
Abstract
Delayed remote ischemic postconditioning (DRIPost) has been shown to protect the rat brain from ischemic injury. However, extremely short therapeutic time windows hinder its translational use and the mechanism of action remains elusive. Because opening of the mitochondria KATP channel is crucial for cell apoptosis, we hypothesized that the neuroprotective effect of DRIPost may be associated with KATP channels. In the present study, the neuroprotective effects of DRIPost were investigated using adult male Sprague-Dawley rats. Rats were exposed to 90 minutes of middle cerebral artery occlusion followed by 72 hours of reperfusion. Delayed remote ischemic postconditioning was performed with three cycles of bilateral femoral artery occlusion/reperfusion for 5 minutes at 3 or 6 hours after reperfusion. Neurologic deficit scores and infarct volumes were assessed, and cellular apoptosis was monitored by terminal deoxynucleotidyl transferase nick-end labeling. Our results showed that DRIPost applied at 6 hours after reperfusion exerted neuroprotective effects. The KATP opener, diazoxide, protected rat brains from ischemic injury, while the KATP blocker, 5-hydroxydecanote, reversed the neuroprotective effects of DRIPost. These findings indicate that DRIPost reduces focal cerebral ischemic injury and that the neuroprotective effects of DRIPost may be achieved through opening of KATP channels.
Introduction
Stroke is the third leading cause of death in the United States. Approximately 795,000 people experience a stroke and> 143,579 people die each year. Of all strokes, 87% are ischemic (Lloyd-Jones et al, 2010). The current lack of clinical treatment for acute stroke necessitates the exploration of novel concepts that may eventually lead to clinical application. One of these concepts is ischemic postconditioning (IPost) (Pignataro et al, 2008a; Zhao et al, 2006), which refers to interference of blood flow by a series of brief, repetitive occlusion and release of cerebral blood vessels after reperfusion. Yang et al (2004) reported that reduction to infarct size by IPost was dependent on opening of KATP channels. However, the extremely short therapeutic time windows may hinder its clinical translation. This specific limitation may prevent its application to those patients in whom reperfusion cannot be immediately and accurately identified.
However, it has been reported that delayed postconditioning conducted 2 days after transient global ischemia attenuates hippocampal injury in gerbils (Burda et al, 2006). In addition, Ren et al (2008) reported that delayed postconditioning reduced ischemic injury after focal ischemia. Recently, a new phenomenon, known as remote IPost (RIPost), was found to induce ischemic tolerance not only within the same piece of tissue, but also in distant tissues as well as in distant organs (Tsubota et al, 2010). Loukogeorgakis et al (2007) revealed that transient limb ischemia induces RIPost in humans by a KATP channel-dependent mechanism. This result suggested that mitochondrial KATP channel activation has a key role in the development of a protective effect during IPost and RIPost. RIPost has greater potential for clinical application than classic IPost as it can be performed in a nonvital organ, avoiding the high risk of IPost in the vital organ. However, all reported IPost procedures have been applied either at the onset of reperfusion or during the ischemic phase. For patients with transient ischemic reperfusion brain injury, pharmacologic and physiologic interventions influence the effective time window of treatment. However, whether delayed remote IPost (DRIPost) attenuates brain injury after cerebral ischemia is unknown. In this study, we tested whether DRIPost, conducted by repetitive occlusion and release of the bilateral femoral arteries, reduces the infarcted area in focal ischemic rats. Accumulating evidence suggests that KATP channel activation is involved in the protective effect of IPost. Thus, we further tested whether DRIPost protects the brain from stroke and the potential protective mechanisms related with KATP channel activation.
Materials and methods
Animals
Male Sprague-Dawley rats weighing 290 to 310 g were provided by the Experimental Animal Centre of the Fourth Military Medical University and housed under diurnal lighting conditions (12 hours darkness/light). All experimental protocols and animal handling procedures were performed in accordance with the National Institutes of Health (NIH, USA) guidelines for the use of experimental animals and the experimental protocols were approved by the Institutional Animal Care and Use Committee of the Fourth Military Medical University. Diazoxide (DIAZ) and 5-hydroxydecanote (5-HD) were purchased from Sigma-Aldrich (St Louis, MO, USA).
