
Abstract
While it has been widely confirmed that cerebral blood flow is closely coupled with brain metabolism, it remains a matter of controversy whether capillary flow is directly controlled to meet the energy demands of the parenchyma. Since the capillary is known to lack smooth muscle cells, it has generally been considered that capillary flow is not regulated
Introduction
Roy and Sherrington (1890) hypothesized that the cerebral blood flow (CBF) is intrinsically regulated by products of energy metabolism to adjust it to the altered metabolic demands associated with changes in functional activity. Subsequent findings that CBF is elevated by a rise in P
Since arteries and arterioles in general are rich in vascular smooth muscle cells (SMCs) while true capillaries do not contain any SMCs, arteries and arterioles have been regarded by most researchers as the main vessels involved in the coupling of CBF with metabolic demand (Attwell et al, 2010; Girouard and Iadecola, 2006; Iadecola and Nedergaard, 2007; Paulson et al, 2010; Straub and Nelson, 2007). However, recent reports on pericyte contractility have suggested that CBF may also be controlled at more peripheral vessels. Since the contribution of the capillaries to total vascular resistance is relatively low in the brain, gross CBF should be controlled mainly by the arteries and arterioles (Faraci and Heistad, 1990).
In the following sections, we first broadly review the possible mechanisms of capillary flow control suggested so far and then present our new model based on the previous data. Finally, we consider the physiological and pathological relevance of capillary flow control in clinical settings.
Candidate cellular factors involved in the control of capillary blood blow
Based on their location, the candidate factors involved in the control of capillary flow can be categorized into three components.
Intravascular components: red blood cells (RBCs), leukocytes (neutrophils, macrophages, and lymphocytes), platelets, fibrinogen, and other coagulation factors may affect capillary blood flow. In the microcirculation, the density and velocity of RBCs differ among capillaries. The heterogeneity of intravascular components may result in a difference in blood flow resistance (Pries et al, 1994). ATP and other chemical mediators released from the RBCs, leukocytes, and other vascular components may affect endothelial cells (ECs). Interaction between blood cells and the luminal surface has been suggested to affect capillary flow and may be regulated to control blood flow. Vascular components: ECs and pericytes are the major cellular components involved in the vascular components. Recently, more findings on pericytes have been reported in experimental animals. Vascular SMCs have been found in the arterial capillaries and could control capillary flow in the region upstream of the capillaries. Parenchymal components: Neurons in the parenchyma and astrocytes are reported to control the blood flow at the level of the arteries and arterioles and may also be involved at the capillary level. Microglia may affect capillary flow under pathological conditions. Some neurons have been reported to terminate at the capillaries (Cohen et al, 1997; Kocharyan et al, 2008; Vaucher et al, 2000).
To control the capillary blood flow, at least two apparatuses should be identified: an apparatus to monitor the demand for blood flow (the activity of neurons, hypoxia, hypoperfusion, hypoglycemia, etc.) and a mechanical apparatus to regulate capillary blood flow. These apparatuses have been thought to function in close proximity but may be separated as described in our new model below.
Red blood cells
Tomita
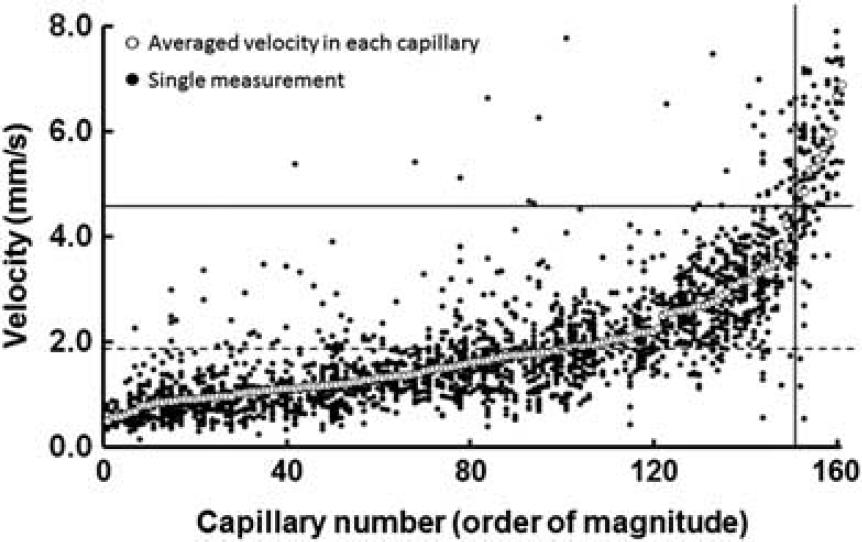
Red blood cell (RBC) velocity measurement in capillaries of the rat cerebral cortex through a closed cranial window. Seven percent of the capillaries showed a much higher RBC velocity (more than mean+2s.d.), suggesting a ‘thoroughfare channel.’ Closed circles: single measurements of RBC velocity. Open circles: average of repeated measurements of RBC velocity in each capillary. Solid horizontal line: mean+2s.d. Dashed line: mean of all capillaries. Solid vertical line: borderline between normal and high-velocity capillaries (Unekawa et al, 2008).
