
Abstract
Hemoglobin is the major protein in red blood cells and transports oxygen from the lungs to oxygen-demanding tissues, like the brain. Mechanisms that facilitate the uptake of oxygen in the vertebrate brain are unknown. In invertebrates, neuronal hemoglobin serves as intracellular storage molecule for oxygen. Here, we show by immunohistochemistry that hemoglobin is specifically expressed in neurons of the cortex, hippocampus, and cerebellum of the rodent brain, but not in astrocytes and oligodendrocytes. The neuronal hemoglobin distribution is distinct from the neuroglobin expression pattern on both cellular and subcellular levels. Probing for low oxygen levels in the tissue, we provide evidence that hemoglobin α-positive cells in direct neighborhood with hemoglobin α-negative cells display a better oxygenation than their neighbors and can be sharply distinguished from those. Neuronal hemoglobin expression is upregulated by injection or transgenic overexpression of erythropoietin and is accompanied by enhanced brain oxygenation under physiologic and hypoxic conditions. Thus we provide a novel mechanism for the neuroprotective actions of erythropoietin under ischemic—hypoxic conditions. We propose that neuronal hemoglobin expression is connected to facilitated oxygen uptake in neurons, and hemoglobin might serve as oxygen capacitator molecule.
Introduction
Hemoglobin (Hb) belongs to a large superfamily of globins which originates from a common ancestor that evolved in bacteria (Vinogradov and Moens, 2008). Besides Hb, vertebrate globins include myoglobin, neuroglobin (Ngb), histoglobin, and globin X (Roesner et al, 2005). Globins are found in all kingdoms of life as a part of the basic molecular setup of many organisms (Hardison, 1996).
Evolutionary modifications around the highly conserved ‘globin fold’ have generated specialized globins with different functions, ranging from the transport of H2S to enzymatic NO reactions and radical elimination (Hardison, 1996; Vinogradov and Moens, 2008). The best-studied function is the reversible oxygen binding of vertebrate Hb, a heterotetramer of two α- and two β-chains in erythrocytes that delivers O2 to the tissue. In the human brain, the recent discovery of Ngb, a distantly related globin that shares less than 25% protein sequence similarity with vertebrate Hb, confirmed the existence of globins in vertebrate neurons (Burmester et al, 2000), although its physiologic function is still a matter of debate (Hankeln et al, 2005).
The globins that are expressed in the nervous system of molluscs like the clam
The hormone erythropoietin (EPO) stimulates the maturation of red blood cells and increases Hb levels in the blood. EPO is also expressed in the brain and has neuroprotective functions. In two previous gene expression screens in the brain of either EPO-overexpressing or EPO-injected mice (Maurer et al, 2002, 2004b), we detected the upregulation of α- and β-globin transcripts despite careful transcardial perfusion of the mice, and thus little possibility of erythrocyte or reticulocyte contamination. We therefore sought to determine whether Hb might be expressed in endogenous cells in the brain.
Materials and methods
Animals
Animal procedures adhered to German law and were approved by the relevant regulatory authorities (Regierungspräsidium Karlsruhe), which are concordant with the NIH guidelines on the use of laboratory animals. Wistar rats, C57BL/6 and NMRI mice were purchased from Charles River (Sulzfeld, Germany), EPO-transgenic Tg(PDGFB-EPO)321Zbz mice were kindly provided by Professor Dr Max Gassmann, University of Zürich, Switzerland, and bred at the animal core facility, University of Heidelberg.
Hypoxia
Female C57BL/6-mice were injected with 60 mg/kg pimonidazole (Hypoxyprobe-1; NPI, Burlington, MA, USA) and exposed to a gas mixture of 8% O2 and 92% O2 for 30 mins. The animals were transcardially perfused and the brains were fixed with 4% paraformaldehyde, removed and embedded in paraffin.
Primary cortical E18 neuronal cultures of Wistar rats were seeded on cover slips. After 1 week, the cover slips were transferred into medium containing 60 μg/mL pimonidazole and 0.8% O2. The cover slips were removed after 2, 5, 10, 15, and 30 mins and immediately fixed in 4% paraformaldehyde.
Oxygen levels were monitored with the Fibox-3 optode system (Presens, Regensburg, Germany). Animals were anesthetized with avertin, transcardially perfused with phosphate-buffered saline (PBS) for 5 mins, followed by 4% paraformaldehyde for 3 mins. Fixed brains were removed and incubated overnight in 70% ethanol, dehydrated in 2 × 30 mins 70% ethanol, 3 × 30 mins 80% ethanol, 3 × 30 mins 90% ethanol, 2 × 30 mins 96% ethanol, 2 × 30 mins 100% ethanol, 2 × 30 mins chloroform, and then kept over night in chloroform. The fixed brains were embedded in paraffin blocks, 5 μm sections were cut and mounted on poly-
Cell Culture
Preparation and maintenance of E18 cortical neurons and P1 mixed whole brain cell cultures isolated from Wistar rats are described elsewhere (Brewer, 1995; Lautenschlager et al, 2000). Cells were plated on glass cover slips in 24-well plates for immunostaining, or in 80 cm2 tissue culture flasks for RNA and protein extraction, and maintained at 37°C and 5% CO2 in a humidified incubator.
