
Abstract
Sustained hemodynamic stresses, especially high blood flow, result in flow-induced outward vascular remodeling. Our previous study showed that macrophage depletion reduced flow-induced outward remodeling of the rat common carotid artery, indicating that macrophages are critical in flow-induced outward vascular remodeling. Macrophage is known to release proteinases, including matrix metalloproteinases (MMPs). Degradation and loosening of extracellular matrix by MMPs may facilitate vascular remodeling. Therefore, we assessed the functions of MMPs in flow-induced outward vascular remodeling by using the flow-augmented common carotid artery model in mice. We validated that ligation of the left common carotid artery increased blood flow and luminal diameter of the right common carotid artery without significant change in blood pressure of mice. To assess the functions of MMPs in flow-induced outward vascular remodeling, we used doxycycline (broad-spectrum MMP inhibitor), SB-3CT (selective MMP inhibitor), MMP-9 knockout mice, and MMP-12 knockout mice. Although there was only a trend for doxycycline treatment to reduce flow-induced outward vascular remodeling, SB-3CT treatment significantly reduced flow-induced outward vascular remodeling. In addition, flow-induced outward vascular remodeling was significantly reduced in MMP-9 knockout mice, but not in MMP-12 knockout mice. These data revealed that MMPs, especially MMP-9, are critical in flow-induced outward vascular remodeling.
Introduction
Flow-induced outward vascular remodeling is an adaptive process for blood vessels to normalize hemodynamic stress in response to an increase in blood flow (Gibbons and Dzau, 1994). The vasculatures sense the changes in blood flow and integrate these stress signals to trigger vascular remodeling (Gibbons and Dzau, 1994; Guyton and Hartley, 1985; Kamiya and Togawa, 1980; Kamiya et al, 1988; Langille and O'Donnell, 1986). This adaptive outward vascular remodeling is characterized by an increase in luminal diameter with relatively small changes in wall thickness. The basic mechanisms of this adaptive process may be involved in pathologic remodeling of arteries observed in atherosclerosis, aortic aneurysms, intracranial aneurysms, and brain arteriovenous malformations (Gibbons and Dzau, 1994; Glagov et al, 1987; Hashimoto et al, 2001, 2006).
Orchestrated effects of many cytokines and proteinases seem to control various tissue remodeling. Among the cytokines and proteinases expressed during these various types of tissue remodeling, matrix metalloproteinases (MMPs) seem to have critical role in tissue remodeling (Abbruzzese et al, 1998; Flamant et al, 2007; Galis and Khatri, 2002; Tronc et al, 2000). In our previous study using rats, we have shown that inflammatory cells, such as macrophages, have critical functions in adaptive outward remodeling of the common carotid artery in response to an increase in blood flow (Nuki et al, 2009). We have shown that the time course of MMP-9 upregulation coincided with that of macrophage infiltration during flow-induced outward remodeling of the common carotid artery, although MMP-2 levels remained unchanged (Nuki et al, 2009). These findings indicate that MMP-9, but not MMP-2, produced by macrophages may be involved in flow-induced outward vascular remodeling.
Flow-induced outward vascular remodeling involves structural reorganization of elastic lamina of the arterial wall. Therefore, in addition to MMP-9, another key MMP produced by macrophages, MMP-12 (macrophage elastase), may be important in flow-induced outward vascular remodeling by directly facilitating structural changes of the elastic lamina (Guo et al, 2008; Luttun and Carmeliet, 2001; Ding and Gray, 2001; Mizobe et al, 2008).
We hypothesized that MMPs produced by inflammatory cells, such as macrophages, are critical in flow-induced outward remodeling of the common carotid artery in response to an increase in blood flow. To examine the functions of MMPs in flow-induced outward remodeling of the common carotid artery, we used a flow-augmented common carotid artery model in mice. First, we validated the model for flow-induced outward remodeling of the common carotid artery in mice. Second, we assessed the effects of doxycycline, a broad-spectrum MMP inhibitor, on flow-induced outward vascular remodeling. Third, we assessed the effects of SB-3CT, a highly selective inhibitor of MMP-9 and MMP-2, on flow-induced outward vascular remodeling. Finally, we assessed the functions of MMP-9 and MMP-12 in flow-induced outward vascular remodeling using MMP-9 and MMP-12 knockout mice.
Materials and methods
All experiments were conducted in accordance with the guidelines approved by the University of California, San Francisco, Institutional Animal Care and Use Committee.
Validation of Model for Flow-Induced Outward Remodeling of the Right Common Carotid Artery in Mice
To study flow-induced outward vascular remodeling, we used a flow-augmented common carotid artery model in mice; this model involves ligation of the left common carotid artery. We have previously shown that ligation of the left common carotid artery augments the blood flow in the right common carotid artery in rats (Nuki et al, 2009). The advantage of this model is that it does not involve direct surgical manipulation of the blood vessel that we are studying and, hence, avoids significant inflammation or surgery-related changes (Nuki et al, 2009).
We assessed changes in luminal diameter and blood flow in the right common carotid artery and blood pressure after flow augmentation to validate this model. C57BL/6J male mice (18 to 24 g, 8 weeks old; Jackson Laboratory, Bar Harbor, MA, USA) underwent sham surgery or flow augmentation of the right common carotid artery by ligation of the left common carotid artery. For flow augmentation of the right common carotid artery, ligation of the left common carotid artery was performed through 3 to 5 mm skin incision at the left side of the neck under anesthesia with isoflurane. For sham surgery, the left common carotid artery was exposed, and a surgical suture was placed and left under the left common carotid artery without ligating the left common carotid artery. During ligation of the left common carotid artery or sham surgery, the vagus nerve was carefully separated from the carotid artery to avoid any potential effects on hemodynamic parameters.
Luminal diameter and blood flow in the right common carotid artery were measured using a small animal dedicated ultrasound imaging system (Vevo660; VisualSonics, Toronto, Ontario, Canada) at 1 mm below the carotid bifurcation using the M-mode image. B-mode image was used as a guide to obtain M-mode image and Doppler-mode image. M-mode imaging shows temporal changes in one-dimensional echo recording of the motion of the interfaces on the y-axis and time on the x-axis. For M-mode imaging, we used the following settings: frequency 35 mHz, ultrasound gain 14 dB, and frame rate 1,000 frames per second. B-mode imaging shows two-dimensional views of the interfaces. For the B-mode imaging, we used the following settings: frequency 35 mHz, ultrasound gain 4 dB, and frame rate 30 frames per second. Mean luminal diameter of the common carotid artery was calculated by dividing diameter-time integral (area between anterior and posterior walls) by time over nine cardiac cycles, and systolic luminal diameter of the common carotid artery was calculated as an average of systolic luminal diameter over nine cardiac cycles. Diameter-time integral was traced and measured using the M-mode image. Flow-velocity curve was measured with the Doppler-mode image. From the flow-velocity curve, VTI (velocity—time integral) was obtained. Blood flow was calculated as VTI × πr2 × cosθ (r, radius = mean luminal diameter/2; θ, Doppler beam angle against the blood flow).
