
Abstract
Crosstalk among brain endothelial cells (BECs), pericytes, and astrocytes occurs by way of soluble factors, including cytokines. Here, we studied cytokine secretion from both mouse BEC monocultures and tri-cultured with pericytes and astrocytes. Four cytokines were constitutively secreted by BEC monolayers, 12 by LPS-stimulated BECs, 10 by tri-cultures, and 14 by LPS-stimulated tri-cultures. Cytokine levels were generally higher with either LPS stimulation or tri-culture when compared to monocultures and highest in tri-cultures stimulated by LPS. LPS-stimulated secretions fell into eight patterns as categorized by the polarization of cytokine secretions. To determine the cellular origin of cytokine increases in tri-cultures, we cultured mouse BECs with human pericytes and astrocytes and measured cytokines in species-specific assays. Thus, cytokines detected in the human immunoassay were from pericytes/astrocytes and those detected in the mouse immunoassay were from BECs. Several unique patterns were thus found. For example, TNF-alpha was only of pericyte/astrocyte origin; granulocyte colony-stimulating factor was only of BEC origin; IL-6, MCP-1, and GM-CSF of astrocyte/pericyte origin were found in both the luminal and abluminal chambers, suggesting the presence of brain-to-blood transporters. We conclude that crosstalk influences cytokine secretion under constitutive and stimulated conditions from both BECs and pericytes/astrocytes.
Introduction
The endothelial cells lining the arterioles and venules and forming the capillary bed of the central nervous system (CNS) are modified to form the vascular blood–brain barrier (BBB). These modifications include those which result in barrier function: the formation of tight junctions; the elimination of fenestrae; a reduction in macropinocytosis.1,2 Additionally, the modifications include the induction of transporters, enzymatic activities, and receptors unique to or enriched in the BBB. 3 These modifications of the BBB result from its cross-talk with the other cells of the CNS in a formation termed the neurovascular unit (NVU). 4 These other cells include the neurons, microglia, mast cells, but most especially the astrocytes and pericytes. Features of the BBB are first induced in the fetus by pericytes and, later, by astrocytes.5,6
Understanding this cross-talk and its mediators is critical to understanding how the barriers function both as protective to the CNS and in its roles as a blood-brain interface. Additionally, elucidation of cross-talk agents and mechanisms has already been shown to be important in understanding disease mechanisms, such as the role of pericytes in BBB functioning in Alzheimer’s disease, AIDS, and diabetes mellitus.7–9 Finally, understanding cross-talk will provide unique targets for the development of CNS therapeutics. 10
The mediators of this cross-talk are largely unknown. However, immune-active agents such as nitric oxide, prostaglandins, and cytokines are secreted by brain endothelial cells (BECs), pericytes, astrocytes, and other cells of the NVU.11–13 These agents are clearly involved in the functioning of the BBB and NVU cross-talk. For example, interleukin-6 (IL-6) and granulocyte macrophage colony-stimulating factor (GM-CSF) secreted from BECs mediate the LPS-enhanced passage of HIV-1 across the BBB. 14 This passage is further enhanced by the presence of pericytes with the enhancement possibly mediated by the pericyte secretion of keratinocyte chemoattractant (KC) and monocyte chemoattractant protein-1 (MCP-1). 7
The cross-talk among the various cells of the NVU is likely to result in the modification of the secretion of their cytokines. But this cross-talk is likely complex and its study is difficult, requiring unique tools. Here, we used the Transwell monolayer culture method to study the interactions among BECs, astrocytes, and pericytes in terms of barrier function and cytokine secretion. By culturing mouse BECs with human astrocytes and pericytes and then using species-specific immunoassays, we were able to determine the cellular source of cytokines and so the influence of one cell type on the secretions of another. We studied both basal secretions and LPS-induced secretions in monocultures (BECs alone) and tri-cultures (BECs, astrocytes, and pericytes). LPS is a powerful inducer of cytokine release from BECs, pericytes, and astrocytes.11,12,15 and also alters many other functions of the BBB, including altering BBB integrity, immune cell trafficking, and transporter functions.16–19 Because the BBB is a polarized layer and pericytes and astrocytes are only on the abluminal side, we measured cytokine secretions in luminal and in abluminal chambers and the effects of LPS introduced either into the luminal or abluminal chambers. We were thus able to characterize the individual secretion patterns and NVU interactions for 15 cytokines.
Material and methods
Mouse and human glial cell cultures
Mouse mixed glial cell cultures were prepared according to McCarthy and de Vellis. 20 Cerebral cortices of newborn mice (0–1 days old) were dissected, stripped of their meninges and mechanically dissociated by repeated pipetting followed by passage through a nylon mesh. Cells were plated onto poly-L-lysine (10 ug/ml; Sigma-Aldrich Co., St. Louis, MO, USA) pre-coated plastic and cultivated in DMEM containing 10% FCS, and 1% GlutaMAX (Gibco) at 37℃, 5% CO2 in water saturated atmosphere. Medium was changed twice a week. Cultures reach confluence after 8–10 days in vitro and were used between 14 and 20 days in vitro. Primary human astrocytes were obtained from ScienCell Research Laboratories (Carlsbad, USA) and cultured according to the manufacturer’s protocol.
Mouse and human brain pericytes culture
Primary mouse brain pericytes were prepared according to Nakagawa et al. 21 Briefly, the cultures of mouse cerebral pericytes were obtained by a prolonged, two-week culture of isolated brain microvessel fragments that contain pericytes and endothelial cells. Pericyte survival and proliferation were favored by selective culture conditions, using uncoated dishes, and DMEM F12 supplemented with 20% fetal calf serum (Sigma-Aldrich Co., St. Louis, MO, USA), 1% GlutaMAX (Gibco) and gentamicin (Sigma-Aldrich Co., St. Louis, MO, USA). Culture medium was changed twice a week. Primary human pericytes were obtained from ScienCell Research Laboratories (Carlsbad, USA) and cultured according to the manufacturer’s protocol.
Mouse brain endothelial cultures
Primary mouse BECs were prepared according to Coisne et al. 22 with modifications. Briefly, meninges were carefully removed from forebrains and gray matter was minced into small pieces. Preparations were pooled and ground with a Dounce homogenizer in Dulbecco's Modified Eagle's Medium/Nutrient Mixture F-12 Ham (DMEM/F12; Sigma-Aldrich Co., St. Louis, MO, USA) supplemented with gentamicin (50 µg/ml, Sigma-Aldrich Co., St. Louis, MO, USA). The resulting homogenate was mixed with 30% dextran (v/v, molecular weight 100,000–200,000, Sigma-Aldrich Co., St. Louis, MO, USA) in DMEM/F12 supplemented with 0.1% bovine serum albumin (BSA; Sigma-Aldrich Co., St. Louis, MO, USA). The suspension was centrifuged at 3000 × g for 25 min at 4℃. The pellet was suspended in DMEM/F12, and the supernatant was centrifuged again under same conditions. After the second centrifugation, the supernatant was discarded and the pellet was re-suspended in DMEM/F12. Then the pellets were filtered through a 70 µm nylon mesh and digested in collagenase/dispase (2 mg/ml, Roche Applied Science, USA) and DNase I (10 µg/ml, Sigma-Aldrich Co., St. Louis, MO, USA) for 30 min. The digested solution was filtered through a 20 µm nylon mesh and seeded on with collagen type IV/fibronectin-coated dishes (both form Sigma-Aldrich Co., St. Louis, MO, USA). Cultures were maintained in medium composed of DMEM/F12 supplemented with 10% plasma-derived serum (PDS, Animal Technologies, Inc., USA), 1% GlutaMAX (Gibco), basic fibroblast growth factor (bFGF; Roche Applied Sciences), heparin, insulin, transferrin and sodium selenite supplement and puromycin (4µg/ml, Sigma-Aldrich Co., St. Louis, MO, USA). Twenty-four hours after plating, red blood cells, cell debris, and nonadherent cells were removed by washing with medium. On the third day, the puromycin was removed from medium. When the cultures reached 80% confluency (5th day in vitro), the purified endothelial cells were passaged by brief treatment with 0.25% Trypsin-EDTA (Invitrogen, Life Technologies), and used to construct in vitro BBB models.
