorded both NAD(P)H and FAD transients induced by the 5-second, 10-Hz glutamate puff trains. Figure 3D shows these signals' combined average. Prominent FAD fluorescence remaining after the cocktail+TTX application confirms the intense mitochondrial activity in astrocytes. Note that with both types of stimulation (electrical-synaptic and glutamate puffs) a significant fraction of the overshoot phase disappeared after the cocktail administration, indicating that it corresponds to neuronal activity. Thus, the metabolic processes associated with the oxidation and overshoot phases of NAD(P)H transients are active in both neurons and astrocytes.
The Link between NAD(P)H Overshoot and Glycolysis
The cell type-specific role of cytosolic glycolysis and mitochondrial oxidative phosphorylation involved in energy generation is a question of primary importance for the conceptual understanding of energy metabolism and neuronal–astrocytic interactions. Since NAD(P)H is produced by both glycolysis and citric acid cycle, the analysis of NAD(P)H autofluorescence may provide important information on energy metabolism components. Indeed, glucose metabolism by glycolysis provides the end product pyruvate, which further serves as a primary substrate for mitochondrial metabolism. Therefore, to distinguish the glycolytic and mitochondrial contributions to the NAD(P)H fluorescence, we compared metabolic signaling recorded in standard ACSF with that recorded after complete replacement of ACSF glucose with pyruvate. Such a substitution was expected to arrest glycolysis while maintaining the mitochondrial function. Importantly, the substitution of energy substrates did not result in any change in ATP content in slices (nmoles/mg of total protein: 9.38±1.05 in 5 mmol/L glucose-ACSF and 9.47±1.28 in 10 mmol/L pyruvate-ACSF;