Systolic blood pressure was measured using a noninvasive tail-cuff method (ADInstruments, Colorado Springs, CO, USA) as previously described (Cervenka et al, 2001; Krege et al, 1995; Nuki et al, 2009; Wang et al, 2005). In our preliminary experiments, we encountered awake C57BL/6J mice to have high locomotor activity and resistance to restrain even after training as previously described (Daugherty et al, 2000; Davis and King, 1967). Therefore, we anesthetized the mice with 1.5% of isoflurane, and systolic blood pressure was measured at the steady state (after 15 mins of equilibration with the end-tidal isoflurane concentration at 1%) to avoid confounding effects of locomotion and excitement on blood pressure measurement. Luminal diameter and blood flow in the right common carotid artery and blood pressure were measured at baseline, 3, 6, and 8 weeks after flow augmentation or sham surgery of the right common carotid artery. Baseline measurement was taken 1 day before flow augmentation and sham surgery. Five animals were studied longitudinally in both the flow augmentation and sham surgery groups.
Detection of MMP Activity in the Right Common Carotid Artery
To detect gelatinase activity, specifically MMP activity, in the right common carotid arteries, in situ zymography using gelatin as a substrate was performed as previously described (Nuki et al, 2009). The right common carotid artery was harvested 1 and 3 days after flow augmentation by ligation of the left common carotid artery or sham surgery. To assess the contribution of MMP activity to the gelatinase activity, we treated tissues with MMP inhibitor, 1,10-phenanthroline monohydrate (500 μmol/L, 1h). To examine colocalization of macrophages with gelatinase activity, immunofluorescence staining using a macrophage marker CD68 (AbD Serotec, Raleigh, NC, USA) was combined with in situ zymography.
Doxycycline Treatment
To assess the effects of doxycycline, a broad-spectrum MMP inhibitor, on flow-induced outward vascular remodeling, four groups of C57BL/6J male mice (18 to 24 g, 8 weeks old) (n = 5 in each group) were prepared; group 1, sham surgery (without flow augmentation); group 2, sham surgery (without flow augmentation) with doxycycline treatment; group 3, flow augmentation; and group 4, flow augmentation with doxycycline treatment. Mice received doxycycline (40 mg/kg per day) through drinking water from 1 day before any surgical procedure until the end of the experiment for 3 weeks, which was the time point when flow-induced outward remodeling of the common carotid artery reached a plateau in our validation study (details of validation study are described in the Results section). Previously, this dose administered through drinking water was used to successfully suppress MMPs involved in various roles in mice (Lee et al, 2006; Pyo et al, 2000; Xiong et al, 2008). Luminal diameter of the right common carotid artery was measured at baseline and 3 weeks after flow augmentation or sham surgery using ultrasound imaging technique.
SB-3CT Treatment
To assess the effects of SB-3CT, a highly selective inhibitor of MMP-9 and MMP-2 (Brown et al, 2000), on flow-induced outward vascular remodeling, four groups of C57BL/6J mice (18 to 24 g, 8 weeks old) (n = 5 in each group) were prepared; group 1, sham surgery (without flow augmentation) with vehicle treatment; group 2, sham surgery (without flow augmentation) with SB-3CT treatment; group 3, flow augmentation with vehicle treatment; and group 4, flow augmentation with SB-3CT treatment. SB-3CT was prepared at a final concentration of 17.5 mg/mL in vehicle of 25% dimethyl sulfoxide/65% polyethylene glycol 200/10% water. Each mouse was administered 70 mg/kg per day of SB-3CT or only vehicle through single daily intraperitoneal injection starting from 2 days before the surgery until the end of the experiment. Luminal diameter of the right common carotid artery was measured at baseline and 3 weeks after flow augmentation or sham surgery.
Flow-Induced Outward Vascular Remodeling in MMP-9 and MMP-12 Knockout Mice
To assess the functions of MMP-9 and MMP-12 on flow-induced outward vascular remodeling, MMP-9 knockout mice (FVB.Cg-Mmp9tm1Tvu/J; Vu et al, 1998), MMP-12 knockout mice (B6.129X-Mmp12tm1Sds/J; Shipley et al, 1996), and corresponding wild-type mice were used (18 to 24 g, 8 weeks old; Jackson Laboratory). Four groups of mice (n = 5 in each group) were prepared for each experiment; group 1, wild-type mice with sham surgery (without flow augmentation); group 2, knockout mice with sham surgery (without flow augmentation); group 3, wild-type mice with flow augmentation; and group 4, knockout mice with flow augmentation. For each experiment, mice in groups 1 and 3 were wild-type mice from the same strain as the knockout mice in groups 2 and 4. Although our preliminary experiments showed that flow-induced outward remodeling of the common carotid artery plateaued at 3 weeks time point in the wild-type mice, we considered the possibility that MMP knockout mice have slower progression of outward vascular remodeling rather than suppressed remodeling. Therefore, we longitudinally followed flow-induced outward vascular remodeling by measuring luminal diameter of the flow-augmented common carotid artery at baseline, 3, and 6 weeks after flow augmentation or sham surgery. Body weight was also measured at baseline, 3, and 6 weeks after surgery.
Statistical Analysis
We used analysis of variance followed by Fisher's least significant difference test, which was performed as a post hoc analysis. When repeated measurements were performed, analysis of variance with a repeated measurement design followed by Fisher's least significant difference test was used. All the data were presented as mean ± s.d. Statistical significance was taken at P < 0.05.
Results
Validation of Flow-Induced Outward Vascular Remodeling of the Right Common Carotid Artery Model in Mice
Figure 1A shows a representative ultrasound B-mode image of the right common carotid artery and its bifurcation. Luminal diameter of the right common carotid artery was measured using the M-mode image (Figure 1B). Blood flow velocity was measured using the Doppler-mode image (Figure 1C).
