
Abstract
The role of dopamine D1 receptors in prefrontal cortex function, including working memory, is well acknowledged. However, relatively little is known about their role in other cognitive or emotional functions. We measured both D1 and D2 receptors in the brain using positron emission tomography in healthy subjects, with the aim of elucidating how regional D1 and D2 receptors are differentially involved in cognitive and emotional functions beyond working memory. We found an inverted U-shaped relation between prefrontal D1 receptor availability and Wisconsin Card Sorting Test performance, indicating that too little or too much D1 receptor stimulation impairs working memory or set shifting. In addition, variability of D1 receptor availability in the amygdala and striatum was related to individual differences in emotional responses and decision-making processes, respectively. These observations suggest that the variability of available D1 receptors might be associated with individual differences in brain functions that require phasic dopamine release. An interdisciplinary approach combining molecular imaging of dopamine neurotransmission with cognitive neuroscience and clinical psychiatry will provide new perspectives for understanding the neurobiology of neuropsychiatric disorders such as schizophrenia, addiction and Parkinson's disease, as well as novel therapeutics for cognitive impairments observed in them.
Positron emission tomography imaging of D1 and D2 receptors and working memory
Because dopamine D1 receptors in the prefrontal cortex (PFC) are several times more abundant than D2 receptors (Hall et al, 1994), the relationship between D1 receptors and PFC functions has been widely investigated. Sawaguchi and Goldman-Rakic (1994) showed that local administration of D1 receptor antagonists into PFC induced impairment in working memory task in nonhuman primate. In human, Müller et al (1998) reported that systemic administration of a mixed D1/D2 agonist facilitated working memory while the selective D2 agonist had no effect, indicating that the dopaminergic modulation of working memory processes is mediated primarily via D1 receptors.
In contrast to D1 receptors, relatively less attention has been paid to the role of prefrontal D2 receptors in cognitive functions partly because their density in extrastriatal regions is very low (Suhara et al, 1999). It was reported that blockade of D2 receptors in PFC did not impair working memory in nonhuman primate (Sawaguchi and Goldman-Rakic, 1994), but some human studies reported that systemic administration of D2 agonist or antagonist modulated cognitive functions that are subserved by PFC (McDowell et al, 1998; Mehta et al, 1999). We measured both D1 and extrastriatal D2 receptor availabilities (binding potentials), indices proportional to receptor density, using [11C]SCH23390 and [11C]FLB457 positron emission tomography (PET), respectively, in healthy male subjects, and aimed to elucidate how regional D1 and D2 receptors are differentially involved in neurocognitive performance including frontal lobe functions. Receptor availability is defined as receptors that are available to be bound by the radiotracer. This means receptors that are available for stimulation by released endogenous dopamine.
A body of animal studies has indicated that stimulation of D1 receptors in PFC produces an inverted U-shaped dose–response curve, such that too little or too much D1 receptor stimulation impairs PFC functions (Cools and D’Esposito, 2011; Goldman-Rakic et al, 2000; Williams and Castner, 2006). Therefore, we conducted quadratic regression analysis to reveal the putative ‘U-shaped’ relation between D1 receptor availability in PFC and PFC function. Although standard linear regression analysis revealed a trend-level negative correlation between D1 receptor availability in PFC and total error of the Wisconsin Card Sorting Test (WCST), a test requiring working memory and set-shifting abilities, a quadratic regression model better predicted the relation (Takahashi et al, 2008). That is, we found a significant ‘U-shaped’ relation between D1 receptor availability in PFC and total error of WCST (because total error of WCST is a negative measure of frontal lobe function, the relation is not ‘inverted’; Figures 1A and 1B). However, neither linear nor quadratic relation was found between D2 receptor availability in PFC and any neuropsychological measures.