In each experiment, similar stimulation intensities (single-pulse Shaffer collateral stimulation) in both substrate-containing solutions activated a similar number of nerve fibers, as manifested by the unaffected size of presynaptic volley (e.g., see Supplementary Figure 2A). However, in all such single-pulse experiments (
The glucose-for-pyruvate exchange could modify the basal cellular redox state and shift NAD(P)H/FAD basal fluorescences (
NADH fluorescence depends on the glucose and pyruvate content in artificial cerebrospinal fluid (ACSF). (
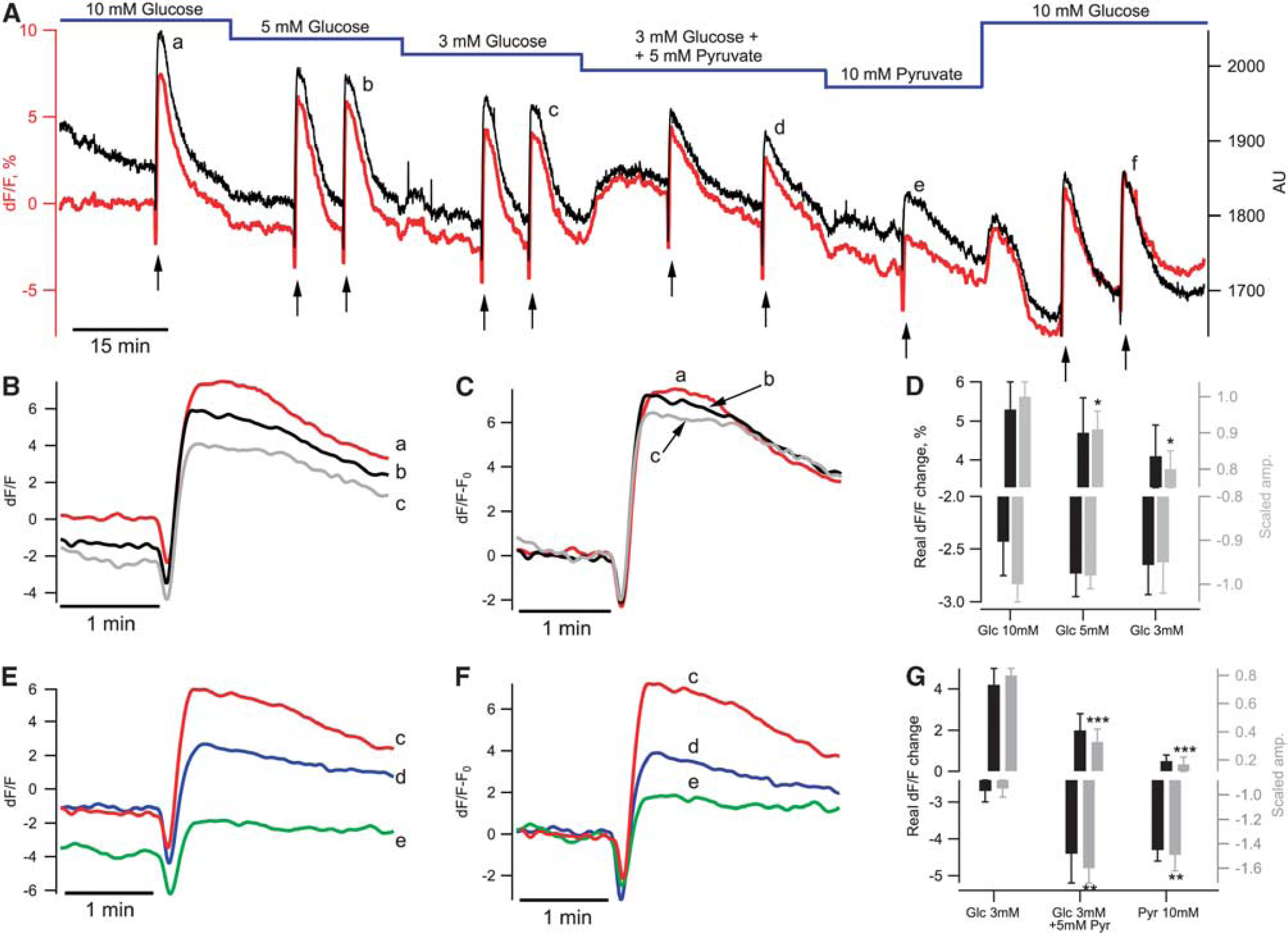
After the initial control 15 minutes, we started Shaffer collateral pulse train stimulation (10 Hz, 10 seconds) and also exchanged ACSF energy substrates (Figure 4A, red). Train stimulations were applied 1 to 2 times in each condition. A decrease in glucose concentration from 10 to 5 mmol/L did not induce significant decreases in basal NAD(P)H fluorescence (−1.9±0.8%,
We also compared NAD(P)H transients evoked by Shaffer collateral train stimulation (shown by arrows in Figure 4A) at different stages of the experiment. A decrease in ACSF glucose concentration had no effect on the oxidative dip (−2.4±0.3%, −2.7±0.2%, and −2.7±0.3% in 10, 5, and 3 mmol/L glucose, respectively; Figure 4D, black bars (corresponded to −1.00±0.04%, −0.98±0.03%, −0.95±0.07% of response in 10 mmol/L glucose,