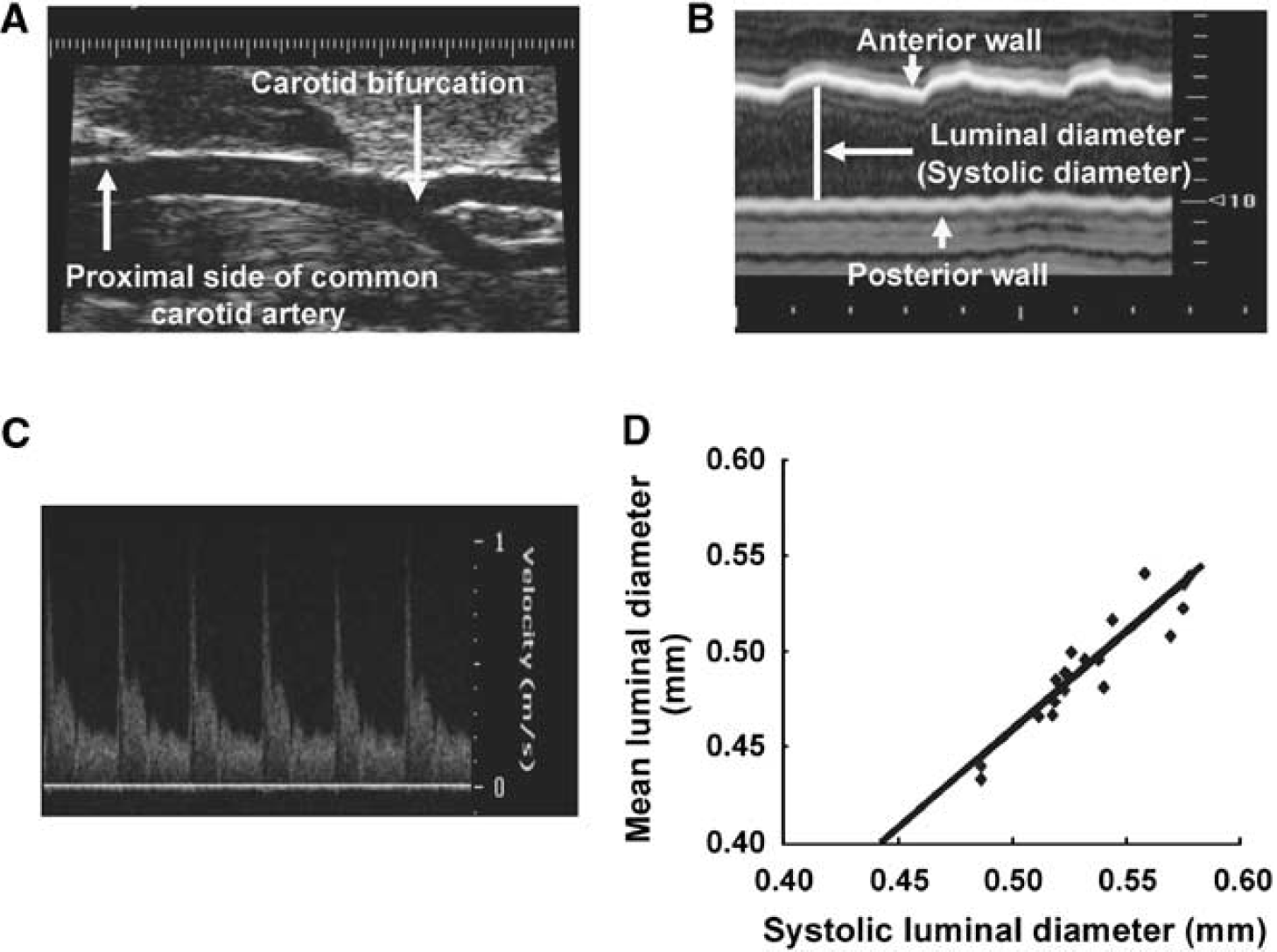
Ultrasound images of the mouse common carotid artery. (
Figure 1D shows the correlation, close linear relationship, between mean luminal diameter and systolic luminal diameter at baseline and after flow augmentation of the right common carotid artery (y = 1.03x − 0.06, R2 = 0.86, P<0.05), indicating that systolic luminal diameter can be used as a surrogate for mean luminal diameter in studying flow-induced outward remodeling of the mouse common carotid artery.
As shown in Figure 2A, blood flow in the right common carotid artery increased by 60% from baseline at 3 weeks after flow augmentation by ligation of the left common carotid artery (baseline versus 3 weeks after flow augmentation: 34.1 ± 2.2 versus 54.2 ± 4.3 mm3/s, P<0.05 compared with baseline) and sustained for at least 8 weeks. Although blood flow levels were higher than baseline at 3, 6, and 8 weeks (baseline versus 3, 6, and 8 weeks: 34.1 ± 2.2 versus 54.2 ± 4.3, 60.4 ± 16, and 68.2 ± 38.8 mm3/s, respectively, P<0.05 compared with baseline), there was no significant difference in blood flow in the flow-augmented right common carotid artery between 3, 6, and 8 weeks. In addition, there was no significant difference in blood flow in the right common carotid artery between baseline, 3, 6, and 8 weeks after sham surgery (data not shown).