Cell Culture Hypoxia
E18 cortical neurons were transferred into 24-well plates containing medium with 60 μg/mL (200 μmol/L) pimonidazole and preincubated for 1 h in the incubator. Then, the cover slips were placed on a rack and dipped into 200 mL of pimonidazole medium in a 500 mL beaker at room temperature (RT) with an oxygen level of 0.8% O2 that was achieved by a bubbling gas mixture of 98.2% N2, 0.9% CO2, and 0.9% O2 at a flow rate of 0.5 L/mins into the medium. Oxygen levels were constantly monitored.
Generation of α-Globin-Specific Antibodies
A total of 15 amino acids (CWGKIGGHGGEYGEE) of the N-terminal rat α-globin sequence (NCBI accession number P01946, positions 14 to 28) were chosen for synthetic peptide generation. Peptide synthesis, immunization of two rabbits, and affinity purification of the antibody with the immobilized synthetic peptide was provided by Biotrend (Cologne, Germany).
Blocking of the Hemoglobin-Specific Antibody
Purified rat Hb was purchased from Sigma (Sigma-Aldrich, St Louis, MI, USA) and diluted 0.01% (w/v) in PBS/1% fetal calf serum, the synthetic peptide used for immunization was diluted 0.0025% (w/v). The antibody was preincubated with either of these dilutions for 1 h and then used for immunostaining.
Immunostaining
Paraffin-embedded brain slides were rehydrated in xylol twice for 10 mins, 100%, 96%, 90%, and 70% ethanol for 5 mins each, and boiled in 1.8 mmol/L citric acid, 8.2 mmol/L sodium citrate buffer for 15 mins in a microwave oven at 700 W. After cooling for 15 mins, slides were rinsed with distilled water and washed twice in PBS. Samples were blocked for 30 mins at RT in Seablock (Pierce, Rockford, IL, USA) and incubated with the primary antibody overnight at 4°C in a humidified chamber using the following concentrations: 1:1,000 rb anti-Hbaa1 (custom antibody; Biotrend, Cologne, Germany); 1:200 ms anti-Ngb (ab37258; Abcam, Cambridge, UK); 1:500 ms anti-MBP (MAB381; Millipore, Billerica, MA, USA), 1:50 ms antipimonidazole (Chemicon Europe, Hampshire, UK), respectively. Slides were washed 3 times for 5 mins in phosphate-buffered saline-Tween 20 and incubated with an appropriate fluorescent secondary antibody for 1 h at RT. Secondary antibodies were anti-rb or anti-ms Cy2-, Cy3-, or Cy5-conjugated IgG (1:400; Jackson ImmunoResearch, West Grove, PA, USA), respectively. After washing twice in phosphate-buffered saline-Tween 20 and once in PBS, nuclei were stained with 46-diamidino-2-phenyl indole (DAPI; 1:5,000; Molecular Probes, Eugene, OR, USA) for 10 mins. Slides were fluoprotected in Mowiol and stored at 4°C in the dark. Tissue culture cover slips were fixed with 4% paraformaldehyde for 30 mins, washed twice in PBS, permeabilized with 0.5% saponin for 10 mins and blocked for 1 h in Seablock. The following steps were identical to those used for the paraffin slides, but PBS/1% fetal calf serum was used instead of phosphate-buffered saline-Tween 20.
Microscopy
Samples were acquired with an Axiovert200M epifluorescence microscope using a × 25 Plan-NEOFLUAR (× 25/0.80 Imm Korr) water immersion objective (both Carl Zeiss, Oberkochen, Germany). The light source used was a USH-102D 100 W mercury lamp (Ushio, Tokyo, Japan) and the filter sets used were: DAPI ex: 350/50 nm, em: 460/50 nm; Cy2 ex: 470/40 nm, em: 525/50 nm; Cy3 ex: 535/50 nm, em: 610/75 nm; Cy5 ex: 628/40 nm em: 692/40 nm. Images were acquired by the AQuest software version 1.1.2002b (TissueGnostics, Vienna, Austria), equipped with a CCD camera (PixelFly qe; PCO, Kelheim, Germany). Images were saved as 8-bit greyscale for each channel.
Shading Correction
A 10% fluorescein isothiocyanate solution was prepared as described elsewhere (Model and Burkhardt, 2001) as shading standard and mounted with Mowiol between a cover slip and a slide. An average of four images at different positions on the slide was calculated and used as ‘blank field’. Image stack intensities were corrected using the formula:
Image normalization
Within each experimental group, the histograms of the pimonidazole and the Hb channels of all stacks were averaged. A correction factor was calculated by dividing the individual histograms by the respective average. The image stacks of each individual animal were normalized by multiplying the pixels of each channel with the corresponding correction factor.