These aspects of heterogeneity in microcirculation may be controlled to change capillary flow under physiological as well as pathological conditions. Using forepaw stimulation, Stefanovic et al (2008) demonstrated that capillaries, defined as small-sized vessels with a diameter <10
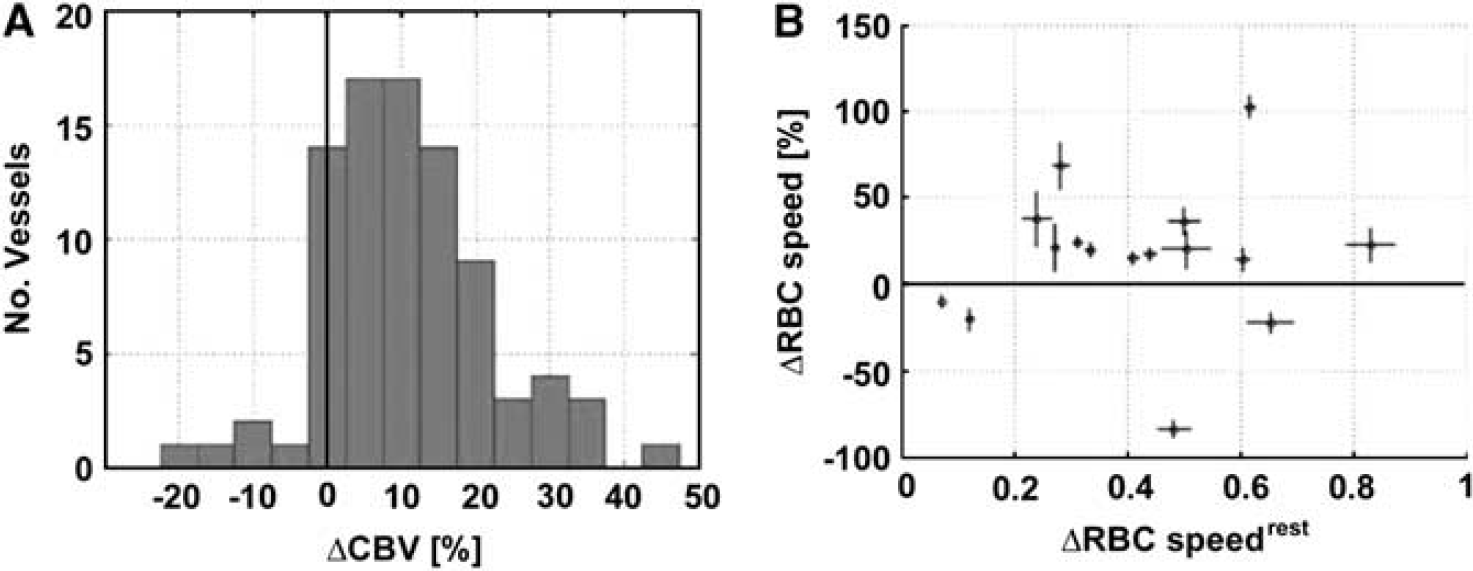
During the function activation, the cerebral blood volume (CBV) increased by 10.9% (
Several authors have proposed that slowly perfused or closed capillaries are functionally recruited upon brain activation (Hertz and Paulson, 1980; Kuschinsky and Paulson, 1992). However,
The average RBC velocity in the capillary was also maintained when the mean arterial pressure was reduced from normal to 60 mm Hg, suggesting the ‘autoregulation’ of RBC velocity (Figure 3) (Hudetz et al, 1995). Since CBF is maintained by the autoregulation, capillary flow and RBC velocity may be preserved as well. During normocapnic hypoxia (Pa
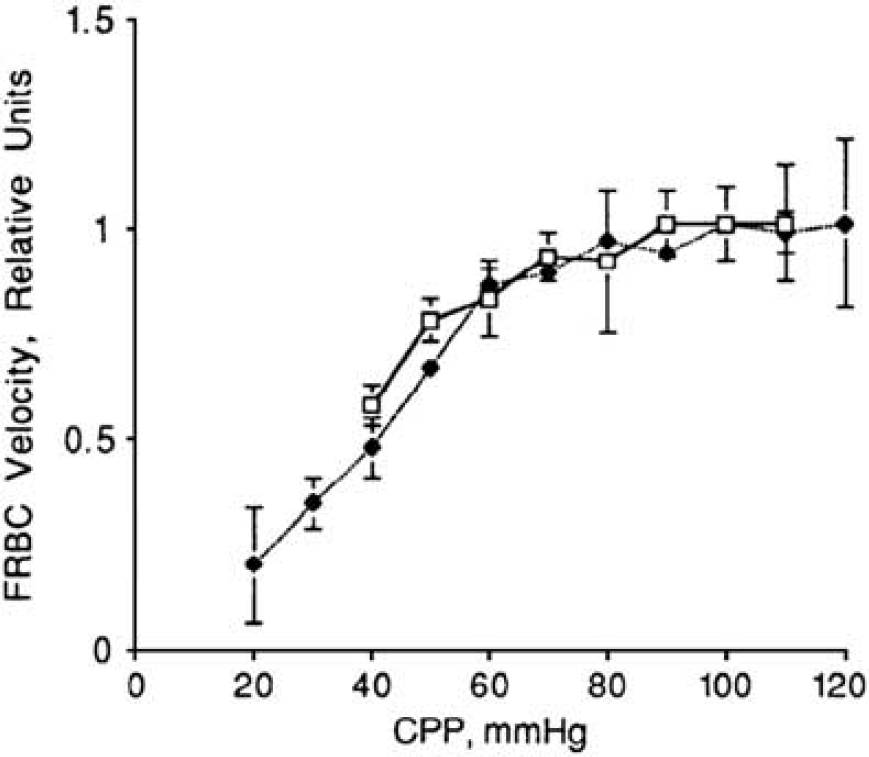
Autoregulation at capillary flow. Systemic blood pressure was decreased by hemorrhagic hypotension (Hudetz et al, 1995).