Tri-cultures and cell stimulation
Primary mouse brain pericytes or human brain pericytes (15,000 cells/cm2) were seeded on the bottom of the collagen-coated polyester membrane (0.33 cm2, 0.4 µm pore size) of the Transwell® inserts (24-well type, Sigma-Aldrich Co., St. Louis, MO, USA). The pericytes were allowed to adhere firmly overnight. Endothelial cells (150,000 cells/cm2) were seeded the next day on the inside of the inserts (Figure 7). The inserts were then placed in the well of the 24-well plate cultured with astrocytes. BBB models were maintained in DMEM/F12 supplemented with 10% plasma-derived serum (PDS, Animal Technologies, Inc., USA), 1% GlutaMAX (Gibco), basic fibroblast growth factor (bFGF; Roche Applied Sciences), heparin, insulin, transferrin and sodium selenite supplemented with hydrocortisone (500 nM; Sigma-Aldrich Co., St. Louis, MO, USA) at 37℃ in a humidified atmosphere of 5% CO2 and 95% air. Experiments were carried out three or four days after endothelial cells were seeded. For stimulation, experiment cells were washed with serum-free medium, and exposed to culture media with or without lipopolysaccharide (LPS) from Salmonella typhimurium (L6511; Sigma-Aldrich Co., St. Louis, MO, USA) at 1 µg/ml for 24 h.
Transendothelial electrical resistance and permeability of markers
Transendothelial electrical resistance (TEER in Ω × cm2) was measured using an EVOM resistance meter (World Precision Instruments, Sarasota, FL, USA). The TEER of cell-free inserts was subtracted from the measured values. For the transport experiments, the medium was removed and inserts were washed with Krebs-Ringer-Phosphate-HEPES (KRPH) buffer containing 1% BSA (141 mM NaCl, 4.0 mM KCl, 2.8 mM CaCl2, 1.0 mM MgSO4, 1.0 mM NaH2PO4, 10 mM HEPES, 10 mM D-glucose and 1% BSA, pH 7.4). The KRPH buffer containing 1% BSA was added to the abluminal chamber of the Transwell® insert. To initiate the transport experiments, 125I-Albumin (labeled by chloramine T method; 5 × 106 cpm/ml) or 14C-Sucrose (1.5 × 106 cpm/ml; Perkin Elmer, USA) was loaded on the luminal chamber. Samples were removed from the abluminal chamber at 10, 20, 30 and 45 min and immediately replaced with an equal volume of fresh 1% BSA/KRPH buffer. 125I-Albumin samples were precipitated with TCA and the radioactivity was determined using a gamma counter. Radioactivity of 14C-Sucrose was measured in a liquid scintillation counter (Tri Carb 1900, Packard Instrument Company, Perkin-Elmer Life Sciences, Courtaboeuf, France). The permeability coefficient and clearance of 125I-Albumin and 14C-Sucrose was calculated according to the method described by Dehouck et al.23
Clearance was expressed as microliters of radioactive tracer diffusing from the luminal to abluminal chamber and was calculated using the initial amount of radioactivity in the loading chamber and the measured amount of radioactivity in the collected samples.
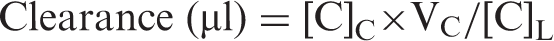
The volume cleared was plotted versus time, and the slope was estimated by linear regression analysis. The slope of clearance curves for the BMEC monolayer plus Transwell membrane was denoted by PSapp, where PS is the permeability × surface area product (in microliters per minute). The slope of the clearance curve with a Transwell membrane without BMECs was denoted by PSmembrane. The real PS value for the BMEC monolayer (PSe) was calculated from
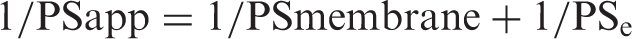
The PSe values were divided by the surface area of the Transwell inserts to generate the endothelial permeability coefficient (Pe, in microliters per minute per square centimeter).
ELISA cytokines
Concentrations of mouse and human cytokines and chemokines secreted to the culture media were measured by commercial magnetic bead-based Multiplex ELISA kit (Bioplex, Biorad, USA) according to the manufacturer’s protocol. Of the 23 cytokines in the mouse kit, we detected 15 different cytokines: Interleukin-1 alpha (IL-1 alpha), IL-1 beta, IL-6, IL12(p40), IL-12(p70), IL-13, granulocyte colony-stimulating factor (G-CSF), GM-CSF, eotaxin, KC, monocyte chemotactic protein 1 (MCP-1), macrophage inflammatory protein-1 alpha (MIP-1 alpha), MIP-1 beta, chemokine (C–C motif) ligand 5 (RANTES), and tumor necrosis factor alpha (TNF-alpha). The 17-plex was used to measure human cytokines and had 10 cytokines in common with the mouse kit: IL-1 beta, IL-6, IL-12(p70), IL-13, G-CSF, GM-CSF, MCP-1, MIP-1 beta, TNF-alpha, and IL-8, the human equivalent of murine KC.
Immunocytochemistry
Cells on inserts were washed in PBS and fixed with 4% PFA for 10 min at 4℃. Cells were permeabilized with 0.2% TRITON-X100, blocked with 5% BSA and then incubated with anti- claudin-5, ZO-1 and occludin antibodies (all from Thermo Fisher Scientific, USA), followed by incubation with corresponding ALEXA Fluor-488 conjugated secondary antibody (Thermo Fisher Scientific, USA). Finally, the membranes were cut out of the inserts, mounted in fluorescence mounting media and photographed with a Nikon ECLIPSE E8000 fluorescence microscope.
Data analysis
All experiments were done in triplicates and repeated once (n = 6). Values are presented as the means ± SEM. Statistical analysis was performed using a two-way ANOVA (GraphPad Prism). Newman–Keul’s multiple comparison tests was used for post hoc comparison. Differences at P < 0.05 were accepted as statistically significant.
Results
Effect of mouse pericytes and mouse astrocytes on tight junction proteins and permeability
Both monocultures (mouse endothelial cells) and tri-cultures (mouse endothelial cells, mouse pericytes, and mouse astrocytes) showed the appearance of the tight junction proteins claudin-5, ZO-1, and occludin. However, tri-cultures tended to have more of the claudin-5 and ZO-1 located at inter-endothelial interfaces and less within cytoplasm (Figure 1).