These results show that a low glucose concentration (3 mmol/L) in ACSF or supplementing/replacement glucose with pyruvate decreased both the overshoot of NAD(P)H transients and the basal NAD(P)H fluorescence. However, the maximal baseline fluorescence fluctuations did not exceed 200 arbitrary units in long-lasting (>2 hours) recordings, representing ∼10% of the NAD(P)H fluorescence at the beginning of experiments. Thus, the maximal error that might be introduced by Δ
Figure 5A shows the original traces from an experiment in which recordings have been performed on the same slice and under similar conditions initially in glucose-ACSF followed by those in pyruvate-ACSF. Train stimulation of Shaffer collaterals resulted in radically different NAD(P)H responses in the presence of either glucose or pyruvate: in pyruvate, the oxidation phase became considerably larger (by 62±5%; (from −3.31±0.11% to −5.33±0.2% amplitude)) while the overshoot strongly decreased (by 72±3%; (from 4.13±0.41% to 1.15±0.15% amplitude)). Similar changes were observed in all 24 experiments (Figure 5B;
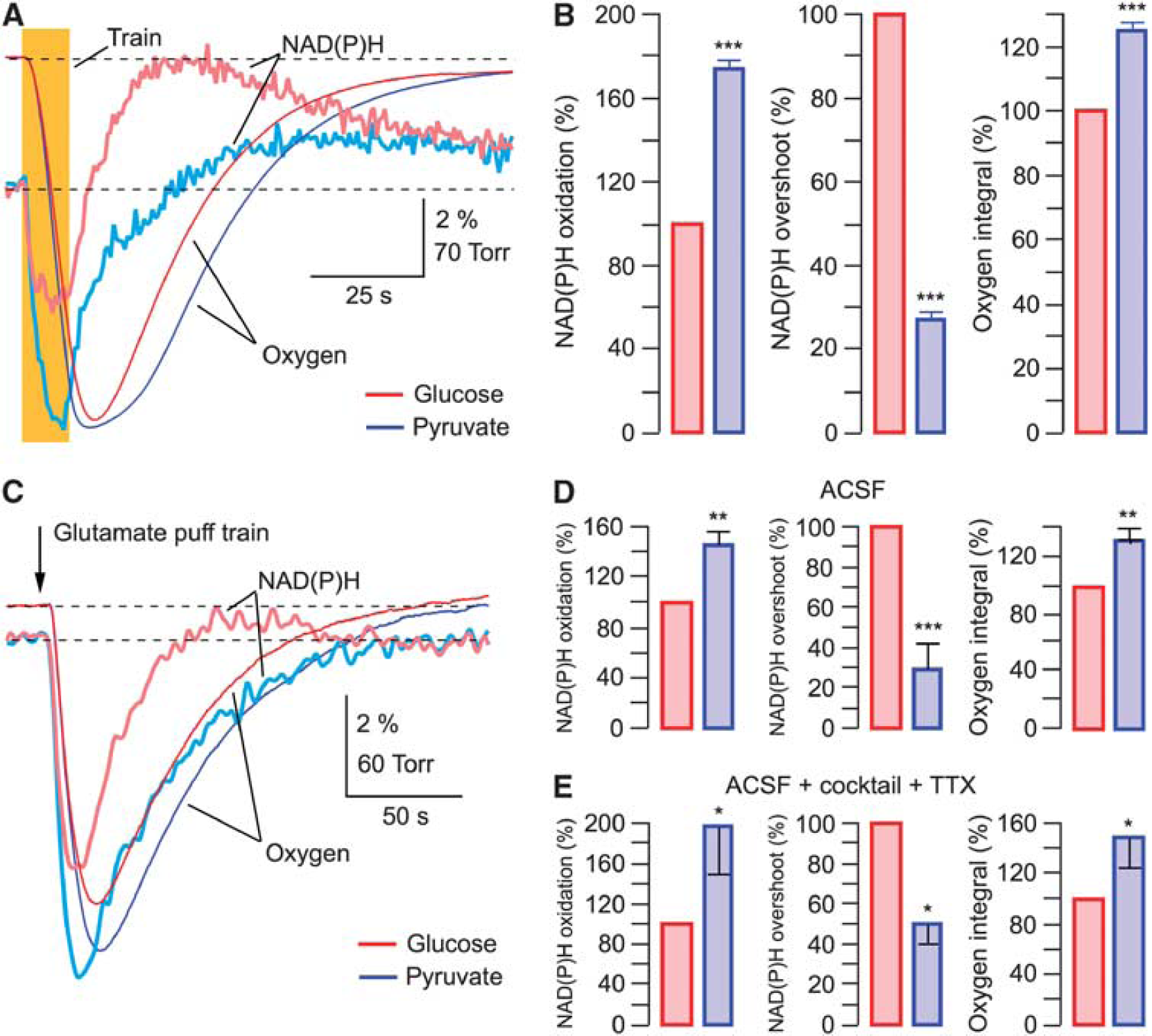
Overshoot in NAD(P)H transients mainly reflects the glycolysis-generated NAD(P)H. (
When NAD(P)H profiles are aligned with pO2 traces (Supplementary Figure 4) it is evident that in glucose-ACSF, NAD(P)H fluorescence decreases to its minimum and starts to recover while oxygen consumption (d(pO2)/d
It is important to note that the oxygen consumption after synaptic activation was always significantly larger in pyruvate-ACSF (by 25±4%) pointing to a more active oxidative phosphorylation compared with glucose-ACSF. This suggests that in pyruvate-ACSF, mitochondrial metabolism is enhanced to compensate for a significant part of energy needs which are normally covered by the glycolysis-produced ATP and NADH in glucose-ACSF (see Discussion).