Focal Cerebral Ischemia
Sprague-Dawley rats were allowed free access to food and water but were fasted 12 hours before surgery. All animals were anesthetized by intraperitoneal injection of pentobarbital sodium (50 mg/kg in normal saline). The transient middle cerebral artery (MCA) occlusion model was performed as previously described (Xiong et al, 2003). Briefly, the right common carotid artery (CCA) and the right external carotid artery were exposed through a ventral midline neck incision, and were ligated proximally. A 3-0 nylon monofilament suture (Ethicon Inc., Osaka, Japan) with a blunt tip made by burning on a flame was inserted through the arterectomy in the CCA just below the carotid bifurcation, positioned into the internal carotid artery and advanced ∼17 to 18 mm until a mild resistance was felt. Under these conditions, the origin of the MCAs was occluded. Reperfusion was accomplished by withdrawing the suture after 90 minutes of ischemia. The incision sites were infiltrated with 0.25% (v/v) bupivacaine hydrochloride for postoperative analgesia.
Regional cerebral blood flow was monitored using a flexible optical fiber probe attached to the skull over the ipsilateral parietal cortex at one point (1 mm posterior and 5 mm lateral to bregma) by laser Doppler flowmetry (PeriFlux system 5000; Perimed AB, Stockholm, Sweden). Rats in which ipsilateral blood flow was not reduced to <20% of the baseline after placement of the intraluminal filament and whose cerebral blood flow signal was not rapidly restored during reperfusion were excluded from subsequent experiments. Cranial temperature was maintained at 36.8°C to 37.5°C by surface heating and cooling during surgery. In a separate experiment, physiologic parameters (cranial temperature, arterial pH, Pa
Animal Recovery and Neurologic Evaluation
Rats were returned to their cages after the suture was withdrawn and were given free access to food and water. The neurologic behavior of rats was scored at 24, 48, and 72 hours after reperfusion by an investigator who was unaware of animal grouping. An 18-point scale of neurologic deficit scores (NDSs) was used for evaluation of neurologic behavior (Garcia et al, 1997). The scale was based on the following six tests: (1) spontaneous activity (0 to 3 points); (2) symmetry in the movement of four limbs (0 to 3 points); (3) forepaw outstretching (0 to 3 points); (4) climbing (1 to 3 points); (5) body proprioception (1 to 3 points); and (6) response to vibrissae touch (1 to 3 points). The six individual test scores were summed up at the end of the evaluation (minimum score, 3; maximum score, 18).
Infarct Volume Measurement
For measurements of infarct volume at 72 hours after cerebral artery reperfusion, rats were killed and the brains were rapidly removed and mildly frozen to keep the morphology intact during slicing. In brief, brains were cut into 2-mm-thick coronal sections in a brain matrix and stained with 2% (w/v) 2,3,5-triphenyltetrazolium chloride (Sigma-Aldrich) for 30 minutes at 37°C followed by overnight immersion in 4% (w/v) paraformaldehyde in phosphate buffer for after fixation. The infarct tissue area remained unstained (white), whereas normal tissue was stained red. Photographs were taken using a digital camera (SONY T9; Sony Corporation, Tokyo, Japan) The infarct area was demarcated and analyzed by Photoshop software (Adobe Photoshop 8.0, Adobe Systems Incorporated, San Jose, CA, USA). To compensate for the effect of brain edema, the corrected infarct volume was calculated as follows: percentage of corrected infarct volume = ((contralateral hemisphere area–(ipsilateral hemisphere—measured infarct area))/contralateral hemisphere area) × 100%. After brain extraction, animals observed to have experienced a subarachnoid hemorrhage were excluded from the study.