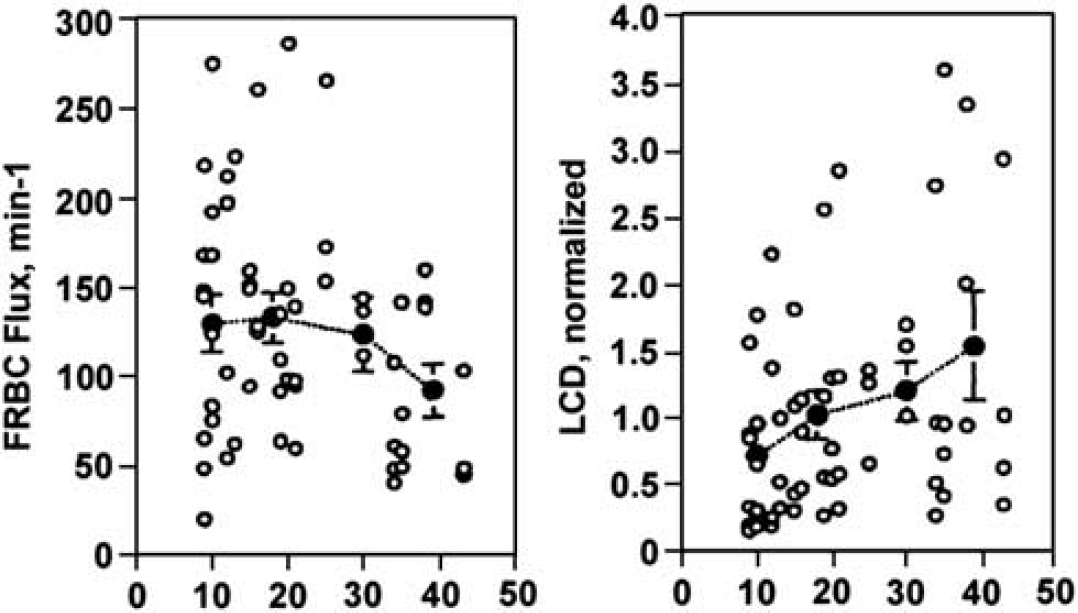
Effect of isovolemic hemodilution on fluorescently labeled red blood cell (FRBC) velocity and FRBC flux. With hemodilution, FRBC velocity increased, whereas FRBC flux was constant (Hudetz, 1997).
In addition to their rheological contribution, RBCs may detect hypoxia and acidosis in ischemic tissue and release ATP, adenosine, and other vasoactive mediators to control capillary flow (Akatsu et al, 2010; Ellsworth et al, 1995; Locovei et al, 2006). This mechanism may function when RBCs are trapped in the thrombus of some systemic organs, but the involvement has not been demonstrated in cerebral capillaries.
Pericytes
Rouget (1873) first identified branched, nonpigmented cells on the capillary wall and regarded them as contractile elements. Vimtrup (1922) observed the tails of young living larvae and noted that ‘the contraction of capillaries begins at one of these cells (pericytes), spreading in both directions, at first slowly, later significantly faster.’ In appreciation of Rouget's work, Vimtrup (1922) termed the observed contractile population at capillaries ‘Rouget cells.’ Since then, the ability of pericytes to regulate capillary flow has been repeatedly discussed. In addition to vascular contraction, pericytes have various functional roles that have been well described in a number of excellent reviews (Allt and Lawrenson, 2001; Balabanov et al, 1999; Dore-Duffy, 2008; Hirschi and D’Amore, 1996; Krueger and Bechmann, 2010; Lai and Kuo, 2005; Thomas, 1999).
Anatomically, the pericytes at the capillary are located between the inner and outer vascular basement membranes, whereas the inner vascular membrane surrounds the endothelium. Astrocytic endfeet are located outside the outer membrane. There is no perivascular (Virchow-Robin) space between the pericytes and astrocytic endfeet at the capillary. The processes of the pericytes, in contrast to those of the SMCs, are oriented along the axis of the blood vessels, while shorter circumferential processes surround the vascular wall (Nehls and Drenckhahn, 1993; Zimmermann, 1923).