We also performed analogous experiments using the glutamate puff train stimulation (Figures 5C to 5E). After the exchange of glucose-ACSF for pyruvate-ACSF, NAD(P)H transients were modified in a similar way as with electrical stimulation (Figure 5C): the oxidation phase strongly increased (by 45±13%,
Altogether, these results indicate that a substantial part of NAD(P)H overshoot in both neurons and astrocytes is likely related to glycolysis. An alternative possibility is that the mitochondrial function strongly differs in glucose- and pyruvate-containing ACSF. We addressed this issue in detail in the following experiments.
Potential Caveats in Exchange of Glucose to Pyruvate
Extracellular pyruvate as well as lactate are transported into cells by the monocarboxylate transporters in conjunction with H+26 and can therefore potentially induce intracellular acidification. This in turn may modulate a number of cellular processes including mitochondrial function.
27
We therefore measured the change in neuronal pHi after the replacement of 10 mmol/L glucose with 10 mmol/L pyruvate or lactate in ACSF (Figure 6D). In CA1 pyramidal neurons, during the glucose–pyruvate transition, the histogram peaks at ΔpHi=0.05 (
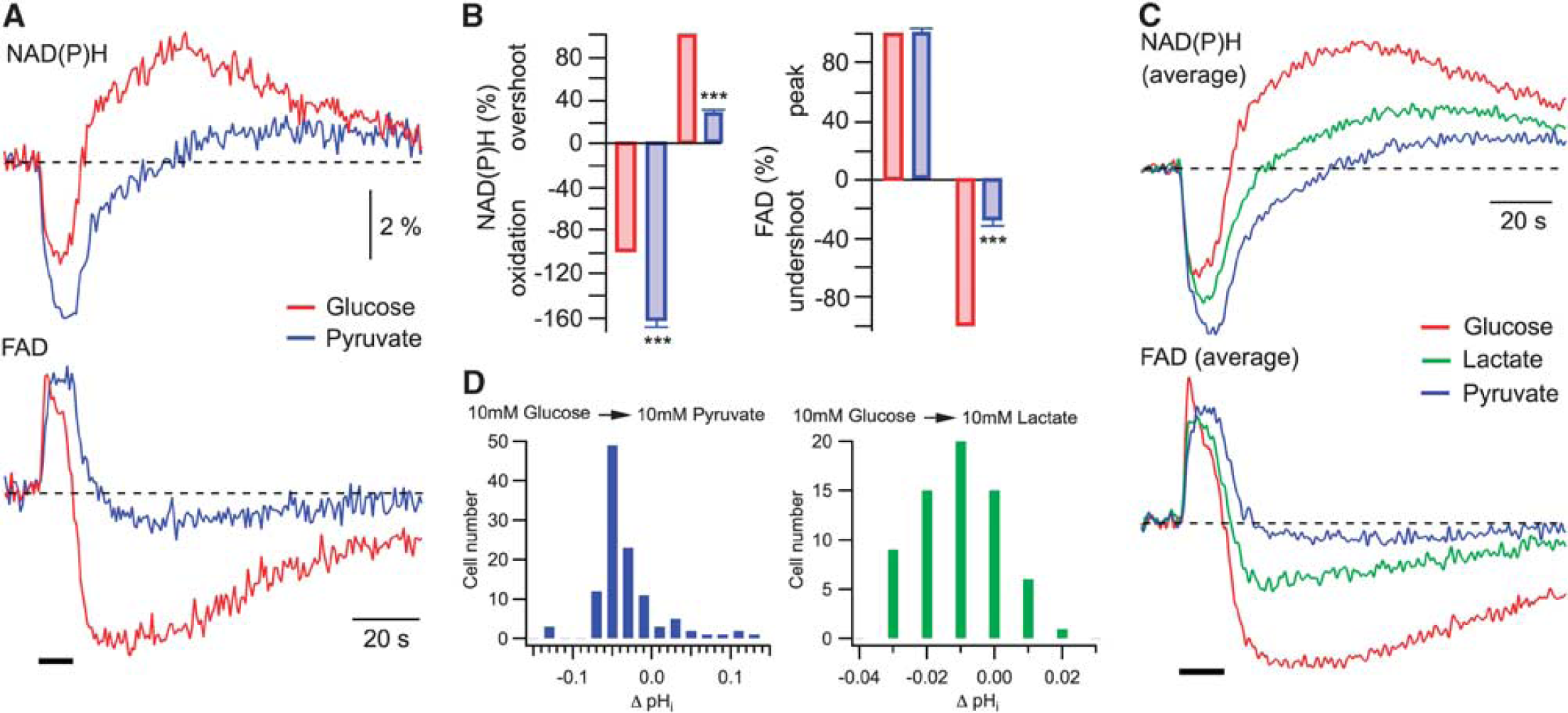
Coupling between cytosolic and mitochondrial redox states. (
One more factor that could affect metabolic parameters after the exchange of energy substrates might be an increase in intracellular Ca2+ concentration. 30 However, measurements of [Ca2+]i in CA1 pyramidal cells induced by stimulation of Shaffer collaterals (10 Hz, 10 seconds) revealed no significant difference between values recorded in glucose-ACSF and pyruvate-ACSF (Supplementary Figure 5A).
In addition, glutathione is the most abundant mammalian antioxidant. It is likely that in the absence of glucose, the cytosolic glutathione-based antioxidant system is inhibited and that may lead to the accumulation of reactive oxygen species. Therefore, we tested effects of an antioxidant (TEMPOL) on NADH/FAD transients. Tempol (4-hydroxy-2,2,6,6-tetramethylpiperidine-1-oxyl) is a member of family of nitroxide compounds that has been studied extensively in animal models of increased reactive oxygen species and is one of the most effective antioxidants. 31 In three experiments, addition of TEMPOL (2 mmol/L) to glucose- and pyruvate-ACSF did not induce significant changes in pyruvate effects on NADH/FAD autofluorescence (Supplementary Figure 5B).
The results from these experiments make unlikely the assumption that the changes in pHi, Ca2+i, and reactive oxygen species during energy substrate substitution in ACSF are the possible reasons for the observed modifications in NADH/FAD autofluorescence.
Glycolysis Affects the Mitochondrial Redox State
To further evaluate the influence of ACSF glucose-to-pyruvate transition on mitochondrial metabolism, we measured FAD autofluorescence that has been frequently used as a selective marker of mitochondrial redox state.5, 6 As was the case with NAD(P)H (Figure 4A), the FAD baseline fluorescence decreased when pyruvate was either supplemented to or completely replaced glucose in ACSF (−13.3±4.2%,