Immunohistochemistry of the tight junction proteins claudin-5, ZO-1, and occludin. Upper panels (a–c) are monocultures of mouse brain endothelial cells (EOO) and lower panels (d–f) are tri-cultures of mouse brain endothelial cells, mouse pericytes, and mouse astrocytes (EPA). Both monocultures and tri-cultures expressed tight junction proteins. Tri-cultures tended to have more of the tight junction proteins located at the intercellular interface and less in cytoplasm.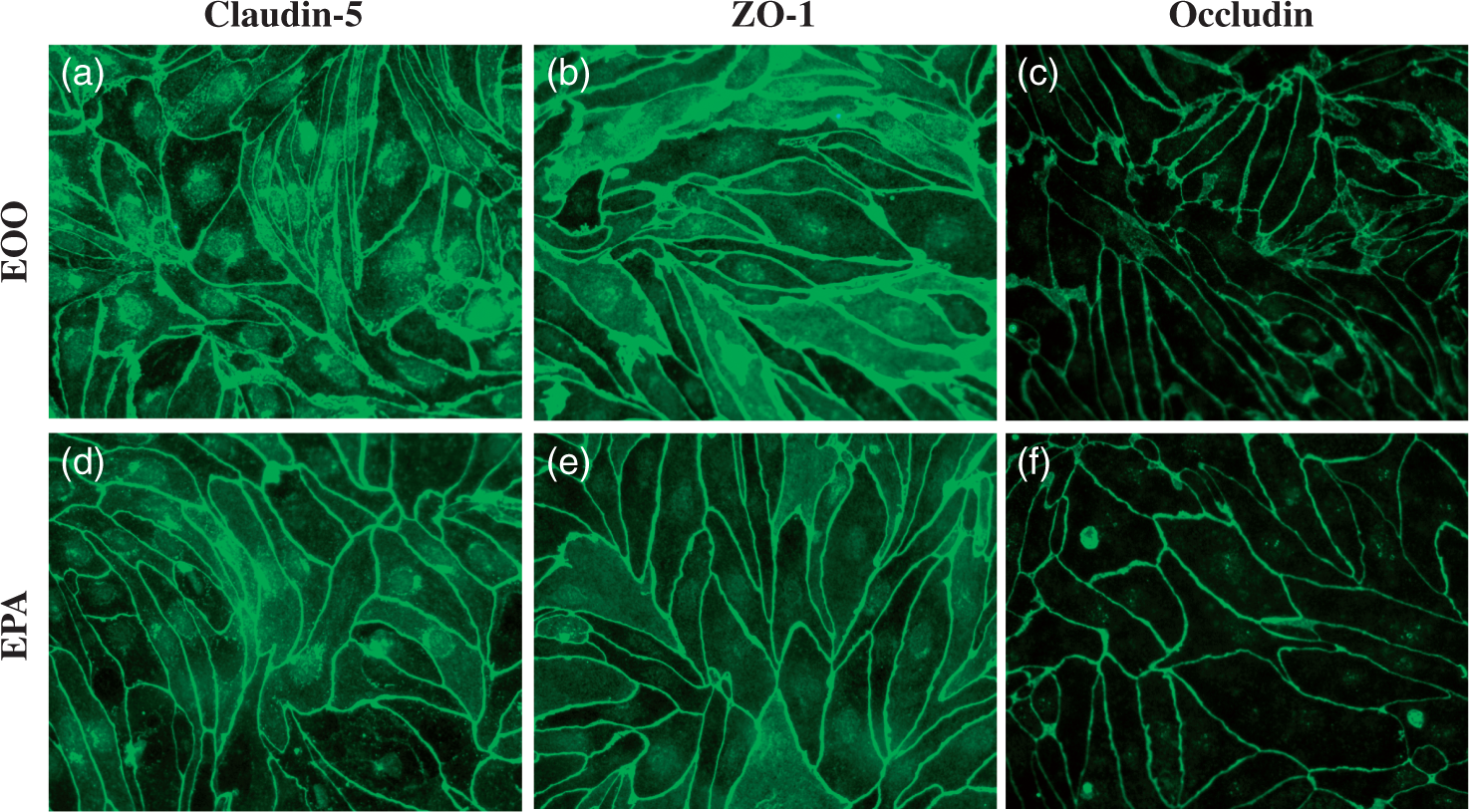
Only tri-cultures significantly increased TEER as measured in Figure 2(a); co-cultures with either pericytes alone or astrocytes alone did not increase TEER to a statistically significant level. Permeability to 14C-sucrose was decreased when co-cultured with astrocytes, and even more so when co-cultured with pericytes or pericytes plus astrocytes (Figure 2(b)). Co-culture with pericytes was as effective at decreasing permeability to 14C-sucrose as tri-culture. Permeability to I-Albumin was not affected with co-culture or tri-culture (Figure 2(c)).
Effects of astrocytes and pericytes on brain endothelial cell monolayer permeability. Panel A shows that only tri-culture with both pericytes and astrocytes improved TEER; n = 5–6; ANOVA: F(3,18) = 11.6, p < 0.001; Newman–Keuls: ***p < 0.001 EOO vs EPA. Panel B shows that as measured by leakiness to sucrose, astrocytes improved barrier function and pericytes even more so. Adding both pericytes and astrocytes was not more effective than culturing with pericytes only; n = 3–4; ANOVA: F(3,10) = 52.4, p < 0.001; Newman–Keuls: *p < 0.05, ***p < 0.001. Panel C shows that neither astrocytes nor pericytes improved BBB leakiness as measured with albumin. Panel D: Mouse brain endothelial cells were tri-cultured with mouse or human pericytes and astrocytes. Pericytes/astrocytes from either species greatly increased TEER, although mouse pericytes/astrocytes were statistically more effective. LPS dramatically disrupted barrier as evidenced by a decrease in TEER regardless of presence or species of pericytes/astrocytes; ANOVA: F(8,119) = 174, p<<0.001; Newman–Keuls: **p < 0.01; ***p < 0.001. EOO: Mouse brain endothelial cell monocultures; EOA: Mouse brain endothelial cells co-cultured with mouse astrocytes; EPO: Mouse brain endothelial cells co-cultured with mouse pericytes; EPA: Mouse brain endothelial cells tri-cultured with mouse pericytes and mouse astrocytes; E + hPA: Mouse brain endothelial cell tri-cultured with human pericytes and human astrocytes.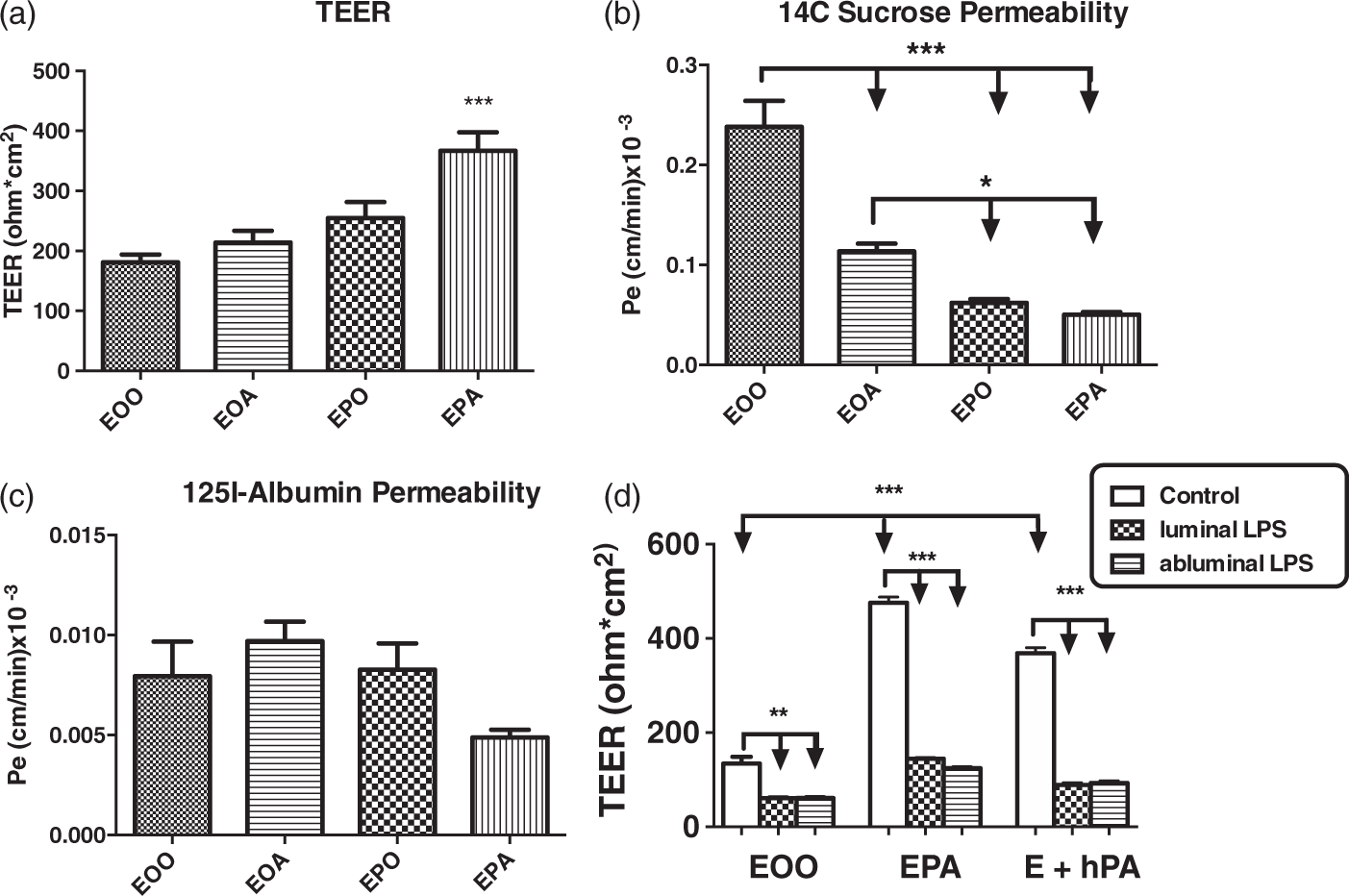
Effect of human pericytes/astrocytes vs mouse pericytes/astrocytes on TEER
Pericytes and astrocytes of either human or mouse origin significantly improved the permeability of monolayers of mouse endothelial cells, although cells of murine origin were somewhat superior (Figure 2(d)). LPS reduced TEER of the monoculture and of the tri-cultures. LPS was equally effective in reducing TEER in tri-cultures with human or with murine pericytes/astrocytes and when placed in either the luminal or abluminal chambers.