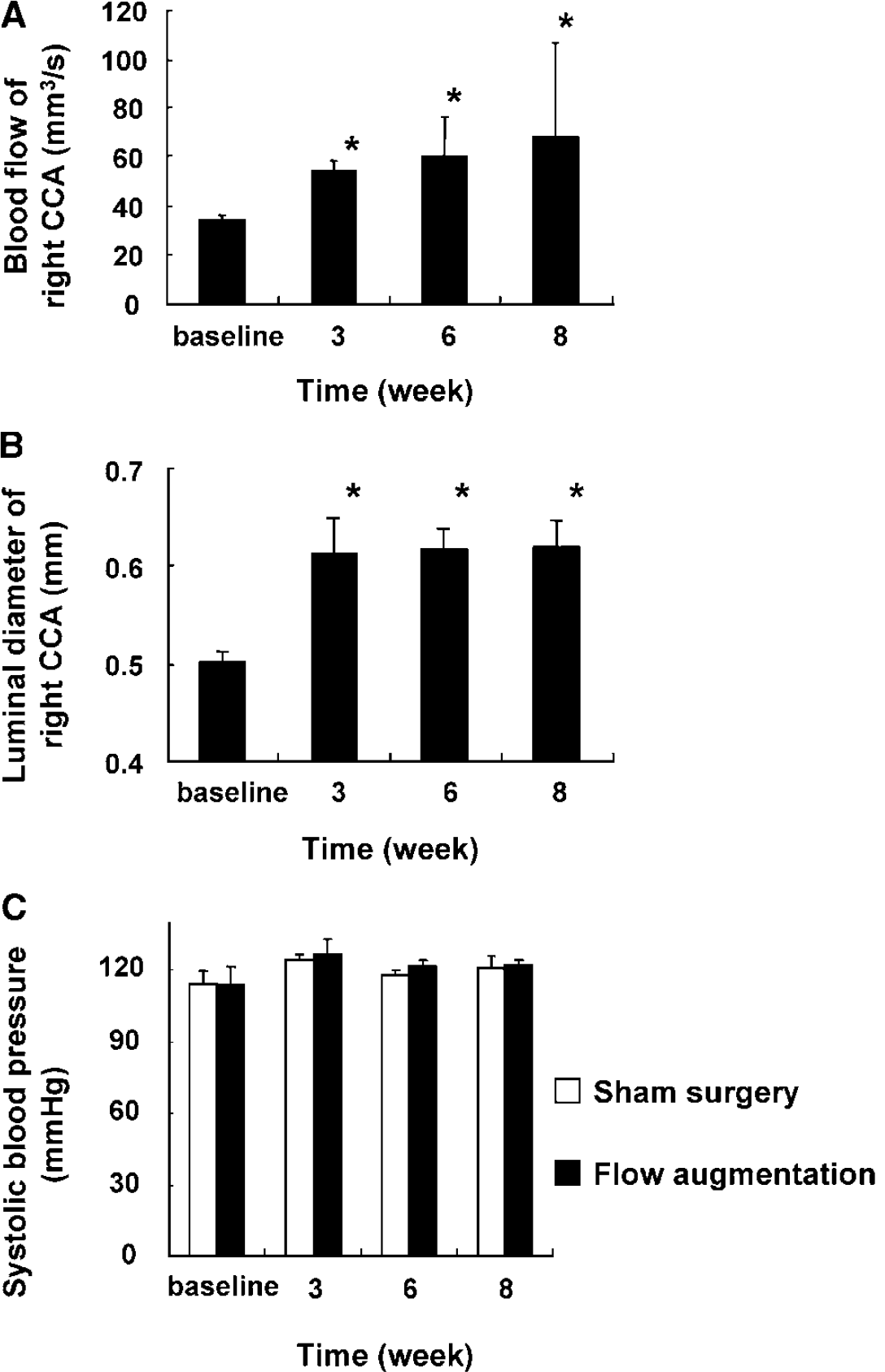
Blood flow and luminal diameter of the right common carotid artery and systolic blood pressure at baseline and after flow augmentation. (
Figure 2B shows the time course of luminal diameter of the right common carotid artery at baseline, 3, 6, and 8 weeks after flow augmentation. There was an increase in luminal diameter of the flow-augmented common carotid artery from baseline to 3 weeks, and this increase reached a plateau after 3 weeks. Luminal diameter of the flow-augmented common carotid artery at 3, 6, and 8 weeks was larger than baseline (baseline versus 3, 6, and 8 weeks: 0.50 ± 0.02 versus 0.61 ± 0.02, 0.62 ± 0.03, and 0.62 ± 0.03 mm, respectively, P < 0.05 compared with baseline). There was no significant difference in luminal diameter of the right common carotid artery between baseline, 3, 6, and 8 weeks after sham surgery.
Figure 2C shows the time course of systolic blood pressure in mice. There was no significant difference in blood pressure at all measured time points among the groups of mice that underwent sham surgery and flow augmentation of the right common carotid artery. Flow augmentation by ligation of the left common carotid artery did not affect blood pressure.
In summary, in this flow-augmented common carotid artery model using mice, there was a sustained increase in blood flow with an increase in luminal diameter of the flow-augmented right common carotid artery by ligation of the left common carotid artery. Furthermore, ligation of the left common carotid artery did not affect systolic blood pressure, and there was no significant difference in blood flow and luminal diameter of the right common carotid artery and blood pressure in mice between before and after sham surgery. These findings were consistent with previous studies using rats (De Ciuceis et al, 2005; Nuki et al, 2009; Schiffers et al, 2000; Tronc et al, 2000). This flow-augmented common carotid artery model can be, therefore, used to study flow-induced outward vascular remodeling in mice.
Presence and Colocalization of MMP Activity with Macrophages in Flow-Augmented Common Carotid Artery
As shown in Figure 3A, the right common carotid artery from sham surgery group showed low levels of gelatinase activity that was detected only between elastic laminae, likely because of constitutive expression of MMP-2 by smooth muscle cells as previously described (Nuki et al, 2009; Southgate et al, 1999; Whatling et al, 2004; Zaltsman and Newby, 1997). In comparison, the flow-augmented right common carotid artery showed gelatinase activity between the elastic laminae and in the adventitia at day 3 (Figure 3B), consistent with previous findings (Nuki et al, 2009). Furthermore, treatment of tissues with MMP inhibitor, 1,10-phenanthroline monohydrate, inhibited gelatinase activity in the flow-augmented right common carotid artery, indicating that the majority of gelatinase activity originated from MMPs (data not shown).