As has been observed previously,8, 19 FAD transients were inverted in respect to NAD(P)H responses and possessed a prominent undershoot, although in our experiments NAD(P)H and FAD signals clearly did not mirror each other and displayed significantly different dynamics (Figure 6A). Surprisingly, an exchange of glucose in ACSF for pyruvate resulted in a strong modification of not only NAD(P)H but also FAD transients with an almost complete disappearance of the latter's undershoot phase. Similar results were obtained in 14 experiments (Figure 6B): in pyruvate, FAD peak did not change (
Clearly, direct verification of the glycerol-3-phosphate shuttle involvement in glycolysis–mitochondrial interactions could be provided by a shuttle blockade. Unfortunately, the pharmacological tools for specific inhibition of the glycerol-3-phosphate shuttle are not yet available (McKenna
DISCUSSION
Metabolic Processes in Neurons and Astrocytes
Network activation stimulates energy metabolism in both neurons and astrocytes. The relative contribution of different cell types to the energy utilization is of interest since the energy use of astrocytes has often been ignored. To separate the neuronal and astrocytic fractions of the metabolic process, we utilized a pharmacological dissection by applying a cocktail of postsynaptic receptor antagonists, blocking neuronal activation. In the case of synaptic stimulation such a blockade was not complete as the remaining responses still contained a presynaptic neuronal fraction. An alternative stimulation by direct application of glutamate provided more evident results since here the presynaptic neuronal fraction could be eliminated by TTX. Separation of neuronal and astrocytic responses reveals their qualitative similarity: NAD(P)H transients in both cell types possess pronounced dip and overshoot (Figures 2 and 3). During synaptic stimulation, however, the relative contribution of oxidation phase (dip) was larger in astrocytes compared with neurons (see Figure 2B). Importantly, the observed NADH dip and overshoot do not directly reflect the actual amount of NADH utilized and generated and therefore cannot be used for quantitative estimation of energy production. 9 These signals may indicate, however, the relative contribution of oxidative phosphorylation and glycolysis (see below) to the overall process. In contrast, oxygen consumption is a direct correlate of oxidative phosphorylation and mitochondrial ATP generation. When induced by synaptic stimulation, astrocytic activity constitutes a notable fraction of total oxygen utilization (at least ∼15%; Figure 2B), suggesting that glial energy demands during synaptic stimulation are significant and should not be ignored when estimating the energy consumption by different network components.