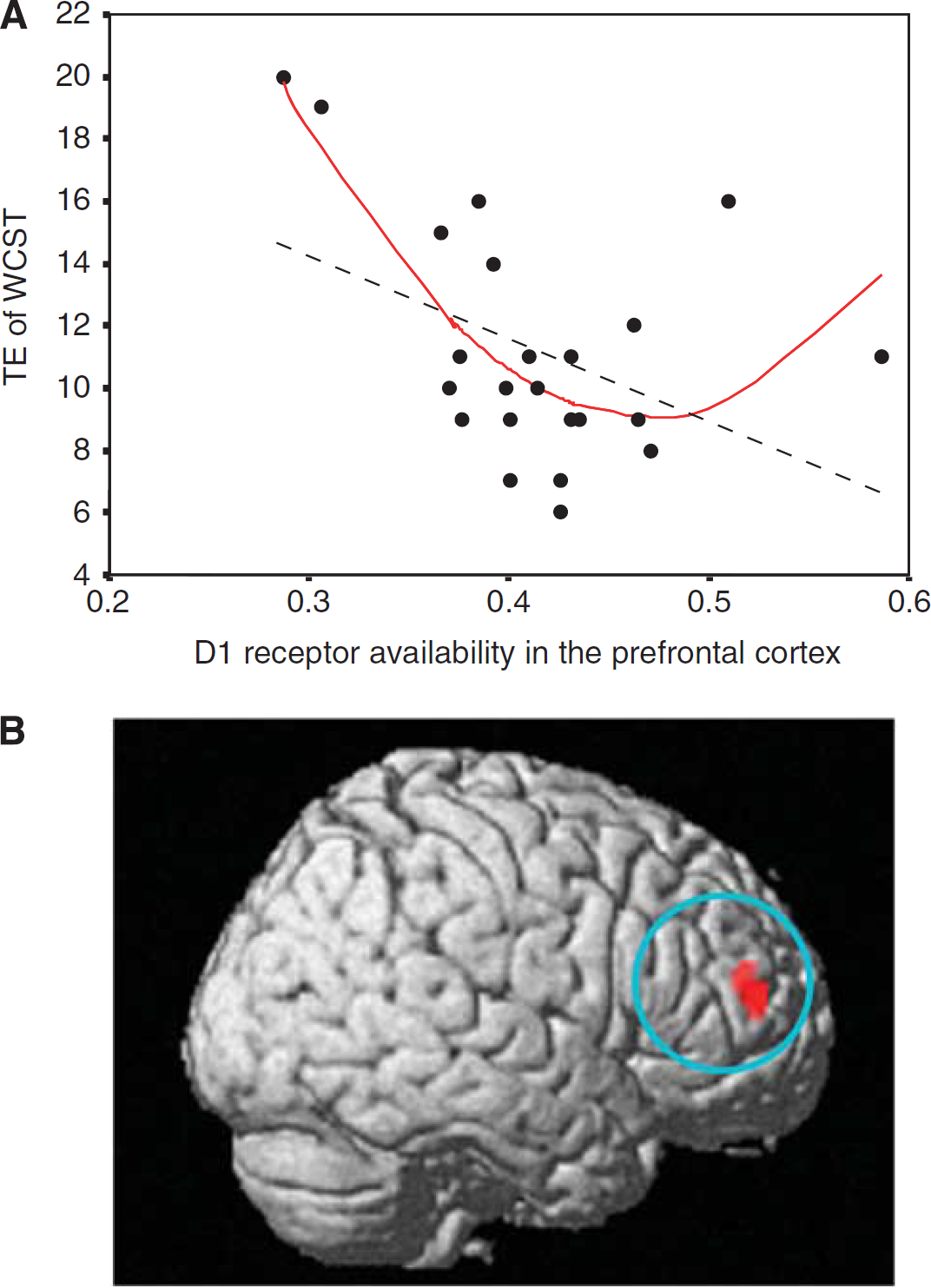
Quadratic (inverted U-shaped) relation between D1 receptor availability in prefrontal cortex (PFC) and performance of Wisconsin Card Sorting Test (WCST). (
Primal animal studies indicated that stimulation of D1 receptors in PFC produces an inverted U-shaped response in working memory, with the response being optimized within a narrow range of D1 receptor stimulation (Castner and Goldman-Rakic, 2004; Goldman-Rakic et al, 2000; Lidow et al, 2003; Seamans and Yang, 2004; Vijayraghavan et al, 2007). Subsequent human studies have investigated the effect of a functional polymorphism in the catechol O-methyltransferase gene, which has been shown to modulate the prefrontal dopamine level, on prefrontal function. Catechol O-methyltransferase gene contains a common polymorphism, a valine (Val)-to-methionine (Met) substitution at codon 158 (Val158Met). The Val allele is associated with higher activity, whereas the Met allele is associated with lower enzymatic activity. Consequently, Val carriers have a lower level of extracellular dopamine in PFC. A PET study using [11C]NNC112 has demonstrated that Val carriers show significantly higher cortical D1 receptor availability than Met carriers, and the authors suggested a mechanism in which a lower level of extracellular dopamine in PFC induces upregulation of D1 receptors in Val carriers (Slifstein et al, 2008). Val carriers show lower performance and increased (inefficient) PFC activation during completion of cognitive tasks related to PFC functions (WCST and N-back task) (Egan et al, 2001; Goldberg et al, 2003). It was reported that amphetamine challenge in Val carriers induced improvement in the performance of WCST and decreased (efficient) PFC activation during N-back task, whereas that in Met carriers caused deterioration in the performance of WCST and increased (inefficient) PFC activation, indicating that too little or too much dopamine signaling would impair PFC functions, although these studies could not identify the receptor subtype that has a central role in this effect (Mattay et al, 2003; Williams-Gray et al, 2007).