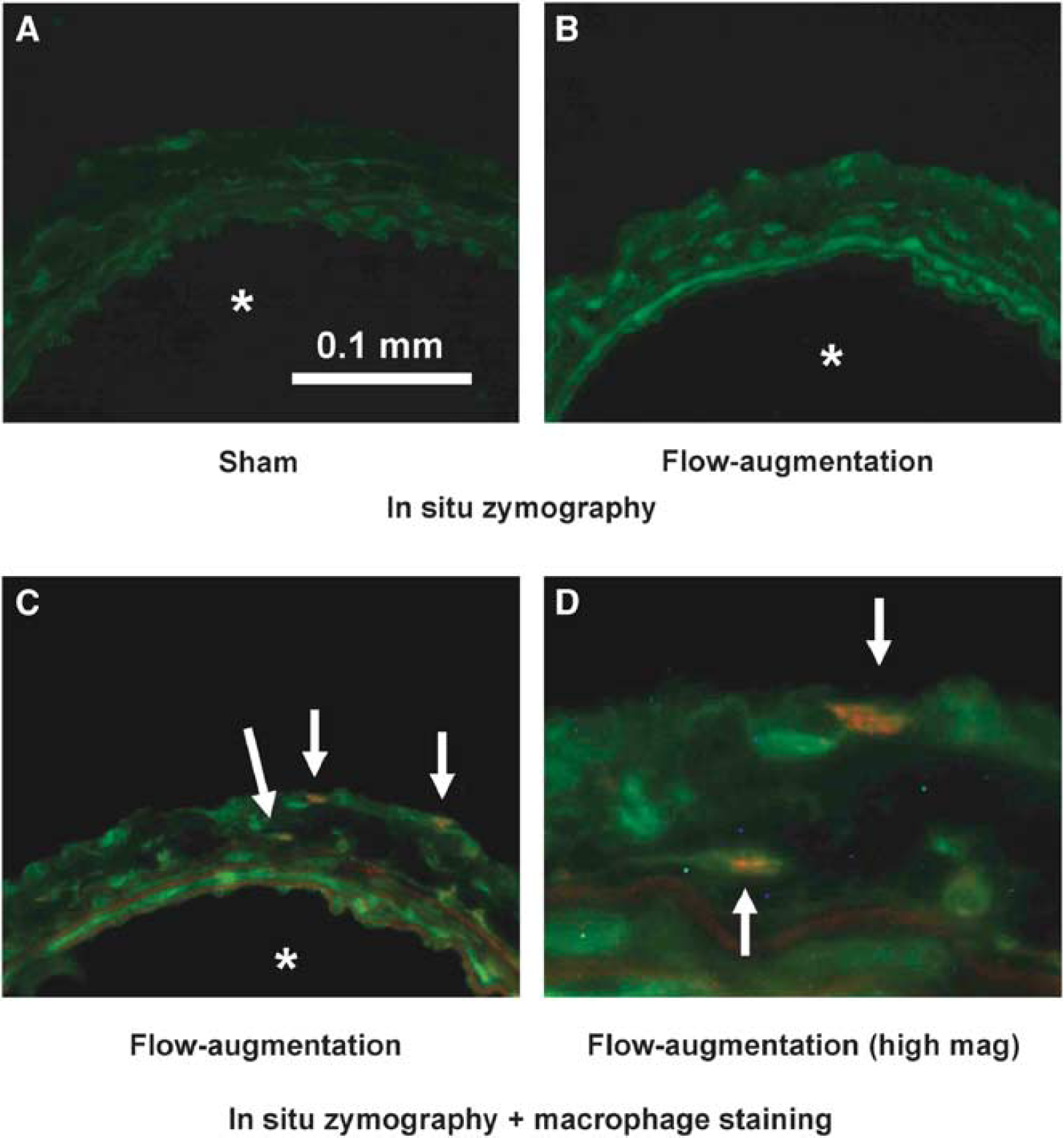
Matrix metalloproteinase expression in flow-augmented common carotid artery using in situ zymography. (
Because macrophages can secrete various proteinases including MMPs, we examined colocalization of gelatinase activity with macrophages by combining in situ zymography with immunofluorescence staining for macrophages. In the flow-augmented right common carotid artery, macrophages were present in the adventitia, consistent with previous findings (Capers et al, 1997; Gavrila et al, 2005; Nuki et al, 2009; Weiss and Taylor, 2008). In addition, macrophages were found to be colocalized with MMP activity in the adventitia of the flow-augmented right common carotid artery (Figures 3C and 3D). Yellow in Figure 3C and 3D represents colocalization of MMP activity and macrophage. All macrophages that were detected in the tissues expressed strong MMP activity; however, there were faint MMP activities by other cell types.
Effects of Doxycycline, a Broad-Spectrum MMP Inhibitor, on Flow-Induced Outward Vascular Remodeling
To assess the effects of doxycycline, a broad-spectrum MMP inhibitor, on flow-induced outward vascular remodeling, luminal diameter of the right common carotid artery was measured at baseline and 3 weeks after flow augmentation or sham surgery in mice with or without doxycycline treatment (Figure 4A). Our validation study described above showed that at 3 weeks after flow augmentation, flow-induced outward vascular remodeling reached a plateau. Doxycycline treatment did not affect luminal diameter of the right common carotid artery in sham surgery groups (group 1 versus group 2, P = 0.64) (Figure 4A). Luminal diameter of the right common carotid artery in mice that underwent flow augmentation was significantly larger than that of mice that underwent sham surgery (group 3 versus group 1: 0.61 ± 0.02 versus 0.53 ± 0.02 mm, P < 0.05; group 4 versus group 2: 0.59 ± 0.03 versus 0.50 ± 0.02 mm, P < 0.05) (Figure 4A). Between the groups of mice that underwent flow augmentation, there was a trend for flow-induced outward remodeling of the right common carotid artery to be reduced in mice that received doxycycline treatment compared with mice without doxycycline treatment (group 4 versus group 3: 0.59 ± 0.03 versus 0.61 ± 0.02 mm, P = 0.15) (Figure 4A). There was no difference in body weight among the four groups at any time point (group 1 versus group 2 versus group 3 versus group 4: 24.4 ± 1.7 versus 25.4 ± 0.8 versus 22.8 ± 1.7 versus 25.5 ± 2.4 g at 3 weeks).