Contribution of Glycolysis to the NAD(P)H Overshoot
The origin of NAD(P)H overshoot is still the matter of debates. Two opposing points of view associate the overshoot either with glycolytic activity7, 20, 21 or primarily with mitochondrial one.
8
The contradictory results of these studies may be explained in part by the experimental confines characteristic for slice preparations. Experimental observations that
To uncover the purely mitochondrial fraction of NAD(P)H signaling during synaptic stimulation, we exchanged ACSF glucose to pyruvate. This resulted in a significant decline in both NAD(P)H and FAD basal fluorescence (Figure 4A; Supplementary Figure 6A). Therefore, the mitochondrial reduction (FADH2) increased in contrast with a decreased amount of NAD(P)H.
This result cannot be explained if the mitochondrial citric acid cycle is assumed to be the major source for autofluorescence—strongly suggesting that the absence of glycolytic NAD(P)H generation underlies our observations.
Considerable overshoot reduction during stimulation in the presence of pyruvate has been reported in both early 21 and recent studies8, 18, 20 where supplementing glucose in ACSF with pyruvate was used for revealing the glycolytic NAD(P)H production. The effects of pyruvate became more prominent after a complete replacement of glucose in ACSF that resulted in: (1) a robust increase in the oxidation phase; (2) considerable slowing down of NAD(P)H recovery to baseline; (3) a strong overshoot decrease; (4) a significant enhancement (by ∼25%) of oxygen consumption. The effects (1) and (4) indicate the augmentation of mitochondrial functioning at similar energy demands and therefore suggest that in glucose-ACSF a part of energy requirements during network activity is covered by the glycolysis-generated ATP. Indeed, cytosolic glycolysis produces two ATP and two NADH. If reducing equivalents from these cytosolic NADH are transferred to mitochondrial NAD (or FAD) by the malate–aspartate (and, presumably, glycerol-phosphate shuttle) this produces two mitochondrial NADH (or FADH2) resulting in five (or three) ATP after oxidation in the respiratory chain. Therefore, glycolysis may potentially provide 5 to 7 ATPs that represents 17% to 22% of the total ATP yield (30 to 32 ATPs) per one glucose molecule. This is consistent with the observed rise in oxygen consumption after glucose exchange for pyruvate. A much faster NAD(P)H recovery and larger overshoot in glucose-ACSF indicate a significant contribution of glycolysis-produced NAD(P)H to the total autofluorescence response.
We observed similar effects of pyruvate using the alternative stimulation protocols, i.e., the glutamate puff trains (Figure 5C). Compared with glucose-ACSF, glutamate puff trains in pyruvate-ACSF induced the increase in NAD(P)H oxidation phase, enhancement of oxygen consumption and a decrease in NAD(P)H overshoot. Interestingly, pyruvate affected both the neuronal and astrocytic metabolic parameters in the same manner (Figure 5E).