We first showed an inverted U-shaped relation between D1 receptors in PFC and executive function including working memory in normal healthy subjects (Takahashi et al, 2008). An inverted U-shaped response has been suggested based on cognitive and behavioral studies, but the exact physiological mechanism of this effect has not yet been fully understood. A recent monkey electrophysiology study has demonstrated a neuron-level mechanism that constitutes the inverted U-shaped response whereby too much or too little stimulation of prefrontal D1 receptors leads to working memory deficits. D1 receptor stimulation had a suppressive effect on the PFC neural activities involved in a spatial working memory task. Moderate D1 receptor stimulation spatially tunes PFC neurons that process target signals by preferentially suppressing nontarget (noisy) neural activities, whereas excessive D1 receptor stimulation induces nonselective suppression of PFC neural activities irrespective of whether the neural activities are task related or not (Vijayraghavan et al, 2007).
Animal studies have suggested that the inverted U-shaped principle of D1 receptor stimulation mediating working memory does not necessarily apply to other prefrontal functions (Floresco and Magyar, 2006). In fact, except for WCST, we did not find any association between D1 receptor availability and prefrontal functions less dependent on the working memory process (word fluency task by phonetic or semantic cues and problem-solving test; Takahashi et al, 2008).
Recently, McNab et al (2009) showed the quadratic relation between the improvement of working memory capacity by training and the change in D1 receptor availability induced by training, although greater reduction in D1 receptor availability was associated with greater improvements in working memory capacity within the measured range. However, a recent study showed that age-related reduction in D1 receptor availability in PFC was associated with age-related reduction in working memory performance and PFC activation during working memory load (Bäckman et al, 2011), indicating that other factors besides D1 receptor availability, such as cerebrovascular pathology, could influence the PFC functions and PFC activation during working memory load in older adults. Furthermore, although [11C]SCH23390 and [11C]NNC112 are selective radioligands for D1 receptors, they have some affinity for 5HT2A receptors. 5HT2A receptor density in the striatum is negligible compared with D1 receptor density, whereas 5HT2A receptor density is not negligible in the extrastriatal regions. Previous reports in the literature have indicated that their affinity for 5HT2A receptors relative to D1 receptors is negligible, and recent
In line with our previous study (Takahashi et al, 2007), we also found that D2 receptor availability in the hippocampus (HPC) was positively correlated not only with episodic memory ability but also with WCST performance (Takahashi et al, 2008). Patients with lesions in HPC sometimes show deficits in WCST (Corkin, 2001; Igarashi et al, 2002). These observations suggest that hippocampal D2 receptors could modulate PFC activity by the HPC–PFC pathway, which has a significant role in the cognitive process (Laroche et al, 2000; Thierry et al, 2000). Accumulating evidence has suggested the modulatory effects of dopamine on HPC–PFC interactions (Aalto et al, 2005; Goto and Grace, 2008; Seamans et al, 1998; Tseng et al, 2007). Conceivably, dopamine influences PFC neurons directly by prefrontal D1 receptors and indirectly by hippocampal D2 receptors via the HPC–PFC pathway. Supporting the importance of hippocampal D2 receptors in PFC functions, MacDonald et al (2009) reported that lower D2 receptor availability in HPC was associated with greater intraindividual variability in episodic memory and executive function, indicating that lower D2 receptor-mediated transmission in HPC leads to noisy neural information processing and results in unstable episodic memory and executive functions.
Müller et al (1998) reported that the systemic administration of the mixed D1/D2 agonist pergolide facilitated working memory while the selective D2 agonist bromocriptine had no effect. However, there is converging evidence from human and animal studies to suggest the involvement of D2 receptors in cognitive functions. It was reported that the systemic administration of the D2 agonist bromocriptine in human improved cognitive functions including working memory and executive functions (McDowell et al, 1998), and the administration of the D2 antagonist sulpiride impaired those functions (Mehta et al, 1999). In an animal study, mice lacking D2 receptors were reported to have a working memory deficit (Glickstein et al, 2002). These studies, however, did not reveal the regions most responsible for these effects. Moreover, although the involvement of D1 receptors in working memory is widely recognized, it was not clear whether D1 receptor stimulation alone or the combination of D1 and D2 receptor stimulation is most effective. Positron emission tomography findings including ours suggested that orchestration of prefrontal D1 receptors and hippocampal D2 receptors might be necessary for normal prefrontal functions (MacDonald et al, 2009; Takahashi et al, 2007, 2008).