Mouse pericytes/astrocytes and cytokine secretions
Of the 23 cytokines assayed, 4 were spontaneously secreted in monocultures of mouse BECs: G-CSF, KC, MCP-1, and RANTES (Figures 3 and 6). These were secreted into both the luminal and abluminal chambers. In tri-cultures of BECs, murine pericytes, and murine astrocytes, these four cytokines were still secreted, although generally in somewhat higher amounts. In addition, six other cytokines were secreted: IL-6, IL-13, TNF-alpha, eotaxin, MIP-1 alpha, and MIP-1 beta. For these six cytokines, abluminal levels were generally higher than luminal levels with the exception of eotaxin.
Constitutive secretion of cytokines by monocultures and tri-cultures. Four cytokines (G-CSF, MCP-1, KC, and RANTES) were constitutively secreted by mouse BEC. When tri-cultured with murine pericytes and astrocytes, six other cytokines (IL-6, IL-13, TNF-alpha, eotaxin, MIP-1 alpha, and MIP-1 beta) were also secreted. EOO: Mouse brain endothelial cell monoculutures; EPA: Mouse brain endothelial cells tri-cultured with mouse pericytes and mouse astrocytes; n = 6/group.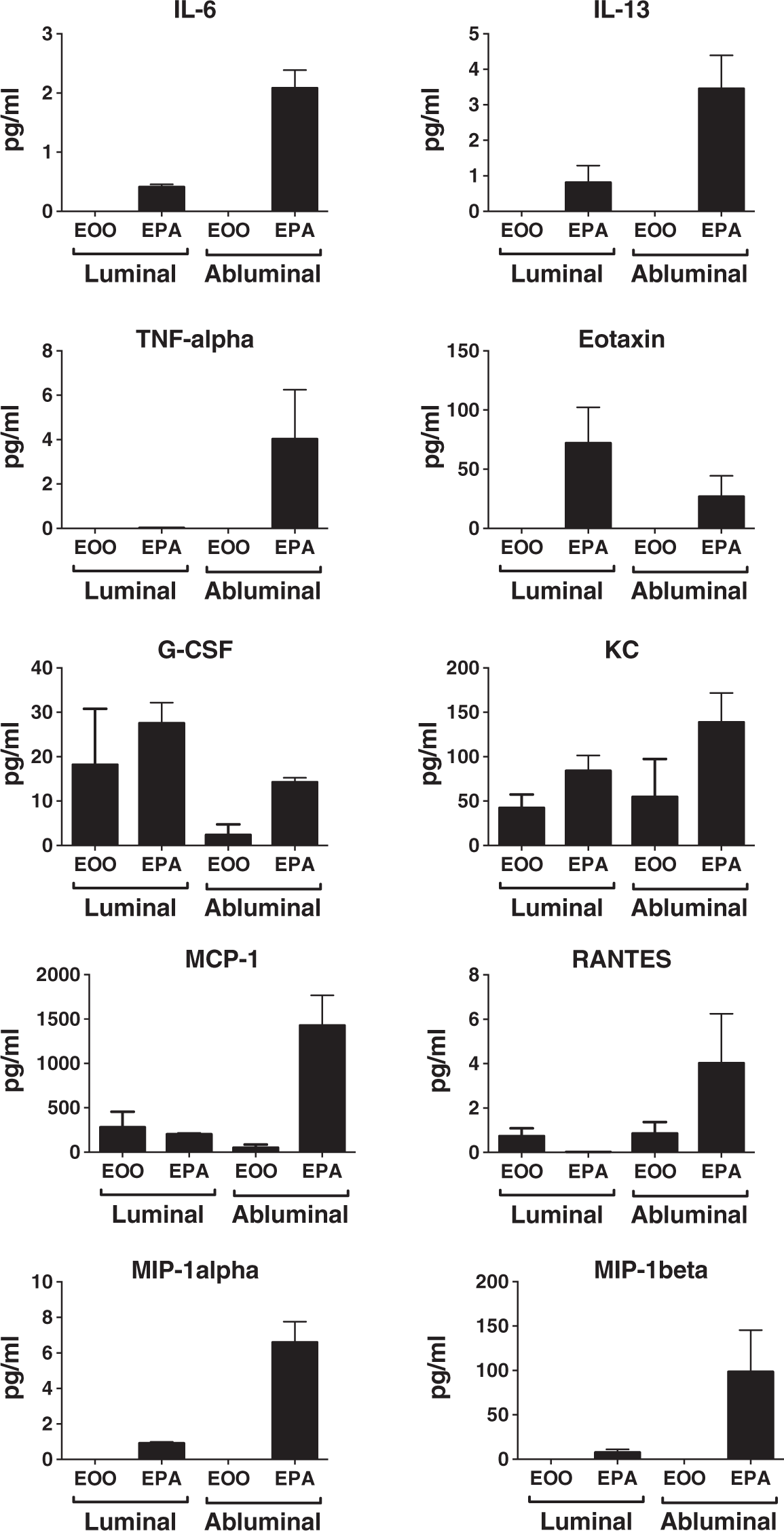
In another experiment, the same four cytokines and only these four cytokines (G-CSF, KC, MCP-1, and RANTES) were spontaneously secreted by monocultures of BEC (Figure 4). LPS placed in either the luminal or abluminal chamber of BEC monocultures greatly increased the secretion of these cytokines and in addition stimulated the release of eight other cytokines: IL-1 alpha, IL-6, IL-12(p40), IL-13, GM-CSF, MIP-1 alpha, MIP-1 beta, and TNF-alpha. When tri-cultures that consisted of all murine cells were exposed to LPS, all the cytokines seen with tri-cultures without LPS plus all the cytokines seen with monocultures plus LPS were observed except for IL-1 alpha, but IL-1 beta and IL-12(p70) were also seen (Figure 6).