Experimental Protocol
Experiments were composed of two parts. The first part was designed to prove the neuroprotective effect of DRIPost and explore the optimal method of DRIPost. The second part was designed to investigate the role of KATP channels in the neuroprotective effect of DRIPost
Part 1: Neuroprotective Effect of Delayed Remote Ischemic Postconditioning Against Cerebral Ischemia—Reperfusion Injury
To explore the optimal parameters required to observe the protective effects of DRIPost, our pilot study tested 56 rats with 7 different serials of DRIPost performed at 3 or 6 hours after reperfusion (
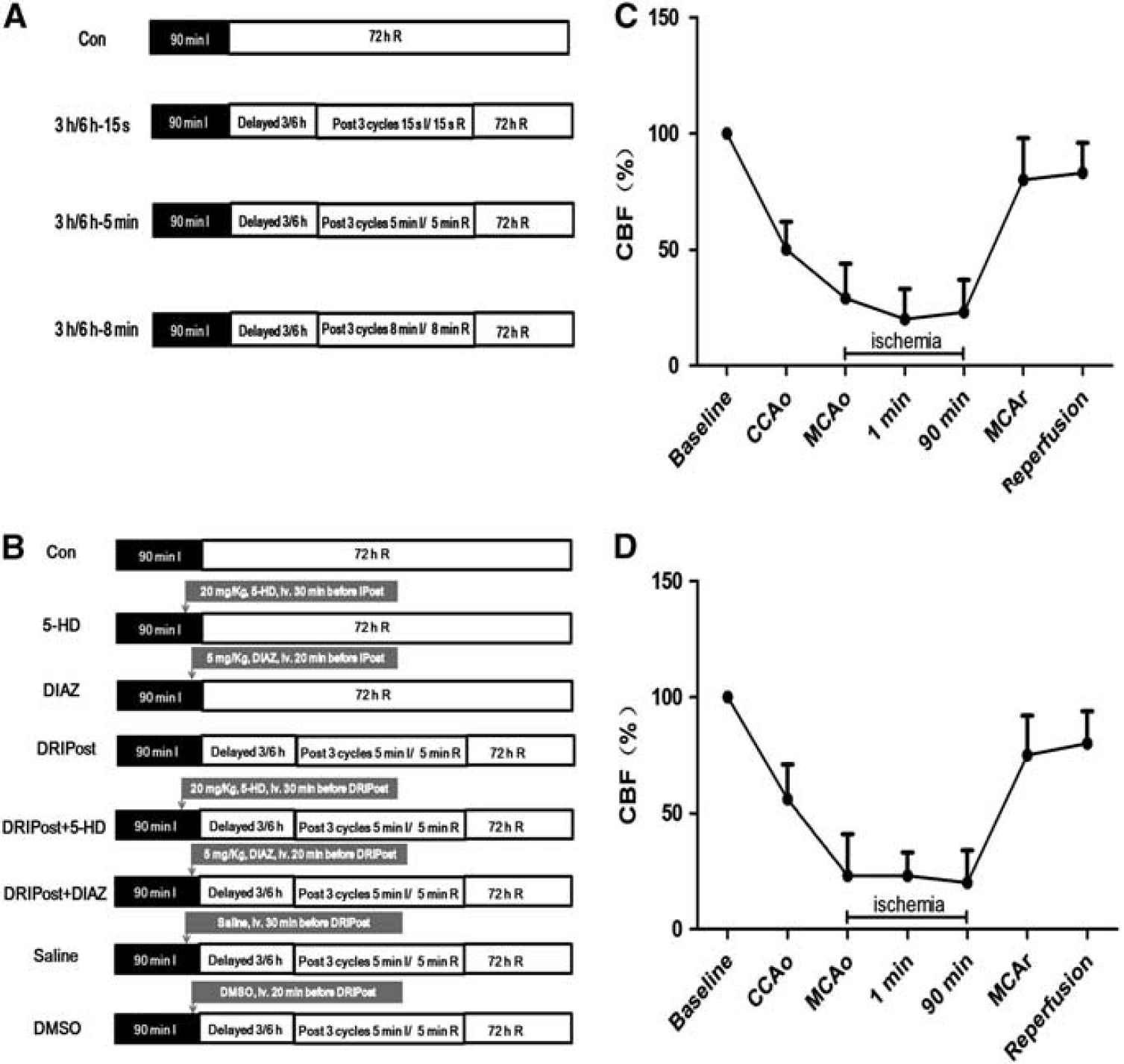
Schematic of the design and changes in cerebral blood flow (CBF) in animals for part 1 and part 2 of the experiment. All animals underwent 90 minutes of ischemia (dark bar) and 72 hours of reperfusion (open bar) (
Part 2: Effect of KATP Channel Blocking/Opening on Delayed Remote Ischemic Postconditioning-Induced Neuroprotection
In part 2, we further studied the role of KATP channels in the neuroprotective effect of DRIPost. Sixty-four adult male Sprague-Dawley rats were randomly divided into eight groups. The experimental protocols are shown in Figure 1B. Delayed remote ischemic postconditioning (5 minutes