In general, the density, morphology, and immunoreactivity to various markers of pericytes are known to differ along the vascular tree (Figure 5) (Nehls and Drenckhahn, 1993), suggesting diverse functions of pericytes based on the location. Boado and Pardridge (1994) studied the cellular distribution of the
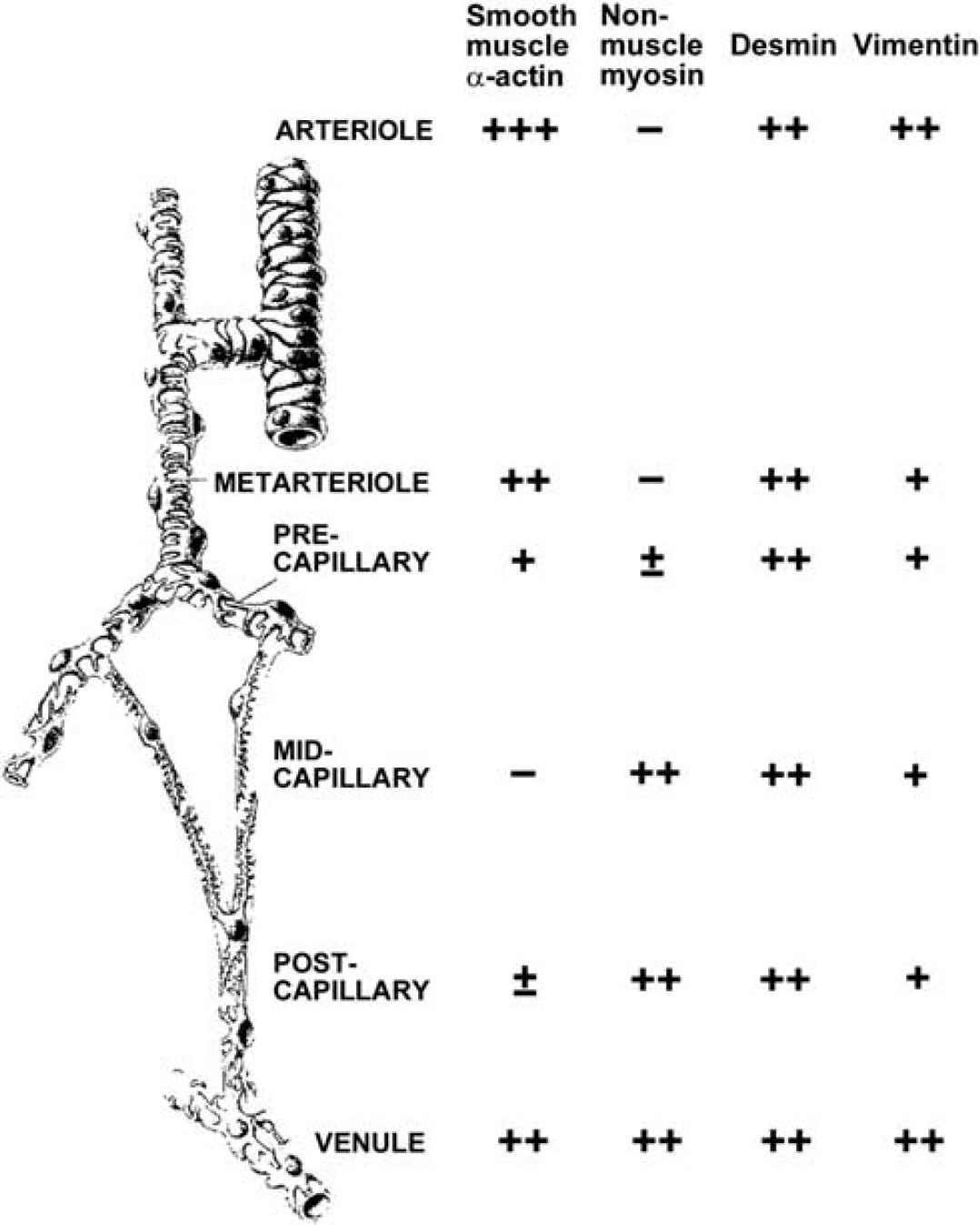
Molecular heterogeneity of pericytes of the rat mesentery, as an example of the microvasculature typical of the peripheral microcirculation. In brain capillaries, even pericytes of pre- and postcapillaries are largely negative for smooth muscle-type actin and most pericytes are negative for desmin (Nehls and Drenckhahn, 1993).
Topical application of vasoactive substances such as nitric oxide, prostacyclin, angiotensin II, endothelin-1, ATP, noradrenaline, and glutamate is reported to induce changes in pericyte contractility (Chakravarthy and Gardiner, 1999; Dehouck et al, 1997; Fernandez-Klett et al, 2010; Healy and Wilk, 1993; Peppiatt et al, 2006). Peppiatt et al (2006) showed that electrical stimulation, superfusion of ATP, and simulated ischemia caused localized constriction of capillaries in the excised whole retina (Figure 6). Since most of these studies were conducted in brain slices without blood flow, sheer stress or perfusion pressure inside the vessels, the vascular reactivity should be further confirmed
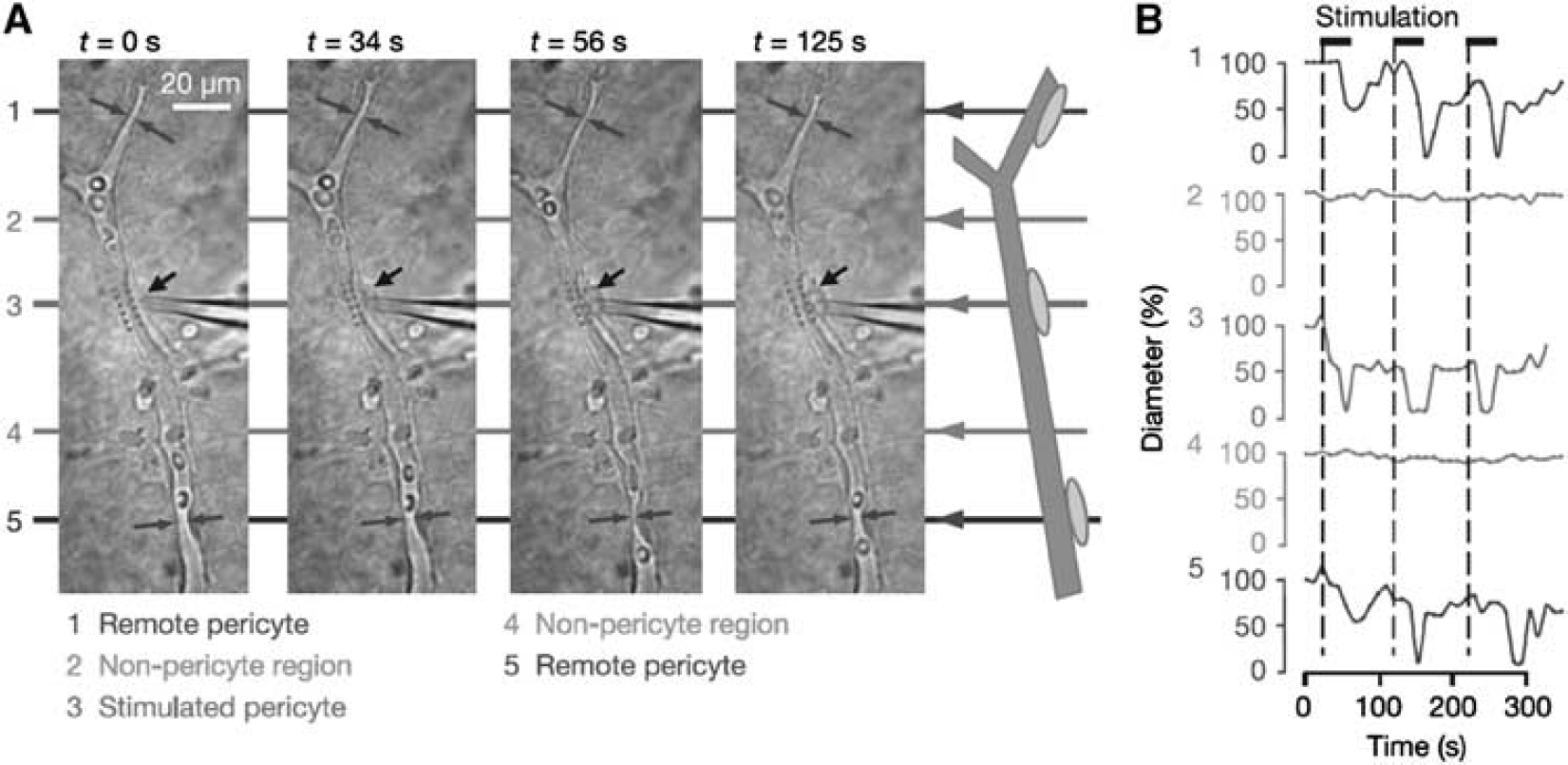
Propagation of retinal pericyte constriction. (
Fernandez-Klett et al (2010) measured the capillary diameter
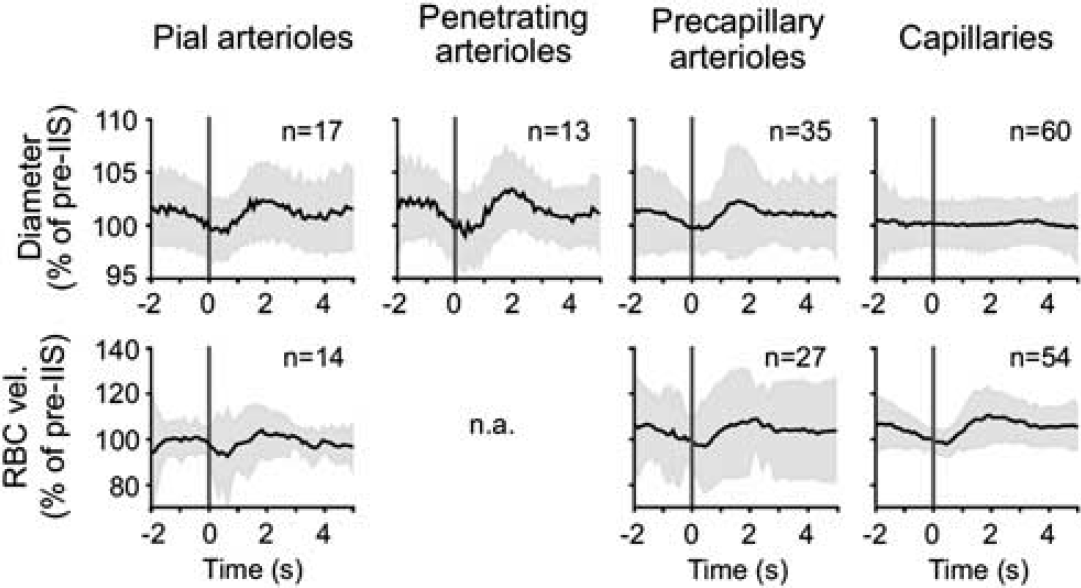
Capillary dilatation does not partake in the hyperemic response induced by bicuculline. Average traces ±s.d. (gray areas) of the pooled diameter (upper) or red blood cell (RBC) velocity responses (lower) of the different vessel types, binned in 200-milliseconds segments (Fernandez-Klett et al, 2010).
Yemisci et al (2009) found numerous capillary constrictions after middle cerebral artery occlusion through the cranial window. Based on this finding, together with the histological examination of fixed brains showing segmental narrowing of capillaries at the site of pericytes and amelioration of the impairment after application of a superoxide scavenger