Positron emission tomography imaging of D1 and D2 receptors and amygdala function
The amygdala has a central role in processing affective stimuli, and in particular, threatening stimuli in the brain (LeDoux, 2000). The amygdala receives a moderate innervation of dopaminergic fibers (Asan, 1998), and dopamine D1 and D2 receptors are moderately expressed in this region (Ito et al, 2008). Dopamine release in the amygdala is increased in response to stress (Inglis and Moghaddam, 1999). It has been shown in animal studies that dopamine potentiates the response of the amygdala by augmenting excitatory sensory input and attenuating inhibitory prefrontal input to the amygdala (Rosenkranz and Grace, 2002). A human functional magnetic resonance imaging (fMRI) study reported that dopaminergic drug therapy such as levodopa or dopamine agonists partially restored amygdala activation due to emotional task in Parkinson's disease patients who showed no significant amygdala activation during drug-off states (Tessitore et al, 2002). In addition, another fMRI study of healthy volunteers has demonstrated that amphetamine potentiated the response of the amygdala during an emotional task (Hariri et al, 2002). More recently, Kienast et al (2008) reported that dopamine storage capacity in human amygdala, measured with 6-[(18)F]fluoro-L-DOPA PET, was positively correlated with fMRI signal changes in the amygdala. However, contribution of dopamine D1 and D2 receptors to amygdala activation in response to affective stimuli is unknown in human. To investigate the relation between amygdala activation and dopamine receptor subtype, we conducted a multimodal
Although robust bilateral amygdala activations induced by fearful faces were identified in a group analysis, there was considerable individual difference in the degree of amygdala activation. Similarly, although moderate levels of D1 and D2 receptors in the amygdala were measured, notably high variances in both receptor availabilities were observed. Importantly, D1 receptor availability in the amygdala was not correlated with D2 receptor availability in the amygdala. Both voxelwise statistical parametric mapping analysis and regions of interest analysis revealed that blood oxygen level-dependent signals in the amygdala induced by fearful faces were positively correlated with D1 receptor availability, but not with D2 receptor availability, in the amygdala (Figures 2A and 2B; Takahashi et al, 2010b). That is, individuals with high D1 receptor density in the amygdala tend to show greater amygdala activation in response to fearful stimuli.
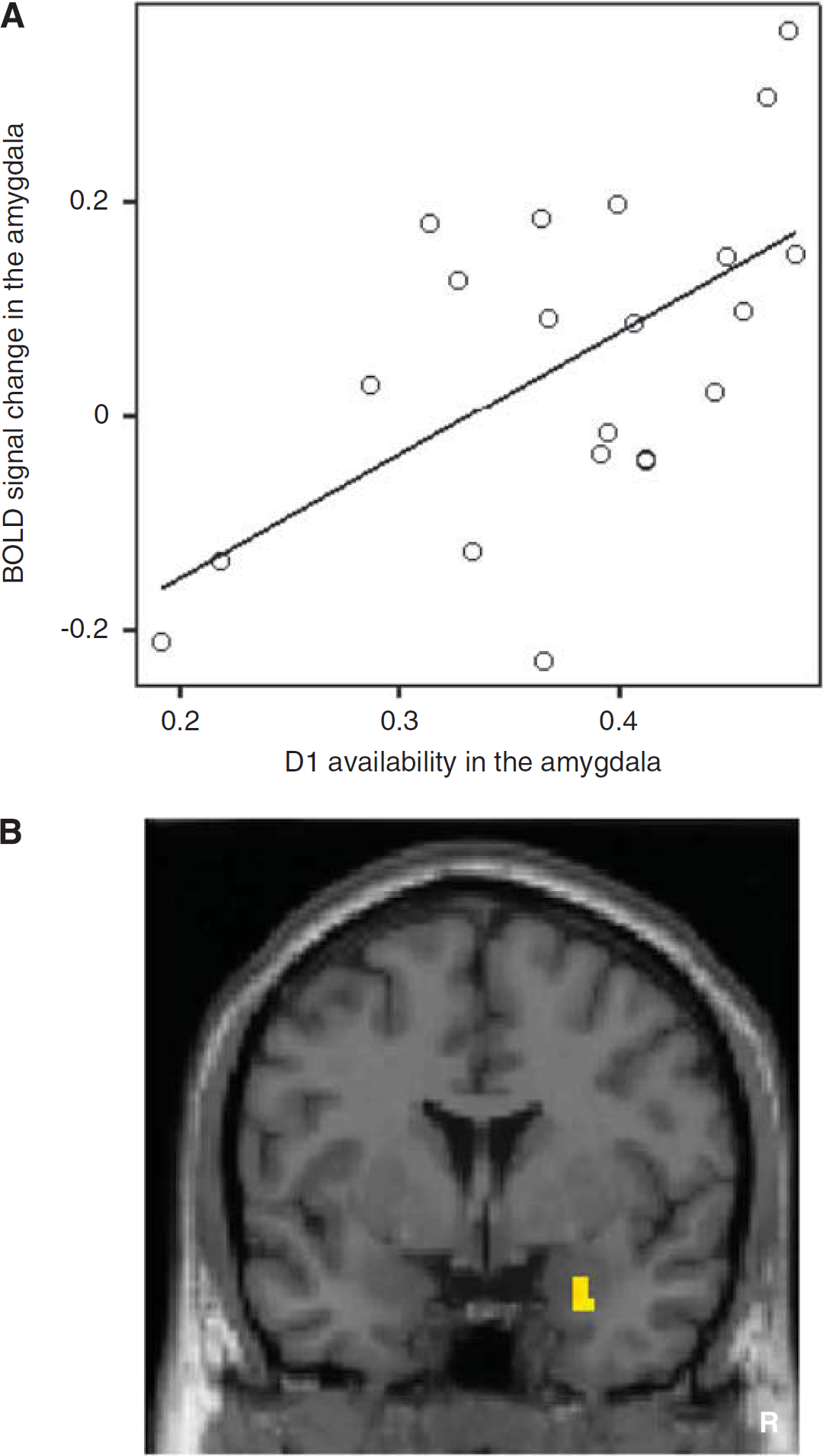
(
In rat studies, Rosenkranz and Grace (2002) showed that dopamine enhances the response of the amygdala by augmenting excitatory sensory input via dopamine D2 receptor stimulation and attenuating inhibitory prefrontal input to the amygdala through dopamine D1 receptor stimulation. More recently, several studies showed that both D1 and D2 receptor stimulations directly enhanced the excitability of amygdala projection neurons via postsynaptic mechanism (Kroner et al, 2005; Rosenkranz and Grace, 2002; Yamamoto et al, 2007). Amygdala projection neurons are under inhibitory control by GABAergic interneurons (Royer et al, 1999). Both projection neurons and interneurons in the amygdala express dopamine D1 and D2 receptors (Rosenkranz and Grace, 1999). Dopamine and D1 receptor agonist have been shown to augment interneuron excitability and increase the frequency of inhibitory postsynaptic current in amygdala projection neurons (Kroner et al, 2005). This is a counterintuitive result, considering the fact that dopamine disinhibits amygdala response