Binary Mask
Normalized channels were transformed into binary masks. A cutoff was set for each channel using the ImageJ multithresholder-filter (pimonidazole: lower=30, upper=250; Hba: lower=50, upper=250), defining signals as pixels within the thresholds and no signal as pixels outside the thresholds. The number of pixels with signals in a 42-image stack was summarized for each channel. To ensure readout from brain tissue only, nontissue (ventricles) and deleted areas, all pixels with a grey value <6 in the normalized stacks were subtracted from the total area. This procedure specified the percentage of Hbα- and pimonidazole-positive area within the tissue of the two hippocampi scanned for each animal.
Hemoglobin Induction by EPO
Recombinant human erythropoietin (ERYPO FS 2000; Janssen-Cilag, Neuss, Germany) was injected i.p. into NMRI mice at a dose of 5,000 IU/kg body weight. Animals were perfused with Hank's balanced salt solution after 6, or 24 h, respectively, and brains were removed. Sham control animals were injected with the vehicle only.
RNA Extraction and Reverse Transcription
Total RNA was extracted from cultured neurons or mouse brains using the RNeasy kit (Qiagen, Hilden, Germany). For reverse transcription, the Omniscript RT kit (Qiagen) and oligo(dT) primers (Promega, Madison, WI, USA) in a 40 μL reaction were used for a template of 2.5 μg RNA.
RT-PCR and Real-Time RT-PCR
One microliter of cDNA was used as template in the DryMix SuperHOTTaq Mastermix (Bioron, Ludwigshafen, Germany) with 3 mmol/L MgCl2, 0.2 mmol/L dNTPs (each), 0.8 μmol/L primers (each), and 2 U per tube Taq polymerase. Rat Hb, Ngb and cytoglobin were amplified in a two-step nested primer PCR reaction (for sequences, see Supplementary Table 1). To ensure amplification from cDNA only, one primer in each step covered an exon/exon junction. RT reaction mixes prepared without reverse transcriptase were run as negative controls. PCR cycles were initial 5 mins at 94°C; 35 cycles of 30 secs denaturation at 94°C, 30 secs annealing at the annealing temperatures specified in Supplementary Table 1, and 30 secs elongation at 72°C; 5 mins final elongation at 72°C. RT-PCR products were separated in 2% agarose/TAE gel electrophoresis. Light cycler amplification of α-globin (primers: forward: 5′-ggtgccctgtctgctctg-3′, reverse: 5′-ggcagcttaacggtacttgg-3′) and β-globin (forward: 5′-ctgactgatgctgagaaggct-3′, reverse: 5′-tccagccaccaccttctgga-3′) was reported previously (Maurer et al, 2002).
Sequencing of the PCR Products
Seventy-five microliters of the PCR product were purified by the NucleoSpin Extract II kit (Machery and Nagel, Düren, Germany) and 100 ng of the purified PCR product were sequenced (Eurofins MWG, Ebersberg, Germany). All primer sequences are listed in Supplementary Table 2.
Quantitative Analysis of Mouse Brain Slides
Coronal sections of mouse brains were cut at stereotactic coordinates interaural 2.34 mm/bregma −1.46 mm (Franklin and Paxinos, 1997). Two uniform regions (3 × 7 tiles) of the hippocampus were scanned per section, resulting in a total of 42 tiles (8.3 mm2) that were combined to an image stack. The exposure times were 6 ms for DAPI (no 12-bit cutoff, gain 0 ×), 209 ms (lower 12-bit cutoff=303, gain 1 ×) for Cy2, and 1,500 ms (lower 12 bit cutoff=400, gain 1 ×) for Cy5. ImageJ was used for further processing of the images (http://rsbweb.nih.gov/ij/).
Hb Quantitation
Hb was quantified by the cyano-Hb method (Zwart et al, 1984).
Two-Dimensional Gel Electrophoresis and Mass Spectrometry
Total brain protein lysates were separated on two-dimensional gels as described (Maurer et al, 2004a). Briefly, brain tissue was homogenized in 2 mL of sample buffer consisting of 40 mmol/L Tris, 7 mol/L urea, 2 mol/L thiourea, 4% (w/v) 3-((3-cholamidopropyl)dimethylammonio)-1-propane sulfonate, 10 mmol/L dithiothreitol, and 1 mmol/L EDTA and centrifuged for 45 mins at 100,000
Peptide mass fingerprinting for the identification of Hb species was performed at the Center for Molecular Medicine (University of Cologne, Germany) as described elsewhere in detail (Feldmann et al, 2005).
Results
Hb is Expressed in Neurons of Rats and Mice
First we isolated neurons from the cortices of 18 days old Wistar rats. The cells were seeded in serum-free medium that was exchanged every 3 to 4 days to rule out the possibility of red blood cell contamination. We detected α- and β-globin mRNAs by RT-PCR in the neuronal cultures up to 2 weeks after seeding. The primers used in the PCR covered exon—exon junctions to guarantee amplification from transcribed mRNA only.
The PCR products were purified and identified by sequencing (Supplementary Table 3). Database comparison of the sequencing results confirmed that the PCR products correspond to α- and β-globin (gi∣17985948∣ref∣NM_033234.1 and gi∣37748074∣gb∣BC059150.1∣).