Effects of LPS on cytokine secretion patterns in mouse brain endothelial cells in monocultures or tri-cultured with murine pericytes/astrocytes. Patterns for 26 cytokines are shown (Figure 4(a) to (z)). LPS was given either into the luminal or the abluminal chambers and cytokines measured in both the luminal (L) and abluminal (A) chambers. IL-1 alpha was detected in monocultures of BECs, but not in tri-cultures, whereas IL-1 beta, IL-12(p70), and eotaxin were detected in tri-cultures, but not in monocultures of BECs (n = 3–4/group. Newman–Keuls done for cytokines with ANOVA having p < 0.05. single letter (e.g. “a”) p < 0.05; double letter (e.g. “aa”) p < 0.01; triple letter (e.g. “aaa”) p < 0.001); a: luminal LPS increases luminal secretion; b: luminal LPS increases abluminal secretion; c: luminal LPS increases luminal secretion more than abluminal secretion; d: abluminal LPS increases luminal secretion; e: abluminal LPS increases abluminal secretion; f: abluminal LPS results in differences between luminal and abluminal levels; g: luminal secretion is greater with luminal LPS than with abluminal LPS; h: abluminal secretion is greater with abluminal LPS than with luminal LPS. EOO: Mouse brain endothelial cell monocultures; EPA: Mouse brain endothelial cell tri-cultured with mouse pericytes and mouse astrocytes; L-luminal chamber; A-abluminal chamber.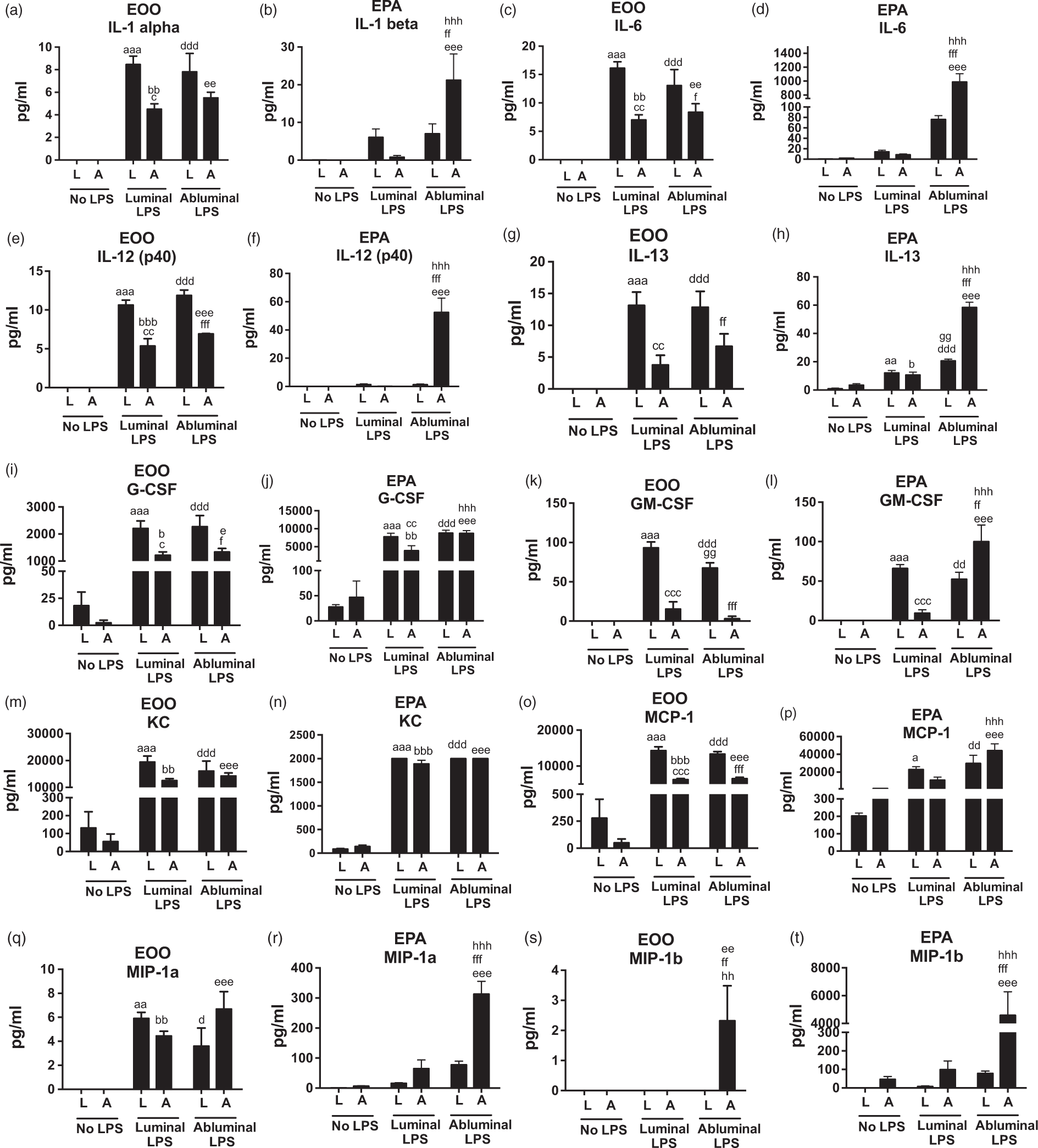
Of the 11 cytokines secreted by both LPS-stimulated monolayers and LPS-stimulated tri-cultures (G-CSF, KC, MCP-1, RANTES, IL-6, IL-13, MIP-1 alpha, MIP-1 beta, TNF, IL-12(P40), GM-CSF), the levels secreted by the LPS-stimulated tri-cultures tended to be higher than those in the LPS-stimulated BEC monocultures, in some cases 10–100 fold higher. IL-1 alpha, KC, and GM-CSF were exceptions. IL-1 alpha was only seen in LPS-stimulated monocultures of LPS, but not in LPS-stimulated or unstimulated tri-cultures. Secretion of KC was about 10 times lower in LPS-stimulated tri-cultures than in LPS-stimulated monocultures. GM-CSF expression was induced in LPS-stimulated monocultures of BEC but was not further enhanced in LPS-stimulated tri-cultures.
Previous results have suggested both that cytokine secretion differs from the luminal vs abluminal side of BECs and that the response to luminal vs. abluminal LPS may vary. 11 Therefore, we measured cytokine levels in the luminal and abluminal chambers after placing LPS in either the luminal or abluminal chamber. The statistical comparisons for each cytokine in Figure 4 showed eight secretion patterns:
Luminal LPS
Abluminal LPS
Increases luminal secretion (indicated statistically in Figure 4 as “d”). Increases abluminal secretion (indicated statistically in Figure 4 as “e”). Produces secretion levels that differ between luminal and abluminal chambers (indicated statistically in Figure 4 as “f”). Luminal secretion is greater with luminal LPS than with abluminal LPS (indicated statistically in Figure 4 as “g”). Abluminal secretion is greater with abluminal LPS than with luminal LPS (indicated statistically in Figure 4 with “h”).
With these eight patterns, cytokine secretion could be profiled. In the monocultures, for example, IL-6, IL-12(p40), G-CSF, and MCP-1 all showed the same pattern: Luminal LPS increased their luminal and abluminal levels with luminal levels being higher; abluminal LPS increased their luminal and abluminal levels, again with luminal levels being higher; LPS produced the same pattern regardless of whether it was given into the luminal or abluminal chamber.
In monocultures, KC, MIP-1 alpha, and RANTES produced the same pattern: Luminal LPS increased luminal and abluminal levels, but there was no difference between luminal and abluminal levels; abluminal LPS increased luminal and abluminal levels, but again no difference between luminal and abluminal levels; LPS produced the same pattern regardless of whether it was placed in the luminal vs. abluminal chamber.
The tri-cultures produced very different patterns from the monocultures in their responses to luminal and abluminal LPS. The most common pattern was exhibited by IL-1beta, IL-6, IL-12(p40), IL-12(p70), MIP-1alpha, and MIP-1beta. For these six cytokines, luminal LPS did not produce any statistically significant effect; abluminal LPS produced a statistically significant increase in abluminal, but not luminal levels; abluminal secretions were higher with abluminal than with luminal LPS. In the cases of IL-6, MIP-1alpha, and MIP-1beta, the lack of an effect of luminal LPS is surprising because the luminal LPS produced arithmetic increases that were equal to or greater than those seen in the BEC monocultures. It may be that the very large increases that occurred with abluminal LPS statistically overwhelmed the luminal LPS changes, causing them to be underpowered. Alternatively, factors produced by pericytes/astrocytes could be suppressing BEC cytokine production when induced by luminal LPS.
A paradoxic pattern for pattern “f” occurred between the monocultures and tri-cultures. In those cases when abluminal and luminal levels of a cytokine differed after abluminal LPS, the abluminal levels were always higher in the tri-cultures, but always lower in the monocultures of BECs.
Human pericytes/astrocytes and cytokine secretion
These patterns raised the question of the respective roles of endothelial cells vs. that of the pericytes and astrocytes in terms of cytokine secretions under both spontaneous and LPS-stimulated conditions. To investigate the respective roles of endothelial cells vs that of pericytes and astrocytes, we repeated the experiments with tri-cultures in which murine endothelial cells were cultured with human pericytes and human astrocytes. By then measuring cytokine levels with ELISAs that were specific for human or for mouse cytokines, we could determine the source of cytokine production (Figures 5 and 7). The human cytokine kit had 10 cytokines in common that had also been previously detected in the murine cell only experiments and therefore available for this sort of analysis: IL-1 beta, IL-6, IL-12(p70), IL-13, G-CSF, GM-CSF, MCP-1, MIP-1 beta, TNF-alpha, and IL-8, the human equivalent of murine KC. Of these, six were detected in either the mouse or human assays: GM-CSF, G-CSF, IL-8, MCP-1, IL-6, and TNF. G-CSF was only secreted by BECs, whereas IL-8 and TNF were only of pericyte/astrocyte origin. IL-6, MCP-1, and GM-CSF were of both endothelial and pericyte/astrocyte origin. All five cytokines secreted by pericytes/astrocytes were found in the luminal fluid and for MCP-1, GM-CSF, and TNF-alpha their luminal and abluminal concentrations were similar. This suggests that a robust brain-to-blood transfer mechanism may be present for these cytokines. Of the four cytokines secreted by endothelial cells, GM-CSF was only found in the luminal compartment, suggesting it is only secreted from the luminal surface. IL-6, MCP-1 and G-CSF of endothelial origin were found in both the luminal and abluminal chambers. For IL-6 and G-CSF, the two compartments had similar levels, but for BEC, MCP-1 was primarily luminal, suggesting preferential secretion from that surface.