Altogether, our results suggest that the NAD(P)H overshoot originates mostly from the glycolytic processes in both neurons and astrocytes that proceed in parallel with mitochondrial metabolism and provide a part of energy for covering cellular demands during network activation.
Astrocyte-Released Lactate Is Unlikely a Preferred Neuronal Fuel in Slices
The ‘lactate shuttle’ hypothesis suggests lactate as a preferred energy fuel for neurons during network activation. 14 In such a scenario in slices, neurons should cover their energy needs mainly by using lactate supplied by astrocytes. However, our observations of a robust exogenous pyruvate and lactate action on cell metabolic parameters contradict such a hypothesis. It is possible to suggest that the effects of lactate and pyruvate in our experiments result from the endogenous lactate washout from a slice tissue due to a high rate of perfusion (15 mL/min). This explanation is not valid due to at least three following reasons: (1) since our slices survived well for many hours in glucose-ACSF and showed electrical activity similar to that in pyruvate- or lactate-ACSF, that would mean that glucose was utilized efficiently by cells in the absence of lactate; (2) in studies that used the interface chambers with a very slow ACSF superfusion rate (1 to 2 mL/min) and high level of oxygenation,20, 36 the NAD(P)H waveform in standard ACSF is very similar to that we observed in our experiments in glucose-ACSF; (3) we have shown previously (Ivanov and Zilberter 18 ; see also Figure 4 and Supplementary Figure 7) that in our conditions, not only a complete exchange by but also supplementation of glucose in ACSF with pyruvate or lactate results in a significant reduction in NAD(P)H overshoot.
Therefore, the observation that in slices, exogenous lactate strongly modifies the process of neuronal energy metabolism contradicts the assumption of a preferential neuronal utilization of astrocyte-released lactate. This does not imply, however, that glucose utilization only by itself is optimal for the neuronal functioning. We have shown previously that in both neonatal and adult slices, supplementing glucose in ACSF with pyruvate or lactate improves synaptic function.18, 37
Redox Coupling Between Cytosol and Mitochondria: A Possible Involvement of the Glycerol-Phosphate Shuttle
During glucose utilization for energy generation, NADH produced by glycolysis in cytosol must be reoxidized to NAD+. Since NADH generated by glycolysis cannot penetrate the mitochondrial membrane but should be regenerated to NAD+, shuttling of reducing equivalents from the cytoplasm into mitochondria is necessary for both glycolytic pyruvate formation and lactate transformation to pyruvate. Two main intracellular redox shuttle systems, the glycerol-3-phosphate shuttle and malate–aspartate shuttle, have been described in the brain. 32 The malate–aspartate shuttle is thought to be the major redox shuttle for regeneration of cytosolic NAD+. 32 It transfers the reducing equivalents from cytosolic NADH to mitochondrial NAD+, forming NADH. Opposite to the malate–aspartate shuttle, the glycerol-3-phosphate shuttle transfers the reducing equivalents from cytosolic NADH to mitochondrial FAD leading to the FAD–FADH2 transition. The malate–aspartate is considered as the most important shuttle in brain while the functioning of glycerol-phosphate shuttle has been poorly investigated—although several studies on cultured astrocytes and cerebellar granule cells provided evidence of the shuttle activity in these cells. 32 It has been reported recently that all components of the glycerol-3-phosphate shuttle are expressed in both neurons and astrocytes and suggested that the shuttle should be operational in both cell types. 39 Moreover, it was reported that Ca2+ entry into mitochondria during cell activation inhibited the malate–aspartate shuttle and suggested that the decrease in reducing equivalents' transfer may be compensated by the glycerol-phosphate shuttle activity. 40 Our experiments reveal a strong coupling between the glycolysis-generated NADH and mitochondrial FAD that may be relevant to the activity of glycerol-phosphate shuttle.
In conclusion, our results show that during periods of network activity in slices from mature mice, both neurons and astrocytes are significant consumers of energy produced in parallel by glycolysis and mitochondrial oxidative phosphorylation, and that glucose is an efficient fuel for these processes.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.