We derived a peptide antiserum against rat α-globin (Supplementary Figure S1). A sequence similarity is only found in the murine Hbα protein (a mismatch of three amino acids). Other globin sequences share little similarity with the peptide sequence. Only two conserved amino acids are shared with Cygb, three amino acids with Ngb. The strong difference in the amino acid composition between Hbα and Cygb/Ngb shows that these proteins are phylogenetically more distant from Hb than other globins.
In immunofluorescent stainings of adult mouse brains, we detected α-globin (Hbα) expression in neurons of the cerebral cortex, cerebellum, hippocampus, and striatum (Figures 1A–1C). The strongest Hbα signals were observed along the dendrites and the axons of individual neurons and in subcortical fiber tracts. We found coexpression of Hbα with the neuronal marker βIII-tubulin (clone TuJ-1), but not with the astrocytic marker glial fibrillary acidic protein (GFAP) and the oligodendrocytic marker myelin basic protein (MBP) (Figures 1D–1O). TuJ-1 did not stain all Hbα-positive neurons, like the granular cells in the dentate gyrus of the hippocampus. Neuronal expression was also confirmed by immunofluorescent staining of mixed whole-brain cultures (P1), consisting of neurons and glia cells (Supplementary Figure S2).
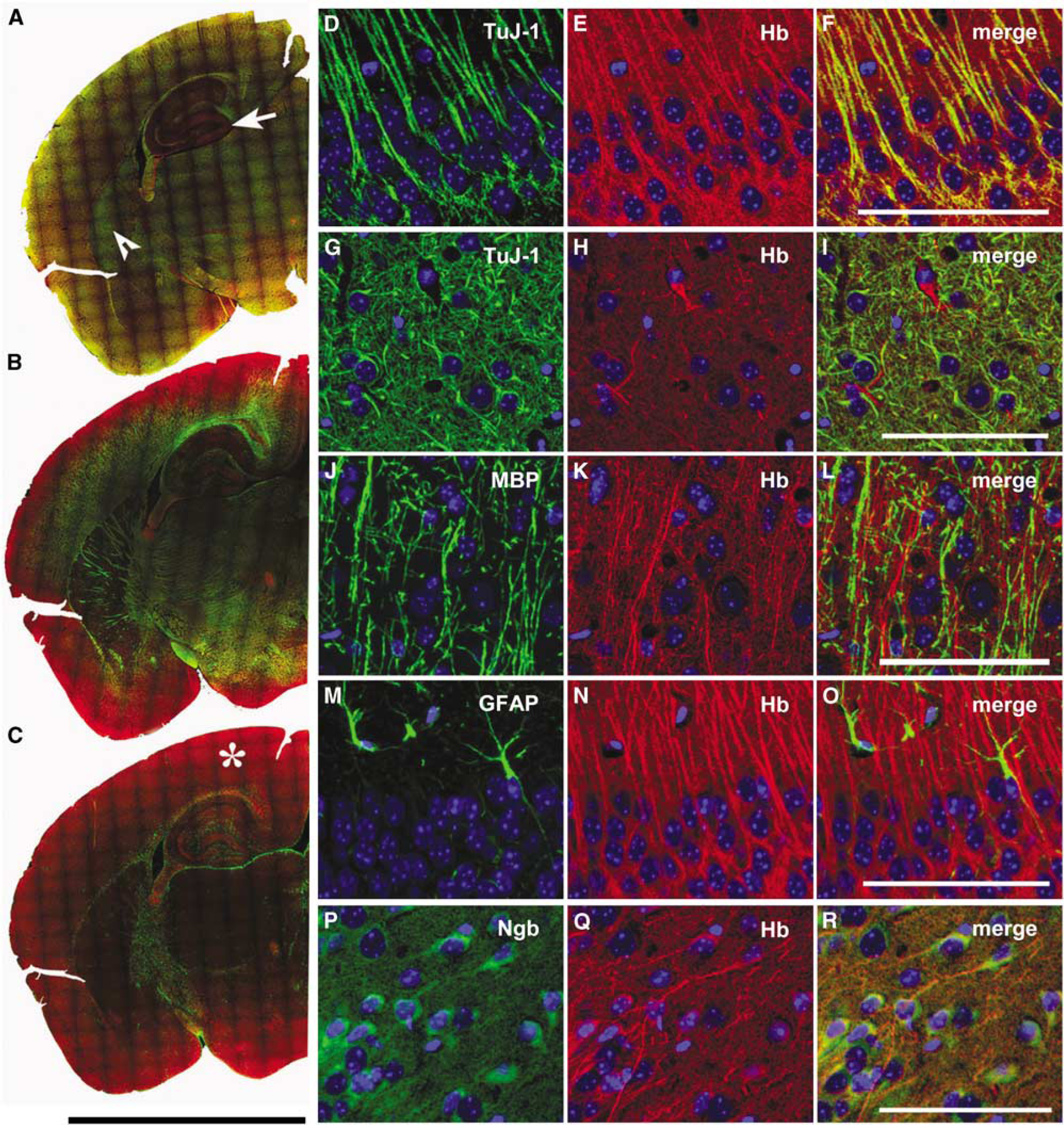
Hemoglobin is expressed in neurons, but not in astrocytes and oligodendrocytes of the mouse brain. Coronal paraffin sections were immunostained and tile scanned. Anti-Hbα was combined with antibodies against different cellular marker proteins. (
Immunostaining also revealed that Hbα expression differed from Ngb expression. Hbα-negative Purkinje cells of the cerebellum expressed Ngb, whereas neurons of the cortex and hippocampus expressed both Hbα and Ngb. The subcellular distribution of Ngb was detected mainly in the perinuclear soma in sharp contrast to the Hbα signal found predominantly in axons and dendrites (Figures 1P–1R; Supplementary Figure S3). A discrete staining pattern was also observed in cultured cortical neurons (Supplementary Figures S4 and S5). The distinct expression pattern both on the cellular and subcellular level suggests that Hbα has a function different from Ngb in neurons.