Cell sources of cytokines in tri-cultures. By culturing mouse brain endothelial cells with human astrocytes/pericytes and then using species specific immunoassays, we were able to identify which cytokines were from brain endothelial cells (detected in the murine immunoassay) or pericytes/astrocytes (detected in the human immunoassay). Six of a possible 10 cytokines so detected are shown here; additionally, there was a notable absence of KC. En-Lum: Brain endothelial secretion in luminal chamber; PA-Lum: Pericyte/astrocyte secretion in luminal chamber; En-Abl: Brain endothelial secretion in abluminal chamber; PA-Abl: Pericyte/astrocyte secretion in abluminal chamber; LPS-Lum: LPS added to the luminal chamber; LPS-Abl: LPS added to the abluminal chamber. Summarized from Figure 5. Origin and occurrence of six cytokines as determined in tri-cultures of mouse brain endothelial cells and human pericytes/astrocytes. Orange indicates secretion by brain endothelial cells, green indicates pericyte/astrocyte origin. Note several cytokines of pericyte/astrocyte origin appeared in the luminal chamber, indicating passage across the BEC monolayer barrier. Larger font indicates a higher level of cytokine.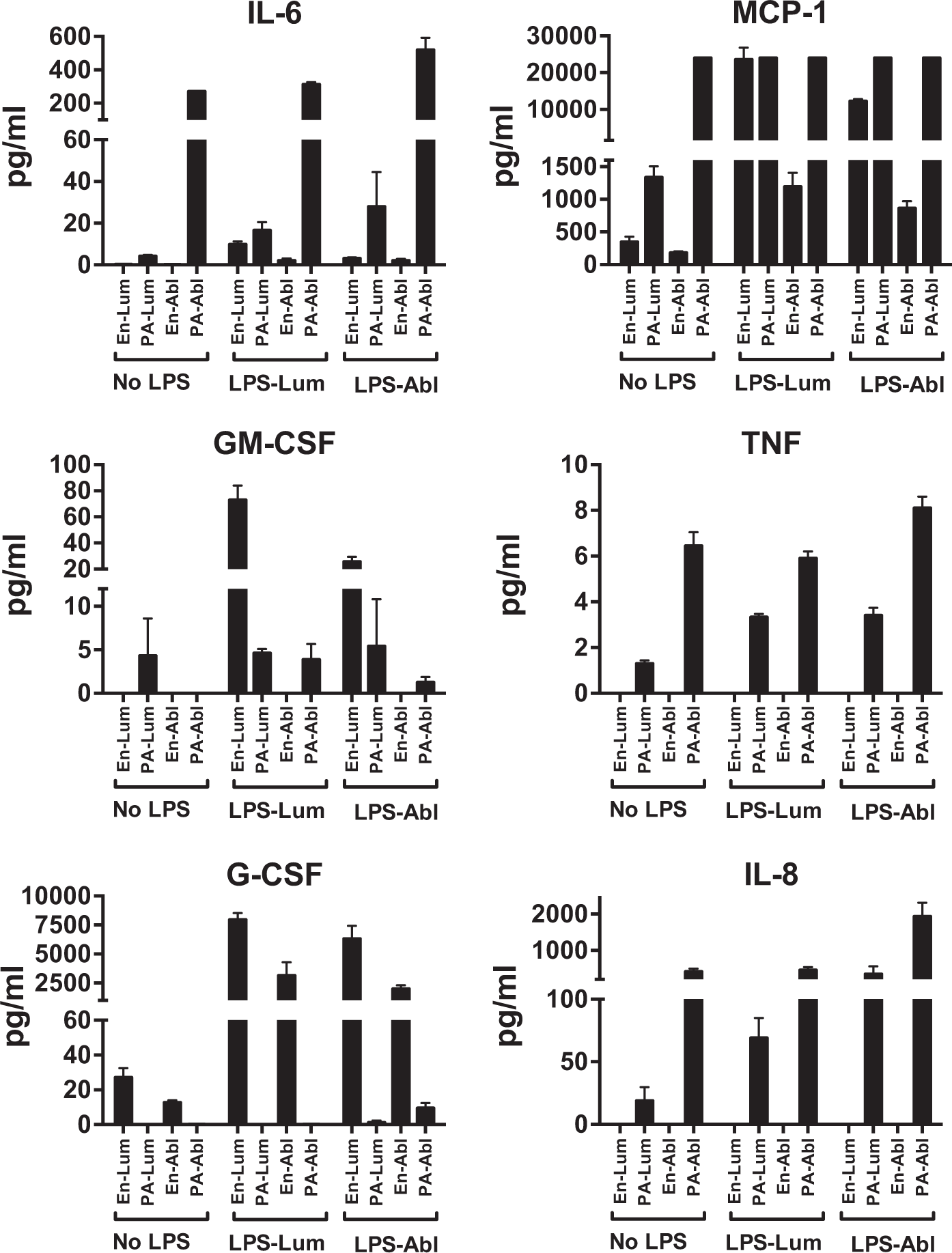
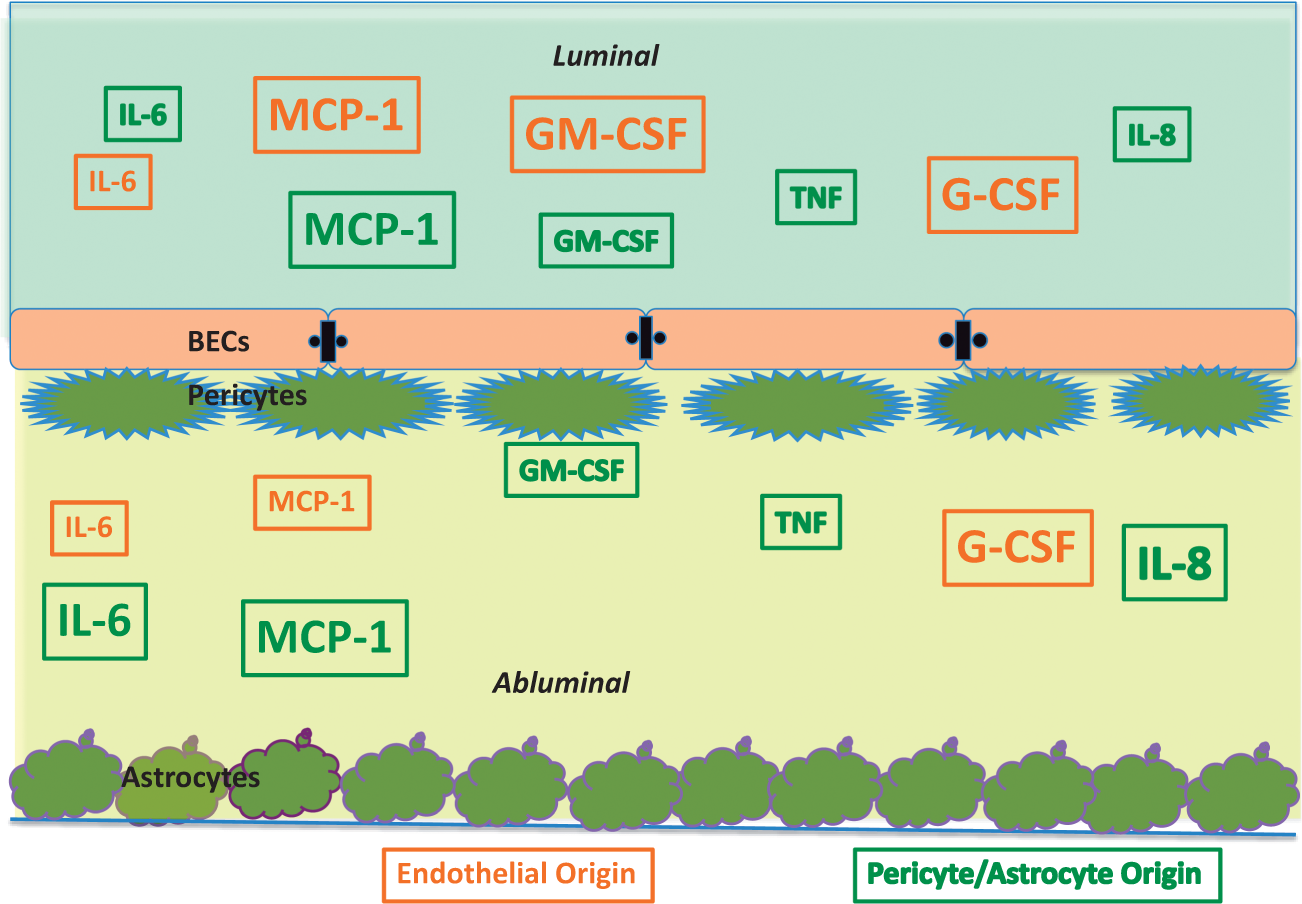
These culture results clarify some of the findings of Figures 3 (monocultures of BECs) and 4 (mouse tri-cultures). The most robust response of IL-6 in mouse tri-cultures was abluminal secretion after abluminal LPS. The results in Figure 5 show that IL-6 is secreted by BECs, but most of its release after either abluminal or luminal LPS is from pericytes/astrocytes. MCP-1 was present in high amounts in BEC monocultures and responded vigorously to abluminal and luminal LPS. In mouse tri-cultures, the most robust response was in abluminal levels after abluminal LPS. We found that MCP-1 was of BEC and pericyte/astrocyte origin and both categories of cells responded to LPS. For GM-CSF, the primary difference was that mouse tri-cultures had higher abluminal levels after abluminal LPS than BEC monocultures; otherwise, LPS had little effect on GM-CSF levels in either mouse monocultures or mouse tri-cultures. In the mouse + human cultures (Figure 5), GM-CSF responses resembled BEC monocultures more than mouse tri-cultures in that the abluminal increase in levels in response to abluminal LPS did not occur. GM-CSF in the absence of LPS was primarily of pericyte/astrocyte origin, but these cells did not increase secretion with LPS; rather, the large increases in GM-CSF were due to BEC secretion. TNF levels were barely detectable in BEC monocultures even with LPS. Even in mouse tri-cultures, only abluminal levels after abluminal LPS were in the reliably detectable range. The mouse + human studies only found TNF of pericyte/astrocyte origin. G-CSF levels in BEC monocultures favored secretion into the luminal surface, but in mouse tri-cultures, not only were levels higher overall, but abluminal LPS resulted in abluminal levels that were equal to luminal levels. This would be consistent with pericyte/astrocyte production predominating in this condition, but the mouse + human cultures showed that G-CSF was essentially of BEC origin; even with abluminal LPS, very little of the abluminal G-CSF (less than 1%) was of pericyte/astrocyte origin. KC levels in BEC monocultures were increased by LPS (either luminal or abluminal) about 10 fold greater than in mouse tri-cultures. KC is only of mouse origin, whereas IL-8 is its human equivalent, so this indicates that pericytes/astrocytes are inhibiting BEC secretion of KC. In the mouse + human studies, KC was totally absent, but IL-8 was present. This suggests a scenario that although BECs are capable of both constitutive and inducible release of KC/IL-8, pericyte/astrocytes suppress BEC secretions and themselves become the major source of KC/IL-8.
Discussion