Using a multimodality
Positron emission tomography imaging of D1 and D2 receptors and decision making under risk
Decision making under risk has been studied in philosophy, psychology, and economics throughout the last century. Normative economic theories (e.g., expected utility theory) assume that individuals are rational decision makers and have purely self-regarding preferences. However, we sometimes make boundedly rational decisions (altruistic behavior, moral judgment, gamble, etc.), which are not accounted for by normative economic theories. Behavioral or experimental economics studies have shown a substantial body of field and empirical evidence that decision makers systematically depart from Camerer and Loewenstein (2004). One type of systematic departure is that subjective weights on probabilities appear to be nonlinear: people often overestimate low probabilities (e.g., playing lotteries) and underestimate high probabilities. A leading alternative to the expected utility theory is the prospect theory (Tversky and Kahneman, 1992). The central feature of the prospect theory is nonlinear probability weighting. Objective probabilities,
A synthesis of economics and neuroscience is called neuroeconomics. Neuroeconomics fMRI studies have demonstrated the neural basis for boundedly rational decision makings under risk, including some features of the prospect theory (De Martino et al, 2006; Tom et al, 2007). A deeper question is how modulatory neurotransmission is involved in the central process of these boundedly rational decision makings (Fox and Poldrack, 2009; Rangel et al, 2008; Trepel et al, 2005). Investigation of the relationship between the dopamine system and prospect theory seems promising, considering the fact that dopamine is linked to risk-seeking behavior (Leyton et al, 2002) and is involved in disrupted decision making observed in neuropsychiatric disorders such as drug/gambling addiction and Parkinson's disease (Steeves et al, 2009; Zack and Poulos, 2004). Based on the circumstantial findings, Trepel et al (2005) speculated in a thoughtful review that dopamine transmission in the striatum might be involved in shaping probability weighting. To test this speculation, 18 healthy male subjects were studied for D1 receptors with [11C]SCH23390 PET, and 18 other healthy male subjects were studied for striatal D2 receptors with [11C]raclopride PET (Takahashi et al, 2010a). To estimate decision weight, certainty equivalents were determined outside the PET scanner, based on the staircase procedure suggested by Tversky and Kahneman (1992). A gamble's certainty equivalent is the amount of sure payoff at which a player is indifferent between the sure payoff and the gamble. Participants were presented with options between a gamble and a sure payoff on a computer monitor. Gambles were presented that had an objective probability
In the first group, with D1 receptors investigated, mean (s.d.)