EPO Increases the Neuronal Expression of Hb in Mice
A lack of oxygen (hypoxia) in the vertebrate organism is counteracted by an upregulation of EPO. This endocrine physiologic response increases the concentration of Hb in the blood, resulting in an enhanced oxygen delivery to the tissue. EPO and the EPO receptor (EPO-R) are also expressed in the brain and induced by hypoxia, mediating neuroprotection (Marti, 2004). Therefore, we asked whether the neuronal Hb expression might also be triggered by EPO.
Initially, we analyzed brain mRNA of the EPO-transgenic mouse line Tg(PDGFB-EPO)321Zbz (Ruschitzka et al, 2000) on a microarray. Compared with wild-type littermates, EPO-transgenic mice had an elevated level of α-globin mRNA (4.76±0.5-fold;
A neuroprotective single dose of human recombinant EPO 5000 IU/kg i.p. also induced Hb expression in the brains of wild-type mice after 24 h on RNA and protein levels. Compared with sham-treated animals, we observed a strong time-dependent induction of α- and β-chain mRNA (1.6- and 1.3-fold at 6 h; 13.7- and 10.8-fold after 24 h, respectively) by quantitative RT-PCR (Figure 2A). Applying the cyano-Hb method, we detected a significant increase of Hb in lysates of perfused brains (Figure 2E). A stronger immunoreactivity in the EPO-treated animals confirmed the increased Hbα expression regionally in the cerebellum and hippocampus (Figure 2B). We performed two-dimensional protein electrophoresis and mass spectrometry to identify the composition of the Hb tetramer. Corroborating the mRNA data, the predominant globin species were α- and β-globin in brain protein lysates (Figures 2F–2G).
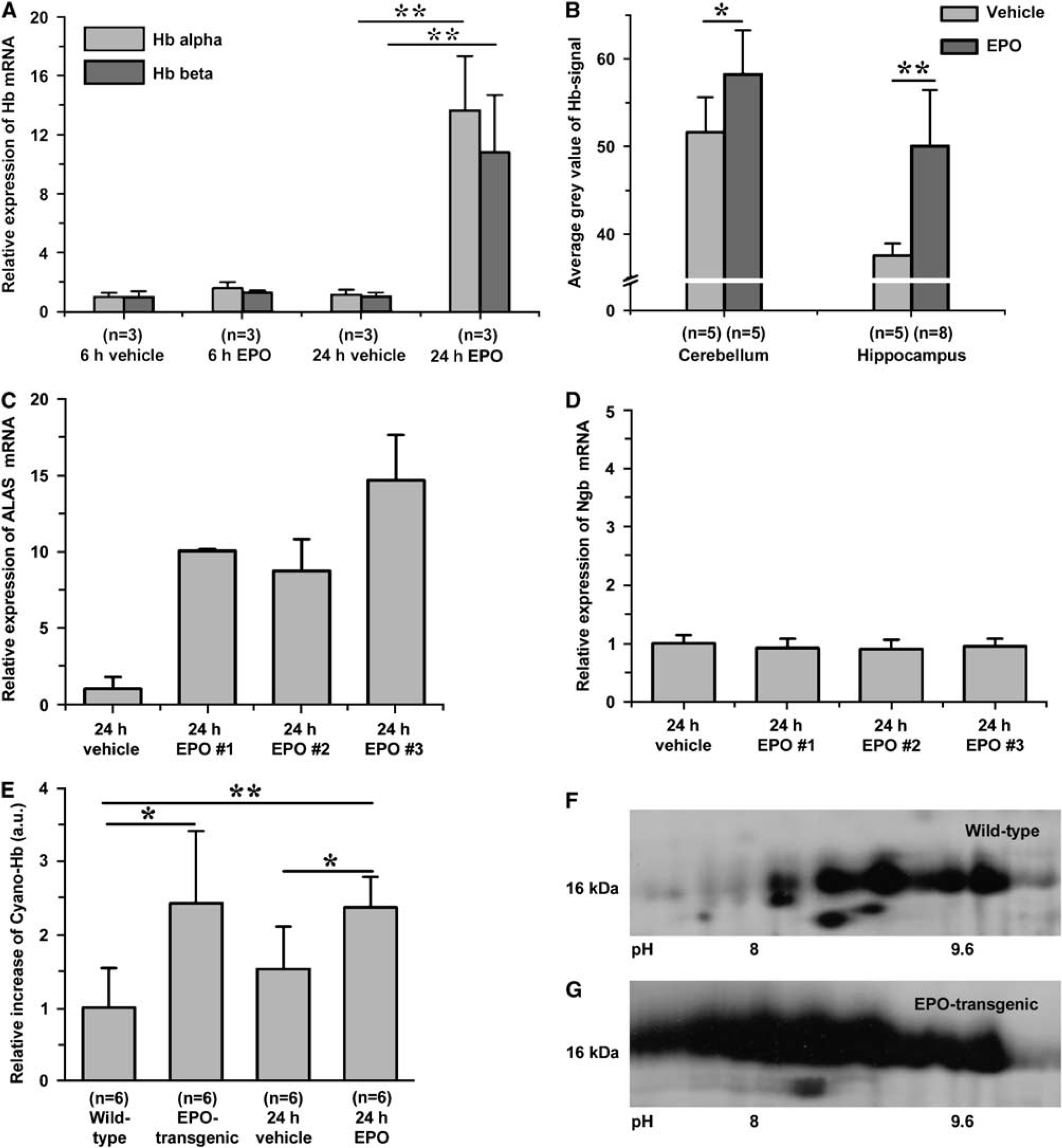
EPO increases hemoglobin expression in the brain after single-dose injection and in EPO-transgenic animals. Perfused brains of EPO-injected and EPO-transgenic mice were used for RNA studies, immunostaining, and protein extraction. (