These studies were designed to assess the interactions of pericytes and astrocytes with BECs. We mainly focused on cytokine and chemokine secretions as these secretions, as discussed in the introduction, have previously been shown to play important roles in autocrine and paracrine communications within the NVU. Advantages of the current study were our use of multi-analyte technology so that we could determine whether there were generalized patterns for many cytokines, assessment of both luminal and abluminal secretions so that we could assess polarized secretion, our comparison of varying conditions (monocultures vs tri-cultures, constitutive vs stimulated, luminal vs. abluminal LPS) so as to assess the influences of cross talk, activation, and polarized stimulation, and use of species specific assays so that we could determine whether the cellular source of a cytokine was BECs or pericytes/astrocytes.
Cross-talk was evident from the effects of co- and tri-cultures on induction of barrier functions in monolayers of mouse BECs. As previously noted, the addition of other cells of the NVU appeared to affect translocation of the tight junction proteins out of cytoplasm and to the inter-endothelial surface; 24 here, this was especially true for ZO-1 and claudin-5. We also found that permeability as measured by TEER, sucrose, or albumin was differentially affected by co- and tri-culturing. This probably reflects the different aspects of BBB permeability that these measures inventory. A decrease in TEER depends on patent pathways between the luminal and abluminal chambers. 25 As such, TEER measures primarily paracellular permeability. Albumin can also cross via the paracellular pathway when that pathway is robustly opened;26,27 otherwise, an increase in albumin leakage can reflect the re-introduction of transcytotic and transcellular pathways. 28 Sucrose, as a small water soluble molecule, can cross via openings in the paracellular pathway too small to allow albumin to cross, transcytotic pathways, and to a very limited extent can cross by transcellular diffusion.29,30 We found that the leakage of these substances across the monolayers was differentially affected by the presence of pericytes and astrocytes, consistent the different mechanisms by which the BBB can become disrupted.
Only four cytokines were spontaneously secreted by monocultures of BEC’s and these were found in both the luminal and abluminal chambers: G-CSF, KC, MCP-1, and RANTES (Figures 3 and 6). This is consistent with previous studies showing that cytokines can be secreted independently from either the luminal or abluminal surface of BECs.11,14 The luminal and abluminal concentrations did not differ statistically for any of these four cytokines and in comparison to tri-cultures and LPS-stimulated studies, levels were low.
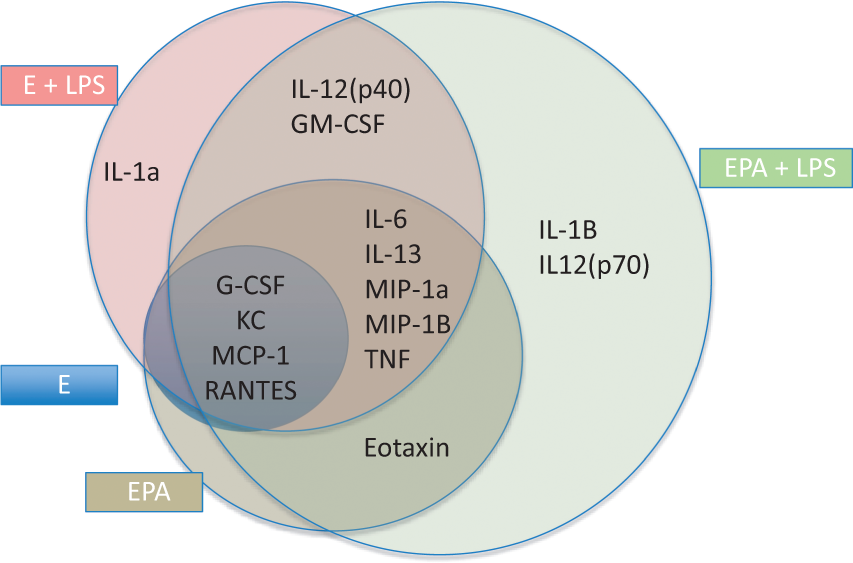
Tri-cultures and LPS-stimulated monocultures of BEC each secreted the same four cytokines seen in unstimulated monocultures of BEC, but at much higher levels, and secreted additional cytokines as well. We found five cytokines (IL-6, IL-13, MIP-1 alpha, MIP-1 beta, and TNF) that were additionally secreted by both LPS stimulation of monocultures and by unstimulated tri-cultures. This suggests that for these cytokines, pericyte/astrocyte cross-talk with BECs and LPS shares some fundamental pathways. In contrast, there were cytokines that were unique to LPS stimulation of monocultures of BECs (i.e. not seen with tri-cultures: IL12p40, GM-CSF, IL-1 alpha) and unique to tri-cultures (eotaxin), consistent with these conditions not being totally interchangeable or synonymous.It may be that TNF should be reclassified as a cytokine unique to the tri-cultures, considering that it was barely detectable in LPS-stimulated BEC monocultures. As discussed below, there is evidence that TNF is not of BEC origin.
Combining LPS stimulation with tri-cultures did not simply produce an additive effect, but produced a somewhat unique cytokine secretion pattern. Specifically, two additional cytokines were secreted (IL-1 beta and IL12(p70)) and IL-1 alpha was no longer secreted. This suggests that synergism does occur between the effects of pericytes/astrocytes and those of LPS. The loss of IL-1 alpha also suggests that not all influences are stimulatory, but that some were inhibitory. However, the vast majority of influences were found to be stimulatory; in no case, for example, was a cytokine level decreased by LPS treatment. The only other clear example of an inhibitory influence was that KC levels were lower in LPS-stimulated tri-cultures than in LPS-stimulated BEC monocultures.