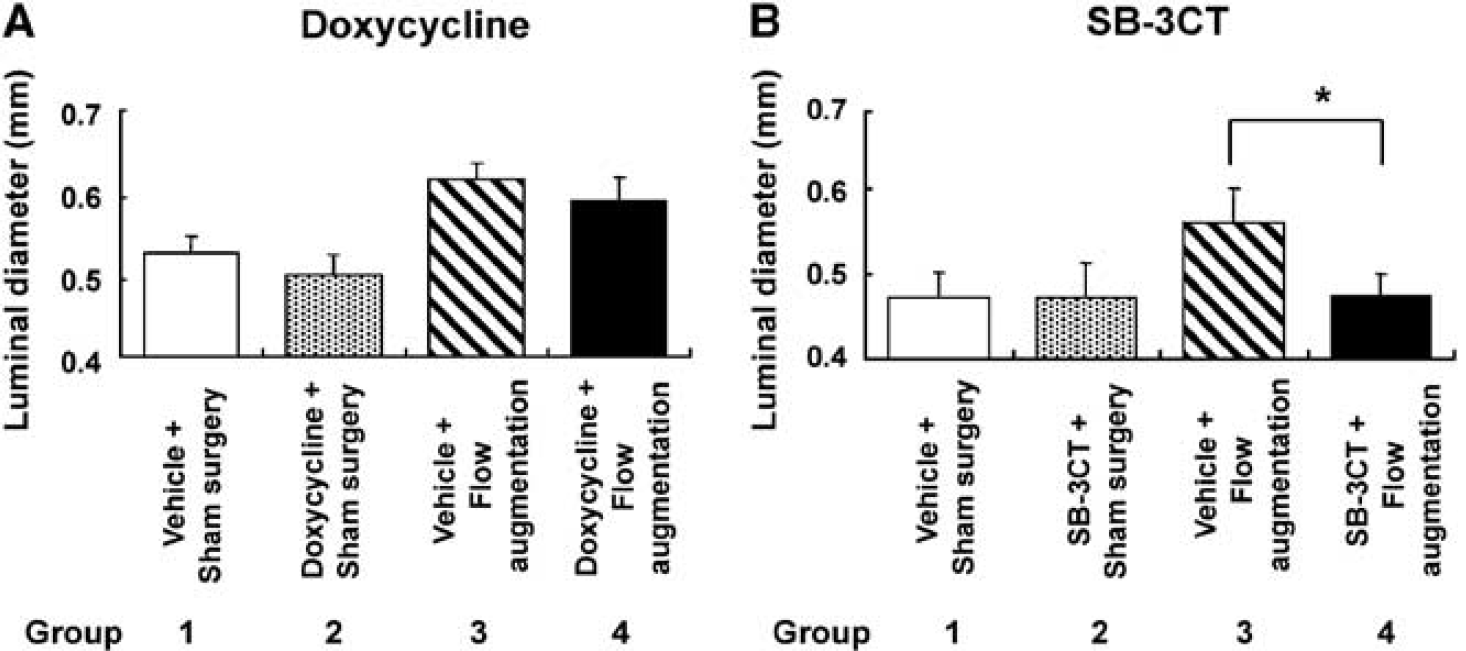
Effects of MMP inhibitors on flow-induced outward vascular remodeling. (
SB-3CT, a Selective MMP Inhibitor, Significantly Reduced Flow-Induced Outward Vascular Remodeling
Because doxycycline is a broad-spectrum MMP inhibitor and has effects other than inhibition of MMPs, we assessed effects of SB-3CT, a selective inhibitor of MMP-9 and MMP-2, on flow-induced outward vascular remodeling by measuring luminal diameter of the right common carotid artery after flow augmentation or sham surgery. As shown in Figure 4B, luminal diameter of the right common carotid artery was measured at baseline and 3 weeks after flow augmentation or sham surgery in mice with or without SB-3CT treatment.
SB-3CT treatment did not affect luminal diameter of the right common carotid artery in sham surgery groups (group 1 versus group 2, P = 0.93) (Figure 4B). Flow augmentation by ligation of the left common carotid artery significantly increased luminal diameter of the right common carotid artery in the group of mice that received vehicle treatment (group 3 versus group 1: 0.57 ± 0.04 versus 0.47 ± 0.03 mm, P < 0.05). Between the groups of mice that underwent flow augmentation, the mice that received SB-3CT showed significantly reduced flow-induced outward remodeling of the right common carotid artery compared with the mice that received vehicle treatment (group 4 versus group 3: 0.48 ± 0.03 versus 0.57 ± 0.04 mm, P < 0.05) (Figure 4B). There was no difference in body weight among the four groups at any time point (group 1 versus group 2 versus group 3 versus group 4: 21.2 ± 1.4 versus 20.6 ± 1.7 versus 19.9 ± 1.7 versus 18.8 ± 1.1 g at 3 weeks).
Reduction of Flow-Induced Outward Vascular Remodeling in MMP-9 Knockout Mice
The experiments using doxycycline and SB-3CT strongly indicated that MMPs are important in flow-induced outward vascular remodeling. Our previous data showed that the time course of MMP-9 upregulation, but not MMP-2, coincided with that of macrophage infiltration into the flow-augmented common carotid artery (Nuki et al, 2009). MMP-2 was constitutively expressed in smooth muscle cells without being affected by flow augmentation. In addition, our preliminary experiment using morphometric analysis of histologic samples showed no difference in luminal diameter of the flow-augmented common carotid artery between MMP-2 knockout mice and wild-type mice (data not shown).
Therefore, we chose to assess flow-induced outward vascular remodeling in MMP-9 knockout mice by measuring luminal diameter of the right common carotid artery in MMP-9 knockout mice and corresponding wild-type mice at baseline, 3, and 6 weeks after flow augmentation or sham surgery (Figure 5A).
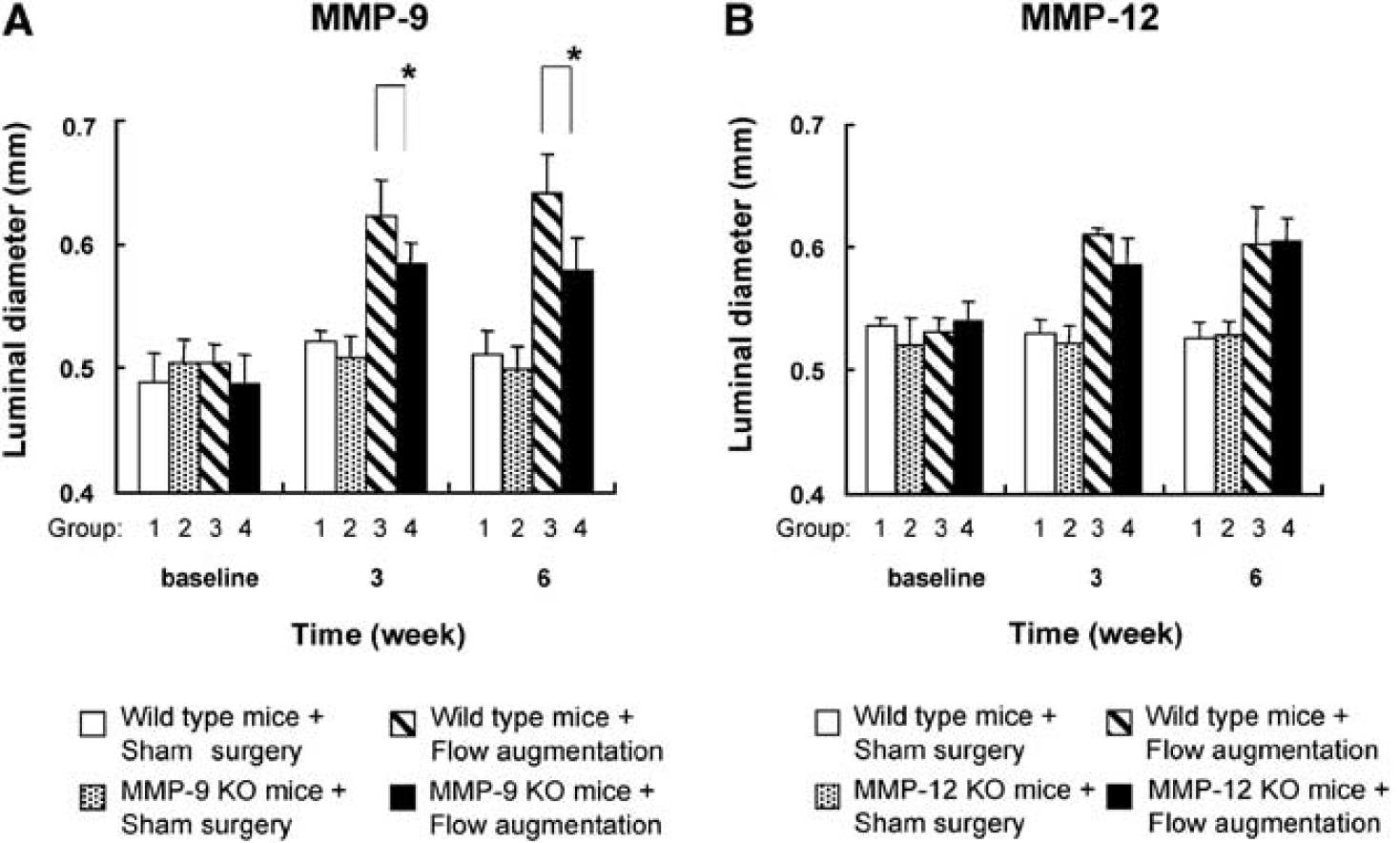
Effects of MMP-9 and MMP-12 on flow-induced outward vascular remodeling. (
There was no significant difference in luminal diameter of the right common carotid artery between MMP-9 knockout mice and wild-type mice at baseline (P = 0.36). At 3 and 6 weeks, luminal diameter of the flow-augmented right common carotid artery was significantly larger than that of sham surgery (group 3 versus group 1 and group 4 versus group 2, both comparisons and at 3 and 6 weeks, P < 0.05). Between the groups of mice that underwent flow augmentation (groups 3 and 4), luminal diameter of the right common carotid artery in MMP-9 knockout mice was significantly less than that of wild-type mice at 3 and 6 weeks (group 4 versus group 3 at 3 weeks: 0.58 ± 0.02 versus 0.62 ± 0.03 mm, P < 0.05; group 4 versus group 3 at 6 weeks: 0.58 ± 0.03 versus 0.64 ± 0.03 mm, P < 0.05), showing reduced flow-induced outward remodeling of the right common carotid artery in MMP-9 knockout mice. There was no significant difference in luminal diameter of the right common carotid artery in sham surgery-operated wild-type mice at baseline, 3, and 6 weeks. There was no difference in body weight among the four groups at any time point (group 1 versus group 2 versus group 3 versus group 4: 27.9 ± 1.5 versus 27.1 ± 1.4 versus 24.9 ± 0.9 versus 27.0 ± 1.0 g at 6 weeks).