A functional Hb contains the prosthetic group heme. We therefore tested for transcriptional changes of the key enzyme in heme synthesis, α-aminolevulinate synthase (ALAS) by RT-PCR. Twenty-four hours after EPO injection ALAS expression had increased more than 8.8-fold, compared with controls (Figure 2C). In the same animals, Ngb levels remained unchanged (Figure 2D).
Hbα Expression Correlates to Tissue Oxygen Levels in the Mouse Hippocampus
As EPO production in the brain is stimulated by hypoxia (Sakanaka et al, 1998; Tan et al, 1992) and as EPO increased neuronal Hb in our experiments, we addressed the question of a possible connection between brain tissue oxygenation and Hb expression. We injected wild-type C57BL/6 mice with pimonidazole (Hypoxyprobe-1), a marker substance suitable to detect local oxygen gradients below 10 mm Hg (Arteel et al, 1995; Gross et al, 1995). This threshold is within the physiologic range of brain tissue oxygen levels (Ndubuizu and LaManna, 2007). In hypoxic cell culture experiments, we verified the detection of hypoxic exposure by pimonidazole in neurons (Supplementary Figure S6). With immunofluorescent double staining (anti-Hbα and antipimonidazole), we assessed the distribution of oxygen-deficient regions in the healthy adult mouse brain. The somata of neurons in the CA3 and partly in the CA1 region of the hippocampus, as well as Purkinje cells in the cerebellum displayed high pimonidazole staining and thus had suffered from low pO2 during the time of pimonidazole binding (Figure 3). Interestingly, these cells also displayed weak Hbα signal and were often in direct neighborhood with cells that had a strong Hbα signals but did not have low oxygen levels (Figures 3D–3F). This indicates a reciprocal relationship between Hbα and hypoxia, leading to the assumption that Hbα-positive cells can be regarded as ‘oxygen privileged’.