Another pathological condition in which pericytes are reported to mediate capillary constriction is head trauma. Pericytes have been shown to express both endothelin-1 and its receptors ETrA and EtrB, and the expression of all three factors increased after head trauma. The amelioration of head trauma-induced hypoperfusion by an ETrA antagonist supports the putative function of pericytes (Dore-Duffy et al, 2011).
Endothelial cells
In the brain arteries and arterioles, ECs are widely known to have a pivotal role in controlling CBF by releasing vasoactive mediators to vascular SMCs, whereas their role in the control of capillary flow is not well understood. Without a contractile apparatus, capillary ECs themselves cannot constrict/dilate a capillary, whereas ECs on the precapillary arterioles may control the precapillary sphincter (Anderson and Anderson, 1978; Baez et al, 1977). The precapillary arterioles, defined as vessels <35
One of the main roles of the capillary ECs is to maintain substance exchange across the BBB. Their disturbance under pathological conditions results in vasogenic edema and is known to compress capillaries, disturbing the capillary flow when the brain edema increases intracranial pressure. The ECs themselves become swollen under ischemia and may impede capillary flow in the brain after ischemia/reperfusion, causing a ‘no-reflow’ phenomenon (Chiang et al, 1968). Because the volume of ECs does not change under physiological conditions, they are not involved in the regular capillary flow control.
Damage to the ECs can expose subendothelial connective tissue, prompting platelet thrombosis and disturbing capillary flow. However, under physiological conditions, the involvement of platelets in capillary flow control is unlikely. Endothelial cells on the capillaries and venules may express many types of adhesion molecules and trap various leukocytes (Lindauer et al, 1996). Adhering leukocytes may plug the capillaries, disturbing capillary flow (del Zoppo et al, 1991). Leukocytes adhering on the lumen may also release vasoactive substances or cytotoxic-free radicals and control/damage ECs (Hamann and del Zoppo, 1994). Even though macrophages are reported to patrol over the surface of venules under physiological control, the role of capillary obstruction by leukocytes under physiological conditions is not known.