ischemia/5 minutes reperfusion in bilateral femoral arteries, three cycles) was induced at 6 hours after reperfusion in the postconditioning groups. This protocol showed a superior outcome in part 1 of the study. To block KATP channels, 20 mg/kg of 5-HD, which was dissolved in normal saline to a concentration of 12 mg/mL, was injected through the caudal vein 30 minutes before DRIPost. To open KATP channels, 5 mg/kg of DIAZ, which was dissolved in dimethyl sulfoxide (DMSO) to a concentration of 3 mg/mL, was administered through the caudal vein 20 minutes before DRIPost. To exclude the effect of administrated drugs on cerebral ischemia—reperfusion injury, two separate control groups were used, which were administered the same dosage of drugs without postconditioning. Vehicle groups (saline group and DMSO group) were also included to preclude any influence.
Terminal Deoxynucleotidyl Transferase Nick-End Labeling and Quantification of Apoptosis
Apoptosis was quantified using a commercially available fluorescent terminal deoxynucleotidyl transferase nick-end labeling (TUNEL) kit, in accordance with the manufacturer protocol (Roche Diagnostics Corporation, Indianapolis, IN, USA). The sections were mounted with 50% (v/v) glycerol for examination under a fluorescence microscope. The total number of TUNEL-positive neurons in the right hemisphere was counted in three different fields for each section in a blind manner by light microscopy at × 400 magnification (BX51; Olympus, Tokyo, Japan), and data from five animals at each stage were averaged.
Statistical Analysis
All data, excepting NDSs, are expressed as the mean ± s.d. Physiologic parameters were analyzed by repeated-measures analysis of variance. The NDSs were expressed as median (range). The NDSs among different groups were compared by Kruskal—Wallis test. When Kruskal—Wallis test showed significant difference, the Mann—Whitney
Results
Physiologic Parameters and Regional Cerebral Blood Flow
No statistical significance was noted among different time points for any of the physiologic parameters including cranial temperature, blood gas, and glucose concentrations (Table 1). Physiologic parameters remained in the normal range during the experimental period. Monitoring of regional cerebral blood flow ensured successful MCA occlusion (Figures 1C and 1D).
Physiological parameters, mean ± s.d. (
DRIPost, delayed remote ischemic postconditioning; Pa
Neurologic Deficit Scores