Lack of MMP-12 did not Reduce Flow-Induced Outward Vascular Remodeling
Our previous study showed critical functions of macrophages in flow-induced outward vascular remodeling (Nuki et al, 2009). As described above, lack of MMP-9 reduced flow-induced outward vascular remodeling; however, outward vascular remodeling was not suppressed. In addition to MMP-9, MMP-12 is another major MMP produced by macrophages, and it may have a critical function in outward vascular remodeling (Banda and Werb, 1981; Wu et al, 2000; Yu et al, 2007). Therefore, we assessed flow-induced outward vascular remodeling in MMP-12 knockout mice.
Figure 5B shows a time course of luminal diameter of the right common carotid artery in MMP-12 knockout mice and corresponding wild-type mice. There was no significant difference in luminal diameter of the right common carotid artery at baseline between MMP-12 knockout and wild-type mice (P = 0.11). Luminal diameter of the right common carotid artery in mice that underwent flow augmentation was significantly larger than that of mice that underwent sham surgery at 3 and 6 weeks (group 3 versus group 1 and group 4 versus group 2, both comparisons at 3 and 6 weeks, P < 0.05). However, between the groups of mice that underwent flow augmentation (groups 3 and 4), there was no difference in luminal diameter of the right common carotid artery between MMP-12 knockout and wild-type mice either at 3 or 6 weeks after flow augmentation (group 4 versus group 3 at 3 weeks: 0.59 ± 0.02 versus 0.61 ± 0.01 mm, P = 0.09; group 4 versus group 3 at 6 weeks: 0.61 ± 0.02 versus 0.60 ± 0.03 mm, P = 0.89) (Figure 5B), showing that lack of MMP-12 did not have any significant effect on flow-induced outward vascular remodeling. There was no significant difference in luminal diameter of the right common carotid artery in sham surgery-operated wild-type mice at baseline, 3, and 6 weeks. There was no difference in body weight among the four groups at any time point (group 1 versus group 2 versus group 3 versus group 4: 26.6 ± 1.6 versus 27.9 ± 1.3 versus 27.2 ± 2.2 versus 27.5 ± 2.3 g at 3 weeks).
Discussion
In this study, we have showed the critical functions of MMPs in flow-induced outward remodeling of the common carotid artery in mice. First, we validated the model for flow-induced outward remodeling of the common carotid artery in mice. In this model, use of an ultrasound imaging technique allowed in vivo longitudinal assessment of luminal diameter of the common carotid artery. Second, we have shown that pharmacologic inhibition of MMPs by SB-3CT, a highly selective inhibitor of MMP-9 and MMP-2, reduced flow-induced outward vascular remodeling in mice. Third, we assessed the functions of MMP-9 and MMP-12, two major MMPs that are produced by macrophages, and found that flow-induced outward vascular remodeling was reduced in mice lacking MMP-9, but not in mice lacking MMP-12. Previously, we have shown that infiltrating macrophages in the remodeling common carotid artery expressed MMPs and that systemic depletion of macrophages significantly reduced flow-induced outward remodeling of the rat common carotid artery (Nuki et al, 2009). Taken together, MMPs that are produced by macrophages, especially MMP-9, seem to have critical functions in flow-induced outward vascular remodeling.