Endothelial cells are known to respond quickly to changes in sheer stress, thereby detecting changes in capillary flow. Adequate sheer stress is necessary for maintenance of the BBB. It has not been precisely evaluated whether ECs on the capillaries can control capillary flow after detecting changes in the flow. The decrease in shear stress on the endothelium of the arterioles/arteries could trigger dilatation of the large artery (flow-mediated vasodilation) (Tousoulis et al, 2005), but there is limited evidence supporting this possibility in the neocortical microcirculation.
Astrocytes
Increased activity of neurons, especially at the synapses, induces Ca2+ elevation in neighboring astrocytes. These astrocytes with their endfeet extending to the outer vascular membrane release vasoactive substances to control the CBF. Neurons, astrocytes, and vessel components are now regarded as a functional complex, a neurovascular unit. The neurovascular unit is widely reported to dilate/constrict vascular SMCs at the arteries/arterioles but not at the capillaries (Girouard and Iadecola, 2006; Haydon and Carmignoto, 2006; Takano et al, 2006).
Proximal integration model
As discussed above, the gap between the location of the metabolites exchange (capillary area) and that of the blood flow control (precapillary and upstream arterioles) has been a longstanding enigma. Here, we postulate a new model in which the control of the capillary flow is based on peripheral signals. We name this the ‘proximal integration model,’ since the integration of the vasoactive signals that are generated around the capillaries and transmitted to the proximal arterioles forms the basis of this hypothesis.
Exchange of substances is solely conducted at the capillaries. Therefore, changes in neural activity must first be detected at the capillaries around which the metabolism is enhanced. Then capillaries can transmit these signals to modify smooth muscle tone up to the precapillary arterioles (Figure 8), analogous to dendritic integration on a neuron. In addition to the SMCs, pericytes at the arterioles may contribute to the regulation of vascular tonus. Dilatation of the precapillary arterioles increases blood flow to the activated area, and signals from the area may be lowered accordingly. The integrated signals at the precapillary arterioles may further be transmitted to the upstream penetrating artery and finally to the pial artery if the integrated signal intensity is large enough (Girouard and Iadecola, 2006). In this model, redistribution of blood flow may be conducted at the branching point of capillaries based on the signals from the periphery, selectively increasing blood flow to the area of neuronal activation while preserving blood flow to the resting area (Figure 9).
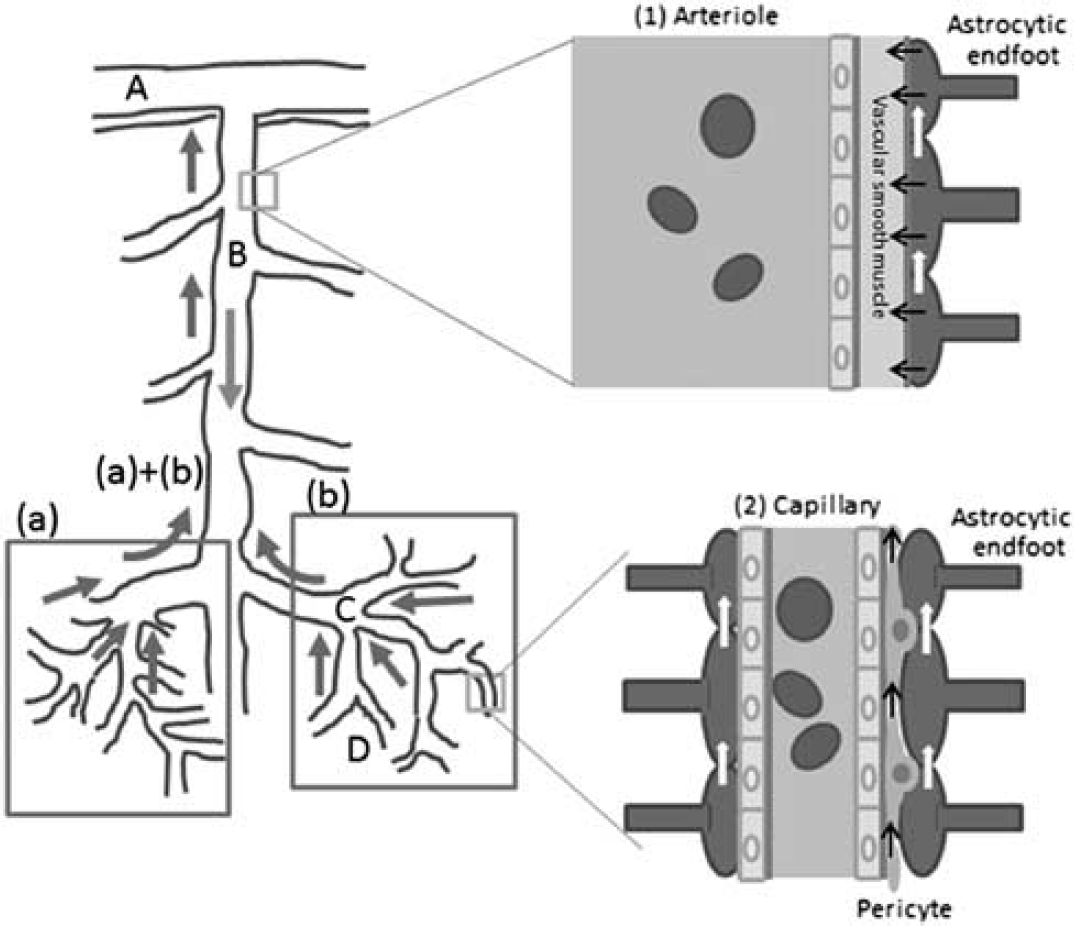
Proximal integration of signals from the areas of neural activation. The arteriole responds to the summation of peripheral signals at each point. ‘A’ denotes pial artery; ‘B’ denotes penetrating artery; ‘C’ denotes precapillary artery; and D denotes true capillary. (a, b) Denote areas of neural activation; (a)+(b) denote integrated signals from areas (a) and (b); red arrow: blood flow; blue arrow: signal conductance of neural activation to the proximal arteriole/artery. (1) Blood flow at an arteriole is