All rats survived until 72 hours after reperfusion. At 24, 48, and 72 hours after reperfusion, the NDS in the 3 h-5 min, 3 h-8 min, and 6 h-5 min groups was significantly higher than that in the Con group (
In part 2 of the experiment, at 24, 48, and 72 hours after reperfusion, the NDS in the DIAZ, DRIPost, DRIPost + DIAZ, and DRIPost + 5-HD groups was significantly higher than that of the Con group (
Infarct Volume
In experiment part 1, the infarct volume results are shown in Figure 2. All DRIPost protocols, except 3 h-15 s (60.48 ± 3.08%), significantly decreased infarct volumes at 72 hours after reperfusion compared with that of the Con group (63.88 ± 3.66%,
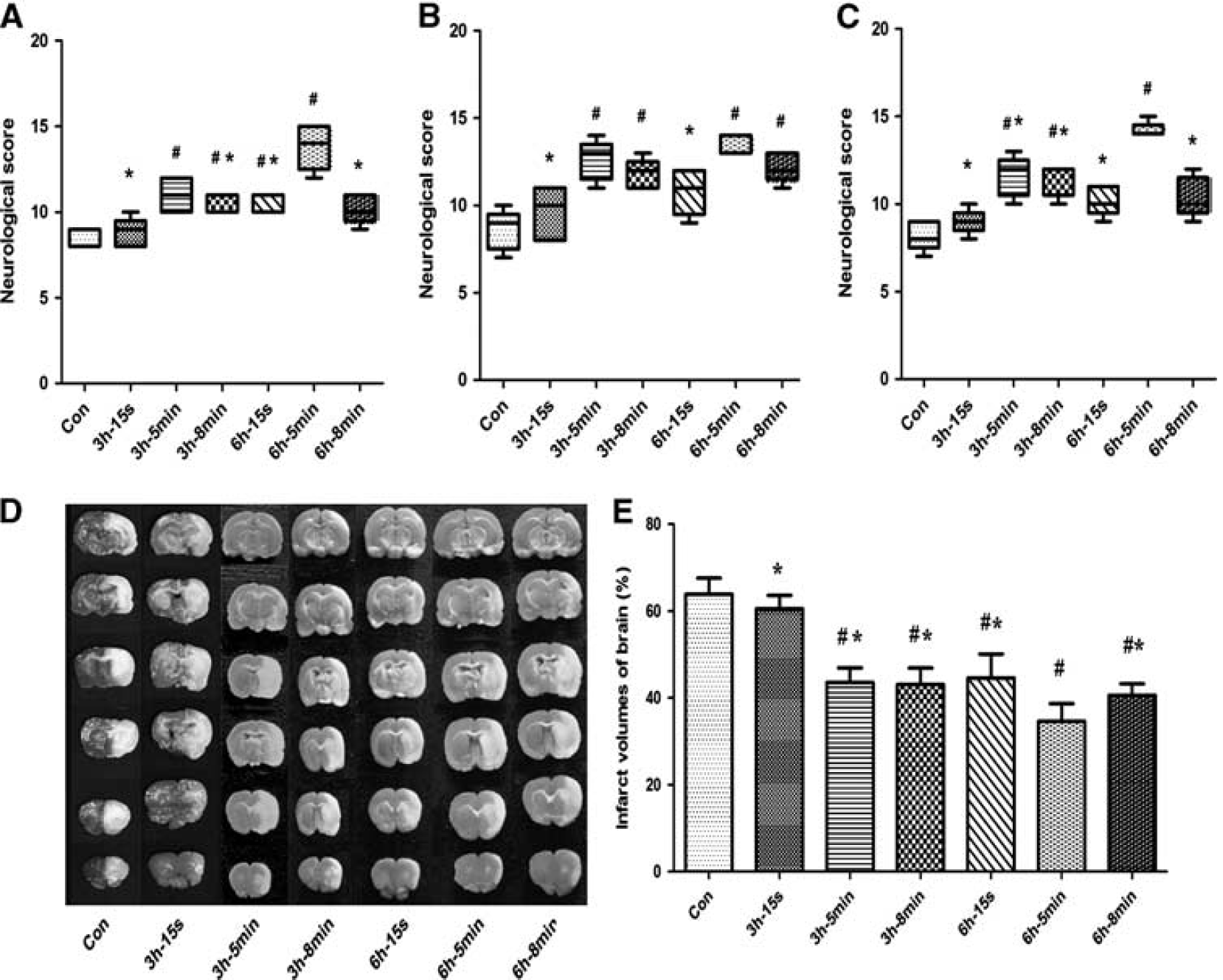
Neurologic scores and infarct volume after 90 minutes of transient middle cerebral artery occlusion in control (CON) and delayed remote limb ischemic postconditioning (DRIPost) groups in part 1 (
In experiment part 2, the infarct volume results are shown in Figure 3. Infarct volume in the DRIPost group (33.45 ± 2.78%) was significantly smaller than that of the Con group (65.58 ± 3.30%) at 72 hours after reperfusion (
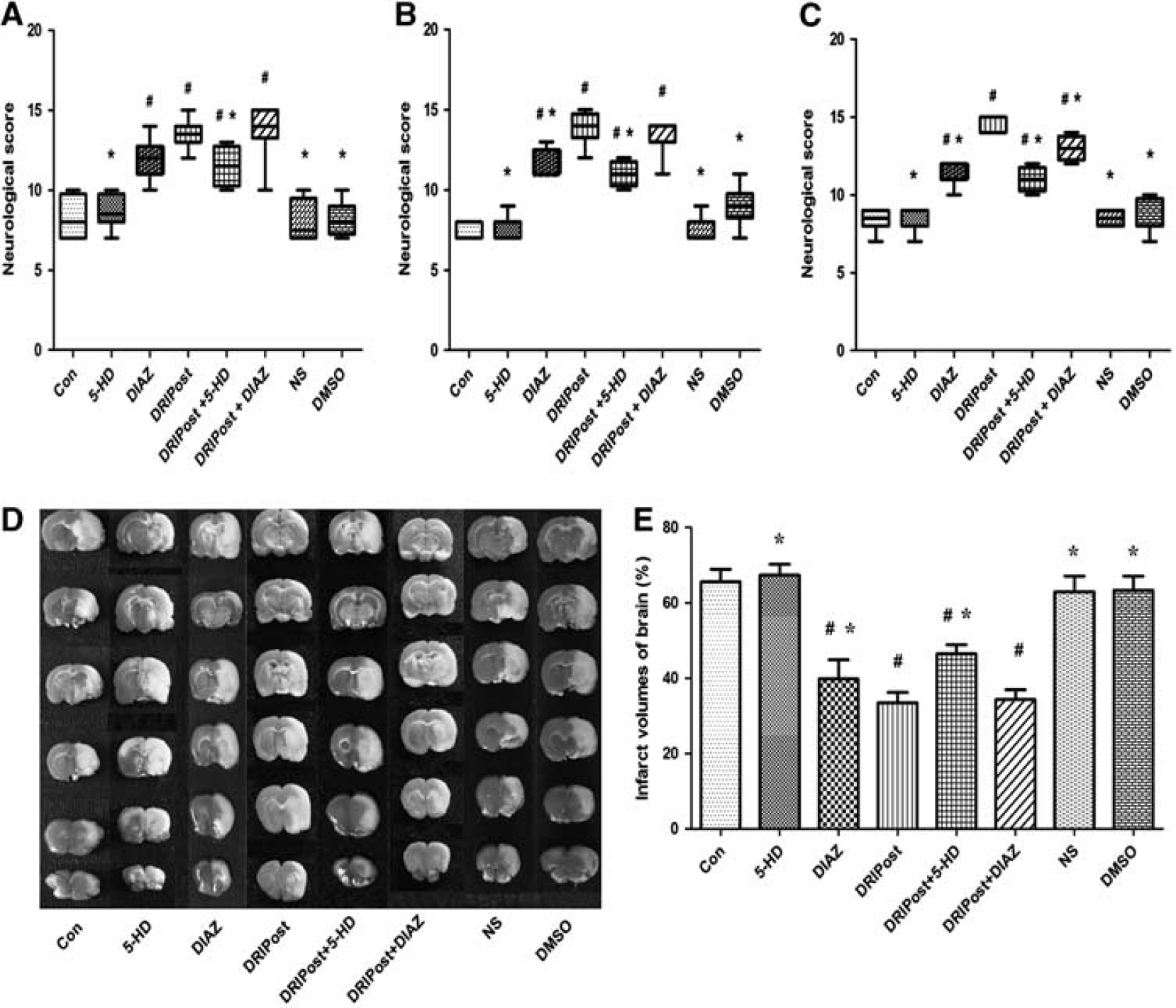
Neurologic scores and infarct volume after 90 minutes of transient middle cerebral artery occlusion in the control (CON), delayed remote limb ischemic postconditioning (DRIPost), 5-HD (KATP blocker, 20 mg/kg, intravenously), DIAZ (KATP opening, 5 mg/kg, intravenously), 0.9% (w/v) saline, dimethyl sulfoxide (DMSO) groups in part 2 (