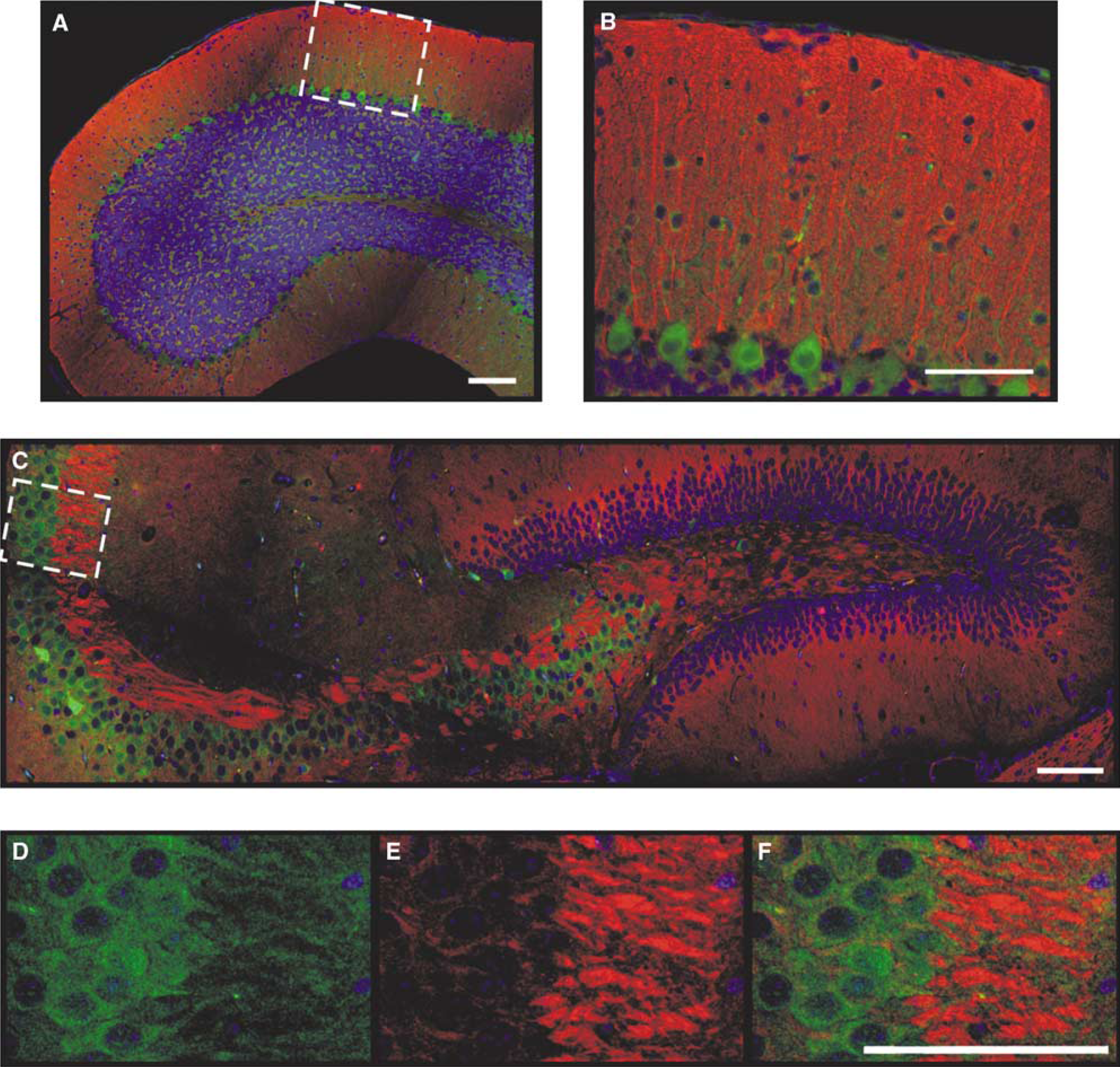
Pimonidazole staining indicates low cellular oxygen levels in areas with a low Hbα expression in the cerebellum and hippocampus. After injection of the hypoxia marker pimonidazole, the adduct was detected by immunostaining. (
EPO Reduces Tissue Hypoxia and Increases Hbα Expression in the Mouse Brain
EPO protects neurons in the hippocampus from hypoxic damage (Sakanaka et al, 1998; Weber et al, 2002). We asked whether a reported neuroprotective dose and treatment scheme of EPO might alter brain hypoxic areas while elevating neuronal Hb levels. C57BL/6 mice were injected with 5000 IU/kg EPO 24 h before the onset of hypoxia (8% O2) for 30 mins. We scanned immunofluorescently stained hippocampal sections and measured the area stained by pimonidazole (Figures 4A–4D). After hypoxia, the stained area had increased from 3.8±2.6 (mean±s.d.)% of the total hippocampus area to 10.5±2.0 (mean±s.d.)% (Figure 4E). The newly appearing areas of pimonidazole binding affected both cells with weak Hbα signals and cells with strong Hbα signals. Hypoxia appeared in focally concentrated regions rather than evenly distributed throughout the hippocampus (Figures 4A–4D). EPO-treated mice showed a significantly reduced pimonidazole-positive area under normoxic (0.3±0.3 (mean±s.d.)%) and hypoxic (6.0±1.5 (mean±s.d.)%) conditions. At the same time, the immunoreactivity for Hbα was elevated in EPO-treated animals (Figure 2B). Thus, EPO induces neuronal Hbα expression and reduces zones of hypoxia both under physiologic and hypoxic conditions. We therefore propose a novel mechanism for EPO action in the brain, counteraction of cellular hypoxia by upregulation of neuronal Hb expression.