controlled with signals from astrocytic endfeet (black arrow), via changes in the tone of the vascular smooth muscles. Signals at the endfeet are the integration of those from the periphery (white arrows). (2) Capillary blood flow may partly be controlled with constriction of local pericytes but is mainly determined by the blood flow coming from the precapillary artery, which is controlled by the signals from the periphery through pericytes (black arrows) or astrocytic endfeet (white arrows). The color reproduction of this figure is available on the
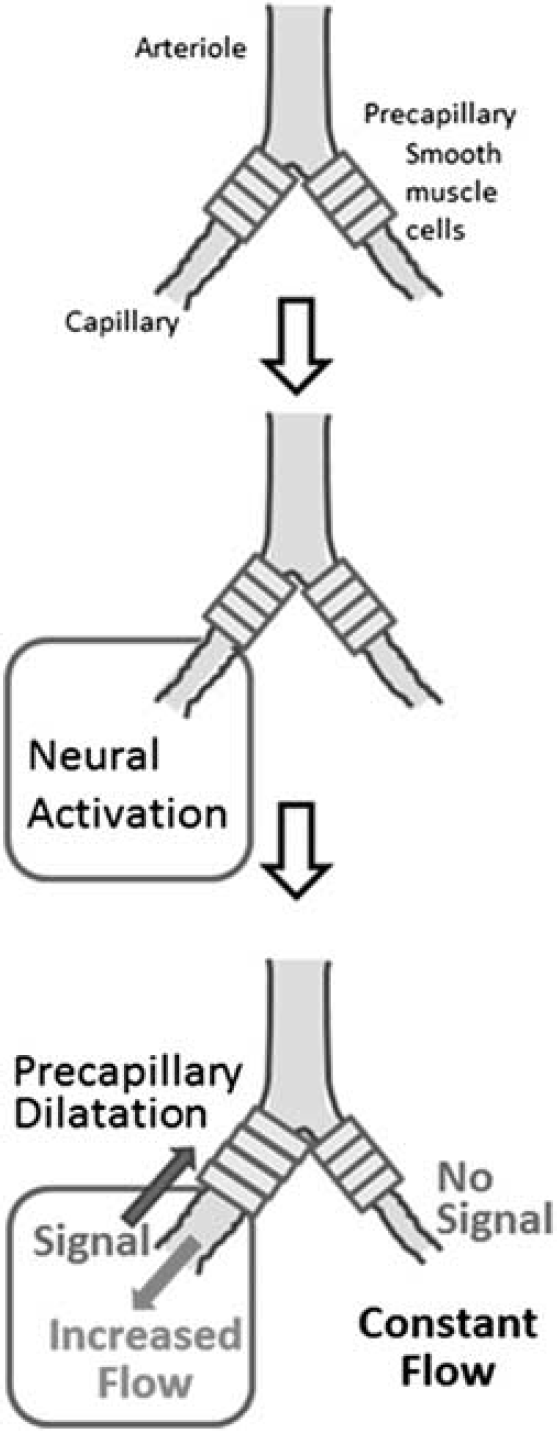
Control of capillary flow. The signal of neural activation at the periphery is transmitted through astrocytic endfeet or pericytes to the smooth muscle at the precapillary arteriole. Dilatation of smooth muscle increases blood flow down to the capillary, whereas blood flow to other branches remains constant.
The candidate cells for signal transduction/integration are pericytes and astrocytes. Pericytes are inside of the outer basement membrane, and astrocytic endfeet surround the outer membrane of the capillary (Simard et al, 2003). Pericytes at the retinal microvasculature have been reported to form the electrical syncytium and to transmit signals through gap junctions (Puro, 2007). Peppiatt et al (2006) demonstrated in a brain slice that local electrical stimulation as well as topically applied chemicals induced constriction of neighboring pericytes, perhaps through gap junctions. A pericyte on the true capillary is morphologically oriented along the capillary axis and may be suitable for signal transduction along the vessel (Nehls and Drenckhahn, 1993).
Astrocytic endfeet are also connected through gap junctions to form the syncytium. Simard et al (2003) showed that Lucifer yellow injected into an astrocyte diffused preferentially along the vessel wall through gap junctions between the endfeet on the vessel wall. The coordination of astrocytes with pericytes for the integration of vasomotor signals needs to be tested further.
In the muscle microcirculation, the intercellular conduction of signals between neighboring ECs or between vascular SMCs via gap junctions was suggested (Segal, 2005). As in flow-mediated dilatation at the arteries/arterioles, ECs on the capillaries may detect changes in shear stress and transmit the signal through the gap junction between ECs to upstream arteries/arterioles, where the blood flow may be controlled. However, there is limited evidence supporting this as a candidate mechanism for the control of capillary flow.
A somewhat similar model was suggested previously. Based on the observation of the microcirculation, Krogh et al (1922) postulated proximal control of the blood flow to the periphery in the brain. Woolsey et al (1996) analyzed anatomical patterns of the vascular and neural network and hypothesized retrograde propagation of vasoactive signals and upstream control of blood flow. In fact, retrograde vasodilation in response to ATP has been demonstrated in isolated rat cerebral arterioles (Dietrich et al, 1996). In addition, upstream vasodilation was observed in cerebellar cortex arterioles during activation of the parallel fibers (Iadecola et al, 1997). Analysis of the temporal profiles of the vessel constriction may elucidate the pathway of signal transmission.