In this study, we tested the effects of two inhibitors of MMPs, SB-3CT and doxycycline, on flow-induced outward vascular remodeling. SB-3CT, but not doxycycline, caused a significant reduction of flow-induced outward vascular remodeling. In this study, we used a similar dose of doxycycline through drinking water as used by others to inhibit effectively the effects of MMPs in mice (Lee et al, 2003; Pyo et al, 2000; Xiong et al, 2008). The difference between the effects of SB-3CT and doxycycline on flow-induced outward vascular remodeling may be because of their different specificities and potencies toward different MMPs. Although SB-3CT is a highly selective inhibitor of MMP-9 and MMP-2, doxycycline is a broad-spectrum inhibitor of MMPs. In addition, doxcycline has other effects in addition to inhibiting MMPs, including modulations of various aspects of inflammation (Beekman et al, 1997; Kim et al, 2005; Lee et al, 2003). These additional effects of doxycycline may have diminished or masked its MMP inhibitory effects.
Nevertheless, the reduction of flow-induced outward vascular remodeling by SB-3CT indicates that MMPs are critical for flow-induced outward vascular remodeling. Previously, we have shown that the time course of MMP-9 upregulation, but not MMP-2, during flow-induced outward remodeling of the rat common carotid artery coincided with that of macrophage infiltration (Nuki et al, 2009). MMP-2 was constitutively expressed in smooth muscle cells without being affected by flow augmentation. Our preliminary study using morphometric analysis of histologic samples showed that there was no difference in luminal diameter of the flow-augmented common carotid artery between MMP-2 knockout and wild-type mice. More importantly, we found that flow-induced outward remodeling of the common carotid artery was reduced in MMP-9 knockout mice using a longitudinal study with an ultrasound imaging technique. These data strongly suggest that MMP-9, but not MMP-2, is critical for flow-induced outward vascular remodeling. Nonetheless, there remains a limitation of this study that we did not directly assess flow-induced outward remodeling of the common carotid artery in MMP-2 knockout mice, using the serial, longitudinal assessment with the ultrasound imaging.
Reduction of vascular remodeling of the flow-augmented common carotid artery in SB-3CT-treated mice was more prominent than in MMP-9 knockout mice. Lower reduction of vascular remodeling in MMP-9 knockout mice may have been because of unknown developmental abnormalities or compensation. Such unknown consequences of MMP-9 gene abrogation may have contributed to the incomplete suppression of flow-induced outward vascular remodeling. Furthermore, the duration of SB-3CT experiment was only for 3 weeks whereas MMP-9 knockout mice experiment was assessed for 6 weeks in addition to 3 weeks. Reduction of flow-induced outward vascular remodeling by SB-3CT may have become less apparent at 6 weeks.
Flow-induced outward vascular remodeling involves structural changes in elastic lamina of the vascular wall (Guo et al, 2008; Luttun and Carmeliet, 2001). Therefore, we investigated the functions of MMP-12, another major MMP produced by macrophages. Although MMP-9 is the main gelatinase produced by macrophages, MMP-12 is a macrophage elastase with little gelatinase activity. Among the MMPs produced by macrophages, MMP-12 could be the responsible proteinase that directly facilitates structural changes of elastic lamina resulting in flow-induced outward vascular remodeling (Ding and Gray, 2001; Mizobe et al, 2008). However, in our experiment, a lack of MMP-12 did not have a significant effect on flow-induced outward vascular remodeling.
Taken together, macrophages seem to be critical in flow-induced outward vascular remodeling through production and activation of MMP-9, whereas MMP-12, macrophage elastase, has little role. Macrophagederived MMP-9 may exert its effects by direct digestion of extracellular matrix and indirect effects by interaction with other cytokines or enzymes to trigger and facilitate flow-induced outward vascular remodeling (Corry et al, 2004; Dumont et al, 2007; Sternlicht and Werb, 2001).
It should be noted that in our study we did not measure the change in blood flow in the common carotid artery in response to ligation of the contralateral common carotid artery in mice that received MMP inhibitors or in MMP knockout mice. We cannot exclude the possibility that the increase in the common carotid artery blood flow may be affected by MMP inhibitors or abrogation of MMP genes. However, the acute increase in blood flow, which triggers the outward vascular remodeling in this model, is primarily a function of the downstream vascular resistance that is largely determined by the vascular anatomy (Nuki et al, 2009). Our preliminary studies showed no apparent differences in the macroscopic anatomy of cerebral arteries between MMP knockout mice and the corresponding wild-type mice, consistent with a previous report (Asahi et al, 2000). It is also unlikely that MMP inhibitor treatments, which were started only 1 to 3 days before the flow augmentation, changed the vascular anatomy.
In this study, we have not examined the effects of MMP inhibition on levels of circulating leukocytes that can potentially contribute to flow-induced outward vascular remodeling. It still remains to be determined what cell types at which locations are the primary target of the MMP inhibition in this setting. It is possible that by decreasing the levels of circulating leukocytes or modulating their MMP-dependent functions, instead of acting on the infiltrated leukocytes—tissue leukocytes—the MMP inhibition could have reduced flow-induced outward vascular remodeling.
In this study, we used flow augmentation of the common carotid artery by ligation of the contralateral common carotid artery. This model has few unique features that make it suitable for studying functions of inflammatory cells in flow-induced outward vascular remodeling. First, although the carotid artery ligation procedure has been widely used to study vascular remodeling triggered by flow cessation, it has not been used to study vascular remodeling in response to flow augmentation; previous studies focused on effects of flow cessation on the ligated side. Second, the models previously used to study vascular remodeling in response to flow augmentation involved direct surgical manipulation of the blood vessels that were being studied (Greenhill and Stehbens, 1983; Korshunov and Berk, 2003; Stehbens, 1974; Tronc et al, 2000). In such models, inflammation caused by surgical procedures could have confounded their studies, and it is difficult to separate effects of surgical inflammation from effects of inflammatory cells activated by flow augmentation. The advantage of our model is that it did not involve direct surgical manipulation of the blood vessel that we were studying and, hence, avoided significant inflammation or surgery-related changes.
In summary, our data showed the critical functions of MMP-9 in flow-induced outward vascular remodeling. Infiltrating macrophages into the flow-augmented common carotid artery may be involved in key processes of flow-induced outward vascular remodeling, mainly by its elaboration and activation of MMP-9.
Footnotes
The authors state no conflict of interest.