Terminal Deoxynucleotidyl Transferase Nick-End Labeling
The number of TUNEL-positive neurons in the CA1 region significantly increased at 72 hours after ischemia—reperfusion injury in the Con group (Figure 4Aa). At the same observation time, the number of TUNEL-positive neurons in the CA1 region of the DRIPost group was significantly lower than that in the Con group (
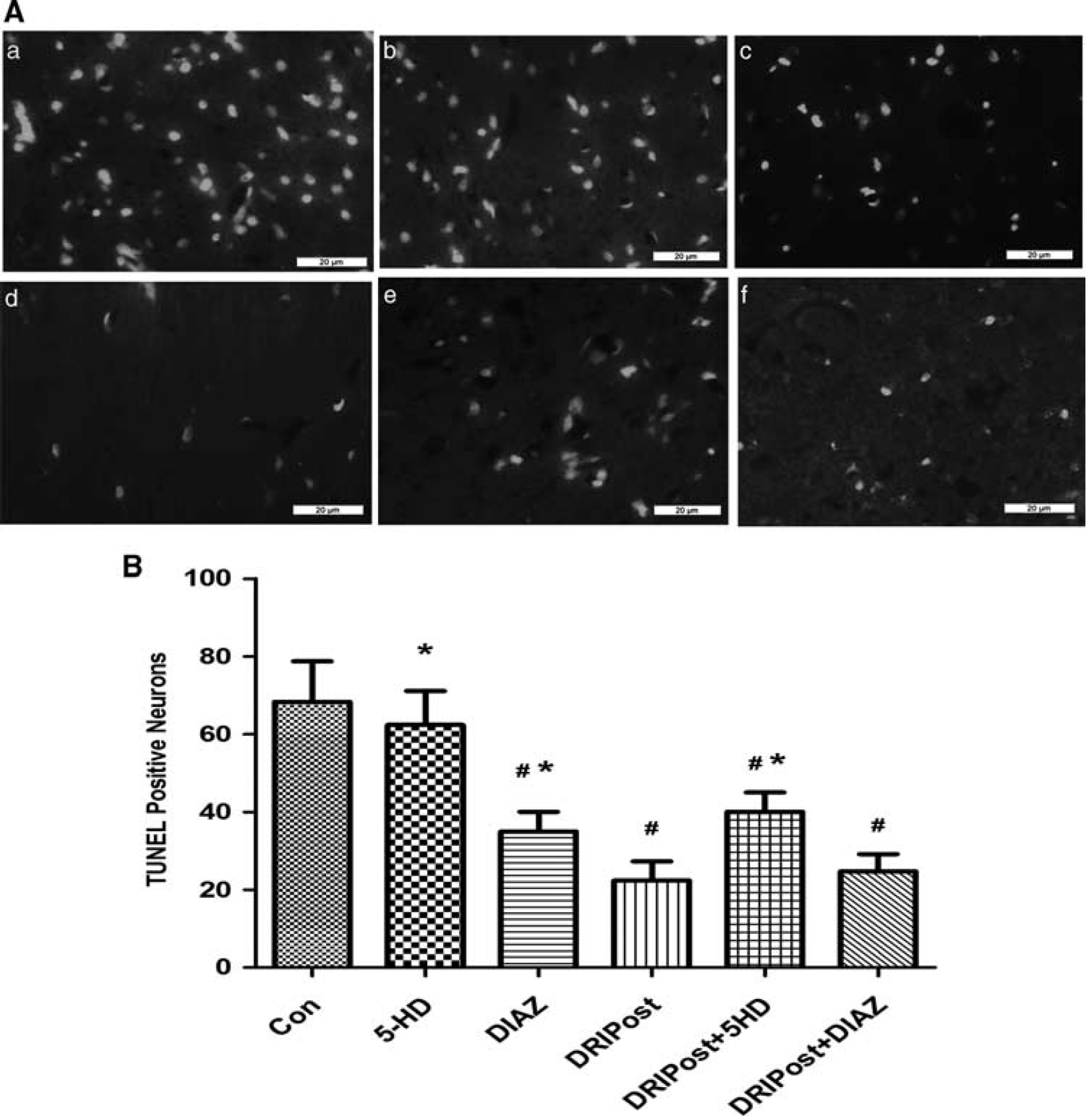
(
Discussion
This study showed that RIPost with three cycles of 5 minutes limb ischemia/5 minutes reperfusion, which was conducted even 6 hours after focal cerebral ischemia—reperfusion injury, could reduce infarct size and improve neurologic deficits. Ischemia—reperfusion-induced neuronal apoptosis was also markedly attenuated by DRIPost. Activation of KATP channels is critical for endothelial protection by DRIPost because the conditioning stimuli were ineffective when applied in the presence of the nonselective KATP channel blocker 5-HD. Taken together, the present study provides evidence that the neuroprotective effect of DRIPost is associated with the activation of KATP channel opening.