Neuronal control
Anatomically, arterioles are innervated with extrinsic/intrinsic neurons, though the significance of the neural control of blood flow under physiological conditions as well as pathological conditions is not fully understood. Intrinsic neurons directly attaching to the vessel walls are reported even down to the capillary level (Cohen et al, 1997; Kocharyan et al, 2008; Vaucher et al, 2000). Since pericytes have some receptors for neurotransmitters (Rucker et al, 2000), neurons around capillaries can activate pericytes and may influence capillary flow, though so far there is no direct evidence supporting this hypothesis.
In addition, Takano et al (2007) reported that during the CSD, neurons around capillaries can be swollen and may compress capillaries. Compared with the brain edema after ischemia, the volume of the swollen neurons is too small to compress capillaries effectively.
Clinical relevance of capillary flow control
(1) Functional activation: Coupling of CBF with metabolism is one of the essential mechanisms to support increased neuronal activity (Roy and Sherrington, 1890). Local activation of neurons should be accompanied with increase in capillary flow in the corresponding area (Stefanovic et al, 2008). The brain is known to redistribute CBF based on the local neural activity constantly and promptly. Therefore, dysfunction of capillary flow control may result in relative ischemia and neural dysfunction. However, cerebral function requiring increased neural activity is clinically indistinguishable from basic neural function. Unlike the heart during angina, the brain does not develop apparent ischemic symptoms even when the reactive flow increase is limited, since the increase in energy demand is smaller than that of the heart (Tune et al, 2004). Mild dementia, slow in response and mild clumsiness in the extremities may be symptoms of limited capillary flow activation.
Clinically functional activation of local flow is widely utilized for the acquisition of functional magnetic resonance imagings. Since neural activation in a limited area is suggested to increase capillary flow in the corresponding area, precise mapping can be obtained by the functional magnetic resonance imaging. In contrast, positron emission tomography can directly measure the increased metabolism with less spatial resolution.
(2) Vascular disease: Atherosclerosis mainly affects the major arteries, but the orifice of the penetrating artery may also be affected. Hypertension is a main risk factor for lipohyalinosis of the small penetrating artery. Diabetic angiopathy affects the major arteries as well as the small arteries. Cerebral angitis can be accompanied with various collagen diseases. These vasculopathies may affect capillary flow control and induce limitations in neural activation. However, capillary flow dysregulation is clinically indistinguishable from mildly reduced basal flow or encephalopathy caused by the diseases and remains just one of the putative mechanisms involved in cerebral dysfunction in these diseases.
(3) Epilepsy: Using bicuculline-induced epilepsy, Fernandez-Klett et al (2010) reported that epilepsy-induced hyperemia was mediated by arterioles without the involvement of capillary dilatation. In this paper, an increase in RBC velocity in the capillaries was regarded as secondary to a proximal increase in RBC velocity. Constant vessel diameter with increase in blood velocity may suggest increase in capillary flow in their experiment. In addition to the local metabolic enhancement, epileptic activity may spread to the proximal artery and dilate it as well.
(4) Cortical spreading depression: Fernandez-Klett et al (2010) observed an increase in capillary and arteriolar diameter during CSD. In their report, changes in the diameter of capillaries and arterioles were synchronous. Takano et al (2007) reported relative hypoxia and neural swelling, suggesting that increased oxygen consumption with a limited supply caused relative hypoxia. Changes in metabolism and control of local flow have not been fully elucidated in CSD. Cortical spreading depression may have some role in the occurrence of migraine aura. Capillary flow disturbance may be a treatment target during CSD to decrease the duration of auras and to prevent a headache attack after an aura occurs.
(5) Ischemia and hypoxia: Yemisci et al (2009) reported that pericyte contraction impairs capillary reflow after reopening of the cerebral artery. Multiple thrombus formation, endothelial swelling, leukocyte plugging, and BBB breakdown followed by brain edema are other factors reported to be involved in the disturbance of the capillary flow. Improvement in microcirculation is now regarded as one of the main targets of acute ischemia.
Acute hypoxia/ischemia is associated with capillary dilatation. The proximal integration model can typically be applied to the pathophysiology. That is, based on the affected area, multiple levels of the arteries may be involved in the recovery of CBF. When the hypoxia/ischemia is severe, larger arteries would be involved.
In contrast, chronic ischemia can induce capillary remodeling (histological reorganization of vessels) to meet the prolonged demand of energy and oxygen (Guo et al, 2011). Both pericytes and ECs are considered to be involved in this capillary remodeling (Krueger and Bechmann, 2010; Rucker et al, 2000).
(6) Autoregulation of capillary flow has been reported (Hudetz et al, 1995). The brain is known to be protected against changes in blood pressure. Autoregulation found in the capillaries may mostly be based on preserved total CBF, though local adjustment of blood flow may also be activated with blood pressure outside the range of autoregulation. Clinically, autoregulation is important for maintaining CBF during hypotension as well as hypertension.
(7) Head trauma: Dore-Duffy indicated that pericyte-mediated vasoconstriction underlies head trauma-induced hypoperfusion (Dore-Duffy et al, 2011). In addition to the direct damage of head trauma, the impaired capillary flow may become a treatment target after head trauma.
As shown above, the impaired capillary flow control may be an important pathophysiological factor and treatment target in many diseases.
Future directions
Recent technologies, including two-photon microscopy and green fluorescent protein (GFP)-labeled vascular components, have enabled capillary flow in the deeper cortex to be visualized
Footnotes
Disclosure/conflict of interest
The authors declare no conflict of interest.