Evidence for polarization was most strongly seen with LPS stimulation. In the monocultures, both luminal and abluminal LPS produced greater increases in the luminal than abluminal chambers, with only MIP-1 alpha, MIP-1 beta, and RANTES being exceptions. Two cytokines showed a differential response to where LPS was placed: GM-CSF was higher in the luminal chamber after luminal than after abluminal LPS and MIP-1 beta was higher in the abluminal chamber after abluminal than after luminal LPS.
The main polarization effect in tri-cultures occurred for IL-1 beta, IL-6, IL12(p40), IL12(p70), MIP-1 alpha, and MIP-1 beta in that abluminal LPS produced much higher abluminal levels than luminal LPS. Polarization allows the BBB to differentially respond to luminal and abluminal insults. Polarization also allows the BBB to receive input from one side (e.g. the brain side) and to release cytokine from the other (e.g. blood) side, as exemplified by the ability of luminal adiponectin to modulate the abluminal secretion of IL-6 in BEC monolayers. 31 Thus, polarization could be the basis for forming a neuroimmune axis capable of conveying information from one side of the BBB to the other side.
The different patterns obtained between monocultures and tri-cultures raised the question of how pericytes/astrocytes are altering secretion patterns. Are they also secreting cytokines or are they modifying cytokine secretion from the single cell source of the BEC? To address this question, we repeated the experiment in Figure 4 using mouse BECs but using human astrocytes and pericytes instead of mouse astrocytes and pericytes. By then assaying luminal and abluminal fluids using assays that are species-specific for human or for mouse cytokines, we could determine whether a cytokine was being secreted by BECs or by the pericytes and astrocytes. Finding that TEER was improved nearly as much by tri-culture with human pericytes/astrocytes as by mouse pericytes/astrocytes supported the assumption that mouse BEC cells were in cross-talk with human pericytes/astrocytes. Both types of LPS-treated tri-cultures also showed a very small preservation of TEER in comparison to the LPS-treated monoculture, possibly reflecting the protection glial cells can offer against LPS-mediated BBB injury. 32
Using this approach, we found a variety of patterns of cross-talk between BECs and pericytes/astrocytes (Figures 5 and 7). Figure 5 shows cytokines that had detectable levels in either the murine or human immunoassays. For several of these cytokines, knowing cellular origin aids in better understanding their functions as previously elucidated.
TNF-alpha is known to have many interactions with the BBB, including disrupting the BBB and being transported by it.33,34 Its transport across the BBB has been linked to its ability to mediate toxicity in animal models of chemobrain and Parkinson’s.35,36 Surprisingly, its secretion from in vitro in cultures of BECs is rarely reported. The results here show that TNF is not readily secreted by BECs, even when stimulated with LPS, but was here of pericyte/astrocyte origin.
IL-6 has been one of the most reported cytokines to be secreted in BEC monolayer cultures.15,37 IL-6 interactions with BECs and the BBB include its transport, increased immune cell adhesion, enhanced iron transport, and nitric oxide secretion.17,38–40 In studies that used only murine cells (Figure 4), IL-6 was not spontaneously secreted, whereas LPS stimulation produced much higher levels in the tri-cultures than in the monocultures. Not surprisingly then, Figure 5 shows that most of the IL-6 was derived from pericytes/astrocytes and less from endothelial cells. This was true even for luminal chamber IL-6 levels after abluminal LPS, suggesting that IL-6 crossed from the abluminal to the luminal chamber.
Figure 4 showed that tri-culture had little effect on the magnitude of GM-CSF secretion, except for increased abluminal levels after abluminal LPS. Figure 5 shows that BECs were the main source of GM-CSF with LPS stimulation. However, some luminal GM-CSF was derived from pericytes/astrocytes and all of the abluminal GM-CSF was from pericytes/astrocytes. This would suggest that GM-CSF is able to cross BECs in the brain-to-blood direction but that it is neither secreted from the abluminal surface of the BEC nor transported in the blood-to-brain direction. The luminal presence of IL-6 and GM-CSF is consistent with these two cytokines mediating the LPS-induced increase in transport of HIV-1 across BEC monolayers. 14 In that case, IL-6 and GM-CSF were proposed to act in autocrine fashion; the findings here are consistent with that hypothesis.
In Figure 4, G-CSF showed higher levels in both the luminal and abluminal chambers after LPS in the monoculture with even higher levels in the tri-cultures. Figure 5, however, shows that nearly all of the G-CSF was of BEC origin. Thus, pericytes/astrocytes increased G-CSF levels primarily through increasing BEC production rather than directly releasing it themselves. Evidence suggests that parenchymal brain cells are a source of G-CSF, 41 but the current results suggest that pericytes/astrocytes are not among those cell types. G-CSF is protective in several models of brain injury, including traumatic brain injury.42–44 G-CSF levels in blood have been proposed as a biomarker for acute traumatic brain injury and stroke.45,46 Its blood levels correlate with brain G-CSF levels and brain levels, in turn, correlate with the brain levels of a number of other cytokines. Thus, blood G-CSF levels are a reflection of post-traumatic neuroinflammation. These therapeutic and biomarker characteristics of G-CSF can be partially explained by its ability to cross the BBB, 47 but its vascular secretion could also play a role.
This approach of using species-specific assays was thus useful in determining the cellular source of cytokines in tri-culture, the movement of cytokines across the BECs monolayer, and the interactions between pericytes/astrocytes and BECs. One shortcoming of this approach is that a cell of one species may not respond to the cytokine of another. A classic example of this is that human TNF-alpha binds to the rodent p55 receptor but not to the rodent p75 receptor. 48 However, the improved TEER values seen in tri-cultures were only slightly less when the cells were mouse BECs plus human pericytes/astrocytes than when the cells were all mouse (Figure 2(d)). Thus, at least for TEER, this interspecies factor produced only a minor effect. However, decreased cross talk between cells because of species differences may be why we detected only 6 of the possible 10 cytokines with this approach.
Just as cells may not respond to stimulatory signals because of species differences, they may also not respond to inhibitory signals. This may explain why KC decreased when cultures were exposed to LPS and why it disappeared altogether when tri-cultured with human pericytes/astrocytes, although the human version of KC, IL-8, was present. This suggests that signals from BECs inhibit secretion of interferon-gamma and KC/IL-8 from pericytes/astrocytes.
Five cytokines of pericyte/astrocyte origin (IL-8, IL-6, GM-CSF, TNF, and MCP-1) were detected in the luminal chamber as well as the abluminal chamber. In most cases, levels in the abluminal and luminal chamber were similar. Such high levels suggest their appearance was not caused by leakage, but suggests that these cytokines may have been able to cross the BEC barrier in the abluminal to luminal direction. Only IL-2 and CCL1149,50 have been shown in vivo to have saturable transport systems in the brain-to-blood direction, but these results suggest that such systems may be much more common than originally thought. The ability of cytokines produced within the CNS to cross the BBB and enter the bloodstream provides a pathway by which the CNS can influence peripheral immune functions.
In conclusion, we examined the interactions of elements of the NVU in terms of BBB integrity and cytokine secretion. Overall, we confirmed that pericytes/astrocytes affect tight junction cytoarchitcture and BEC barrier functions as measured by TEER and sucrose permeability. We found that tri-cultures of BECs, pericytes, and astrocytes have different patterns of cytokine secretion both spontaneously and in response to LPS. In general, either tri-cultures or stimulation with LPS produced more and higher levels of cytokines. By culturing murine BECs with human pericytes/astrocytes and determining cytokine levels with species-specific immunoassays, we were able in several cases to study the cellular origin, movement, and cellular interactions of specific cytokines in the tri-cultures. We found different, individual patterns for the six cytokines so studied with these patterns providing evidence of cross-talk among the cells that comprise the NVU.
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by NIH RO1 AG046619 and Veterans Affairs.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Authors’ contributions
WAB contributed to experimental design, statistical analysis, interpretation, and writing.
AK and YM contributed to experimental execution, experimental design, interpretation, and writing.