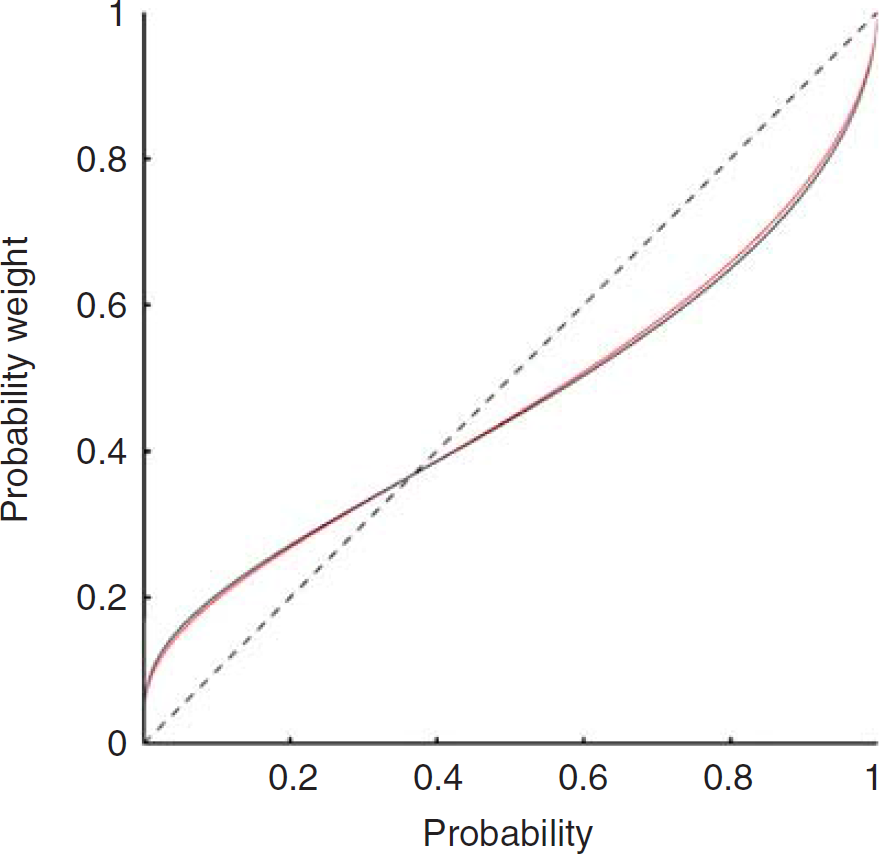
Average fitted probability-weighting function. Red line represents the first group with D1 receptors investigated, and black line the second group with striatal D2 receptors investigated.
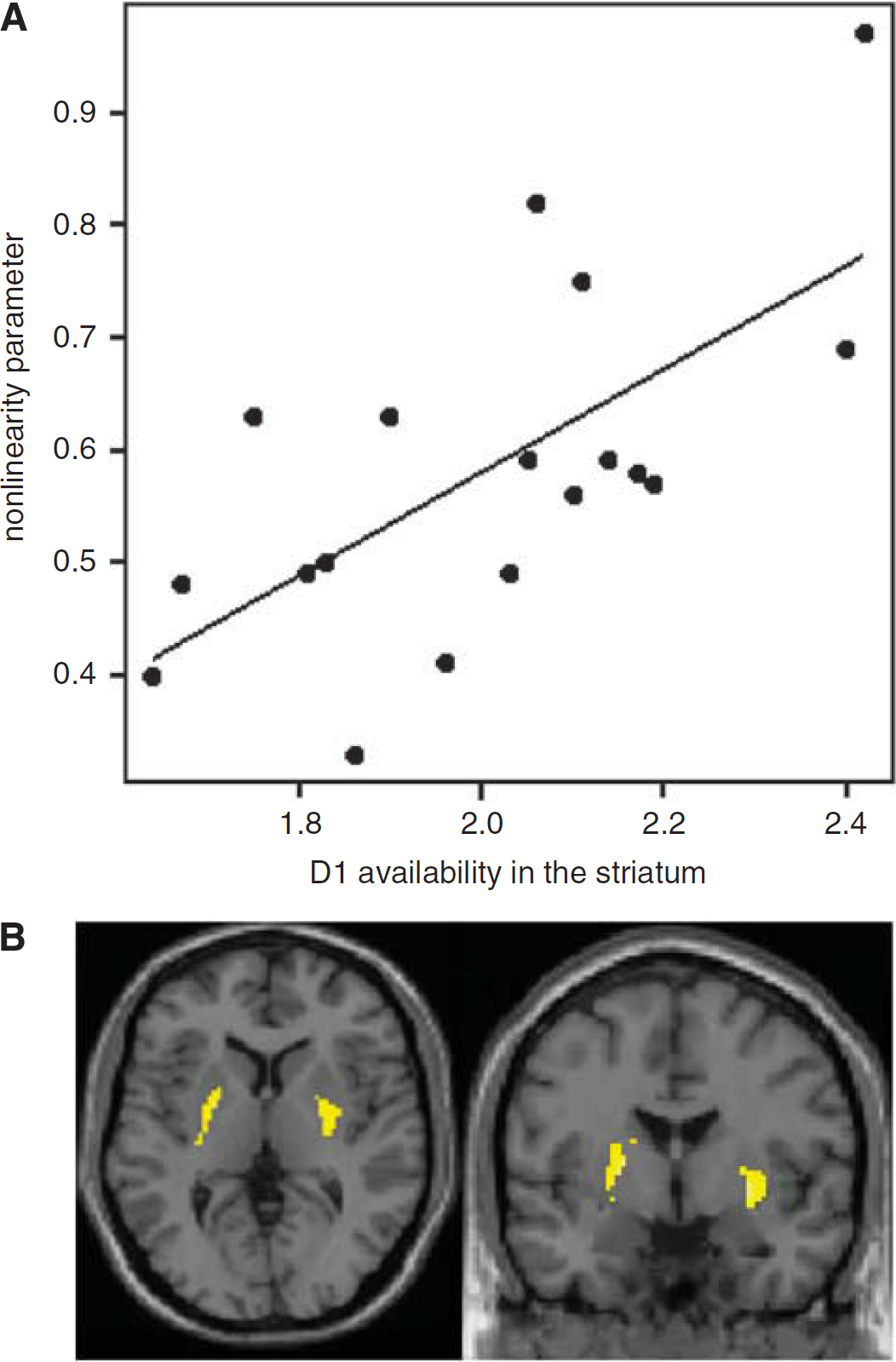
Correlation between nonlinearity of probabilities weighting and D1 receptor availability in the striatum. (
A neuroeconomics fMRI, using a simpler exposure-choice paradigm, showed that Prelec's nonlinearity parameter
Functional significance of individual difference in D1 receptors
All of our three studies mentioned above showed that individual differences in D1 receptor availability in the brain predicted the individual differences in brain functions (working memory/set shifting, emotional reaction, and decision making under risk) better than that of D2 receptor availability (Takahashi et al, 2008, 2010a,2010b). We do not think that dopamine D2 receptors have minimal roles in these brain functions. However, can we learn something from these studies showing the predominance of D1 receptors in terms of predicting these brain functions?
Dopamine neurons are known to show tonic firing and phasic (burst) firing, and in turn tonic and phasic dopamine release are induced, respectively (Grace, 1991; Grace et al, 2007). Phasic dopamine release in the striatum occurs during reward and reward-predicting stimuli (Grace, 1991; Schultz, 2007b). Phasic dopamine release in the amygdala is also induced in response to stress or emotional stimuli (Inglis and Moghaddam, 1999). Although both tonic and phasic dopamine release are necessary for PFC functions, phasic dopamine release has a crucial role in working memory and set shifting (Braver et al, 1999; Phillips et al, 2004). Thus, phasic dopamine release seems to be important for the brain functions that we investigated (working memory/set shifting, emotional reaction, and decision making under risk).