We found that the optimal procedure for DRIPost, performed a few hours later, differed from that of rapid RIPost. Currently, there are at least two types of IPost, conventional IPost and RIPost. The conventional IPost refers to a series of brief, repetitive mechanical occlusions/reperfusions at the onset of reperfusion after long-term ischemia (Zhao et al, 2003). As for brain injury, conventional IPost was first reported by Zhao et al (2006) in a rat cerebral ischemia model. This protective concept of IPost has been confirmed by a number of groups using
The underlying protective mechanisms of DRIPost are unknown. Nevertheless, our previous research from remote ischemic preconditioning (RIPC) against cerebral and spinal cord ischemia may shed light on our understanding of the protective mechanisms of DRIPost (Dong et al, 2010; Su et al, 2009). Emerging evidence from animal studies suggests that RIPC, IPost, and ischemic preconditioning share common signaling pathways, including triggers (adenosine receptor stimulation) (Kerendi et al, 2005; Steensrud et al, 2010), mediators (protein kinase C activation), and end effectors (opening of mitochondrial KATP channels, activation of prosurvival kinases, and inhibition of mitochondrial permeability transition pore opening; Hausenloy and Yellon, 2009; Meier et al, 2005; Yang et al, 2004). A popular theory has proposed that a substance or humoral factor, such as oxygen free radicals, is carried in the blood stream from the preconditioning organ or tissue to the organ where it manifests its protective effect. After humoral factors arrive to the remote organs, the end effectors (such as KATP channels) appear to be activated and have an important role in the induction of ischemic tolerance by RIPC. The protective effect of KATP has been showed in several tissues, including the intestines, kidneys, liver, and brain (Hai et al, 2005; Tawfik et al, 2009). Opening of KATP channels has been shown to be a prerequisite event for the induction of protection against ischemia—reperfusion injury by RIPC, IPost, and RIPost (Schmidt et al, 2007; Steensrud et al, 2010; Yang et al, 2004). Zhang et al (2010) used an
Several limitations of this study merit comments. First, only six different protocols were designed in this study. The results showed that DRIPost is still effective when applied at 6 hours after reperfusion. Whether the same results can be drawn at even later phases after injury or protocols where other parameters are altered (i.e., changes in the frequency of I/R cycle, application time or duration of occlusion and reperfusion of limb postconditioning) show more potent neuroprotection deserves further elucidation. Our results only exhibited a partial reversal of neuroprotection induced by DRIPost after 5-HD injection, suggesting that opening of KATP channels is not the only cause for the protective effect. In addition, differences could be due to different animal models (heart or brain ischemia—reperfusion injuries) used. Both timing and length of IPost have been confirmed to be important in determining the magnitude of its protective effect. Ren et al (2008) also found that the protective effect of limb RIPC depends on the number and duration of the limb ischemic stimulus. Previous studies have shown that endogenous activation of adenosine receptors, especially the A2A and A3A subtypes, is involved in IPost- and RIPost-mediated cardiac protection (Kerendi et al, 2005; Tsubota et al, 2010). Wang et al (2011) revealed that RIPost performed in one limb alleviated reperfusion injury after focal cerebral ischemia through reactive oxygen species-mediated inhibition of endogenous δ protein kinase C activation signaling cascade in an
In conclusion, DRIPost, even when applied at 6 hours after reperfusion, can induce a potent neuroprotective effect against focal cerebral ischemic reperfusion injury and inhibit apoptotic injury in the ischemic brain. The protective effects may partially be due to opening of KATP channels. Delayed remote ischemic postconditioning may be established as a potential practicable treatment for comparatively late-hour ischemic stroke patients who receive medical care hours after the onset of stroke.
Footnotes
The authors declare no conflict of interest.