It has been shown that D1 receptors have much less affinity to endogenous dopamine than D2 receptors (Richfield et al, 1989). Furthermore, cortical and striatal D1 receptors are known to be predominantly extrasynaptic (Caille et al, 1996; Smiley et al, 1994). These facts suggest that D1-mediated neurotransmission is mainly governed by volume transmission (Dreher and Burnod, 2002; Garris et al, 1994), which might be induced by the phasic dopamine release from axonal terminals (Schultz, 2007a). Therefore, it can be suggested that available D1 receptors are preferentially stimulated by phasically released DA, whereas low-level baseline tonic dopamine release is sufficient for stimulating D2 receptors (Frank et al, 2007; Schultz, 2007b). A recent computational model also showed that phasic dopamine release primarily increases D1 occupancy, whereas D2 occupancy was less affected (Dreyer et al, 2010). Thus, these considerations lead us to believe that the variability of available D1 receptors might be more associated with individual differences in brain functions that require phasic dopamine release.
Clinical implications
Our previous PET study using [11C]SCH23390 revealed that, compared with normal controls, D1 receptors in PFC were decreased in schizophrenia, which was associated with poor performance on WCST (Okubo et al, 1997b). However, another PET study using [11C]NNC112 reported that increased D1 receptors in PFC were associated with working memory deficits in schizophrenia (Abi-Dargham et al, 2002). The same research group recently replicated increased D1 receptors in PFC of drug-naive schizophrenia patients (Abi-Dargham et al, 2011). The group also reported that PFC D1 receptor availability measured by [11C]NNC112 was significantly upregulated in chronic ketamine users, although no significant relationships were found between PFC D1 receptor availability and performance on working memory tests (Narendran et al, 2005).
It has been discussed that these inconsistent results might stem from several factors including differences in radioligands, but our more recent PET study measuring cortical D1 receptors with both [11C]SCH23390 and [11C]NNC112 in the same schizophrenia population showed that prefrontal D1 receptors were decreased in chronic schizophrenia regardless of radioligands (Kosaka et al, 2010). Still, the reasons for these inconsistent results need to be clarified in the future. An inverted U-shaped response might account for working memory deficits in schizophrenia patients, whether D1 receptors in PFC are increased or decreased in patients.
The central profile of most antipsychotics is the D2 receptor blockade property. Antipsychotics are reasonably effective in ameliorating positive symptoms in schizophrenia. However, negative symptoms and cognitive impairments of schizophrenia are typically not responsive to antipsychotic therapy. This has led to the investigation of alternative agents for the treatment of cognitive impairments in schizophrenia, and a body of data from animal and human studies support the utility of the D1 agonist (Buchanan et al, 2007; Okubo et al, 1997a). However, the efficacy of D1 agonists on cognitive impairments has not so far been proven due to several practical issues of drug development. In addition to these issues, we need to taken into account the fact that schizophrenia is a heterogeneous disorder. D1 receptor density might be different according to the type of the disease, changeable even in a single patient according to its stage (prodromal phase, first episode phase, and chronic phase). The inverted U-shaped property of D1 receptor stimulation might lead to bidirectional effect of D1 agonist depending on the type or stage of schizophrenia. Anhedonia or blunted affect is one of the central features of negative symptoms. Some neuroimaging studies have suggested that reduced amygdala activation was associated with these symptoms (Dowd and Barch, 2010; Takahashi et al, 2004). Therefore, similarly to the strategy for cognitive impairment, D1 agonist might be useful for restoring amygdala activation, and consequently improve these negative symptoms.
Misestimating risk could lead to disadvantaged choices such as initiation of drug use/gambling and transition to regular drug use/gambling (Kreek et al, 2005). Our studies have shown that people with lower striatal D1 receptor availability tend to misestimate the weight of probabilities, and in particular, to overestimate low probabilities of winning gambles (risk seeking). This finding led us to the intuitive conjecture that D1 agonist, again, might be useful for easing misestimation of risk, and consequently beneficial for pathological gambling. However, on the contrary, clinical reports have indicated the association between dopamine agonist medication and the emergence of pathological gambling in Parkinson's disease patients (Gallagher et al, 2007). Although early reports implicated D3 receptor agonists as being most likely to induce pathological gambling in Parkinson's disease patients (Dodd et al, 2005), it has been reported that mixed D1/D2 receptor agonists can also promote pathological gambling (Lu et al, 2006). These clinical findings appear to challenge our prediction, but indeed they may not. Pathological gambling is a complex behavior, which has been related to failures in impulse control or response inhibition as observed in Parkinson's disease, but also to impaired decision making, including risky or ambiguous decision. Estimation of risk requires the latter high-level processing, and we would argue that this is related to striatal D1 receptor availability, leading to the following hypothesis: low-level striatal D1 receptor availability (which might in part be determined by genetic factors) is linked to a risk-seeking trait. The risk-seeking trait was reported to be linked to enhanced activation and DA release in the striatum during risk-seeking behavior (Leyton et al, 2002; St Onge and Floresco, 2009). Chronic exposure to unusually high release of DA by risk-seeking behavior might induce downregulation of D1 receptors (Moore et al, 1998; Yasuno et al, 2007). The further decrease in D1 receptor availability then leads to further risk seeking. Low-level striatal D1 receptor availability could therefore be a gateway to a vicious cycle, creating a predisposition to drug addiction and pathological gambling. Recently, circumstantial evidence to support this hypothesis has been reported. Martinez et al (2009), based on their PET study, suggested that reduced D1 receptor binding may be associated with an increased risk of relapse in cocaine addiction. Needless to say, this tentative hypothesis needs to be tested in future investigations, and we believe that understanding the molecular mechanism of extreme or impaired decision making will contribute to the assessment and prevention of drug and gambling addiction as well as the development of novel pharmacological therapies for these addictions. In conclusion, interdisciplinary approach combining molecular imaging techniques with cognitive neuroscience and clinical psychiatry will provide new perspectives for understanding the neurobiology of neuropsychiatric disorders and their innovative drug developments.
Footnotes
Acknowledgements
A part of this study is the result of ‘Integrated research on neuropsychiatric disorders’ carried out under the Strategic Research Program for Brain Sciences by the Ministry of Education, Culture, Sports, Science, and Technology of Japan (MEXT), a Grant-in-aid for scientific research on innovative areas: Prediction and Decision Making (23120009), a Grant by Takeda Science Foundation and a consignment expense for the Molecular Imaging Program on ‘Research Base for PET Diagnosis’ from MEXT.
Disclosure/conflict of interest
The authors declare no conflict of interest.