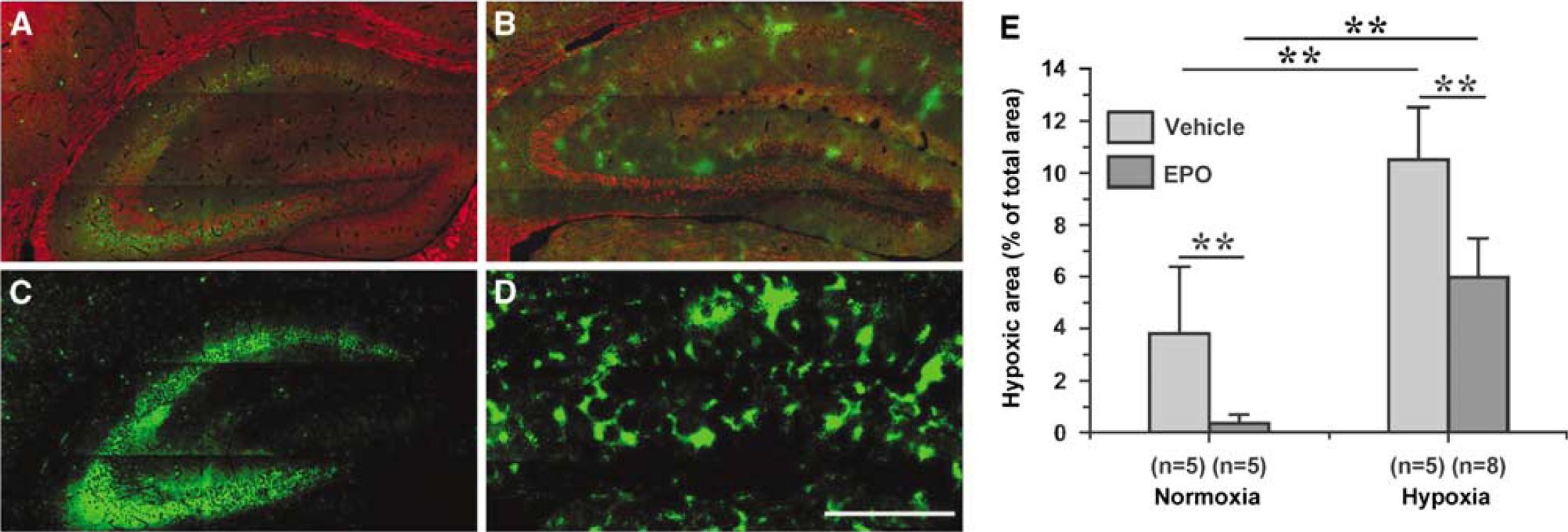
EPO induces hemoglobin expression and diminishes brain hypoxia
Discussion
Hb Expression in Neurons is Specific
We show here that Hb is expressed in rodent neurons. Hb-containing axons of neurons seem to be oxygen privileged in the mouse brain and an increase in neuronal Hb mediated by EPO reduces the overall hypoxic areas in the mouse brain under normoxic and hypoxic conditions.
Other studies reported the expression of Hb in lens cells (Wride et al, 2003), macrophages (Liu et al, 1999), and epithelial cells (Dassen et al, 2008; Newton et al, 2006), which further subverts the notion that Hb expression in vertebrates is restricted to red blood cells, where it is most abundantly found. In the brain, detectable Hb expression has so far been regarded as an artefact resulting from insufficient perfusion (Gauss et al, 1999; Humphries et al, 1976; Tsugita et al, 2000). Short-term expression of Hb in neurons was shown in an earlier study (Ohyagi et al, 1994), but mRNA expression in cell culture could only be detected for a maximum of 15 h after seeding, leaving the possibility of traces of red blood cells in these cultures, even though immunoreactivity was detected up to 63 h after seeding (Ohyagi et al, 1994).
We observed immunohistologic signals in cultured neurons for up to 2 weeks and expression of α- and β-globin mRNA beyond the lifespan of erythrocytes in culture (after 1 week). Neuronal Hb most likely consists of the mature form of Hb (α2β2) as concluded from the mass spectrometry analysis, and from the mRNA expression data. In additional studies of our group we also detected expression of the γ- and ε-globin subunit mRNAs in neuronal cultures.
EPO Regulates Neuronal Hb Expression in the Mouse Brain
EPO as a regulator of neuronal Hb expression provides an argument for a function of neuronal Hb connected to improving the oxygen situation in the brain, supplementing the stimulated production of red blood cells. This hypothesis is supported by the reciprocal relationship between neuronal Hbα expression and oxygen levels that we observed. With the pimonidazole hypoxia marker we detected low levels of oxygen on cellular level in the CA3 neurons of the hippocampus and in the somata of Purkinje cells. An unspecific binding of pimonidazole or the pimonidazole antibody used for immunofluorescent detection to the CA3 cells is unlikely, as the binding characteristics were oxygen dependent. Binding was reduced by EPO and increased by hypoxia treatment. The regions described to express the EPO-R (Digicaylioglu et al, 1995; Morishita et al, 1997; Siren et al, 2001) overlap with sites of high Hbα expression, for example the granular cells of the cerebellum, the pyramidal cells of the cortex, the hippocampus, and fiber tracts. In the brain, globin induction on the mRNA level was induced after 6 h, which precedes the response of erythroid progenitor cells in the bone marrow to single dose of EPO (Bugelski et al, 2008).
Tissue Oxygen Levels and Neuronal Hb Expression
The oxygen affinity of human Hb in the blood is
Globins with different oxygen affinities within the same cell could conduct an intracellular oxygen cascade. In this model, O2 is released from the bloodstream (
Rapid oxygen delivery by neuronal Hb within the neuron could buffer tissue shortages and temporal peaks of oxygen consumption, as already proposed (Offenhauser et al, 2005). The authors observed a linear proportionality between the synaptic activity of climbing fibers of the cerebellum and the disappearance rate of oxygen in the tissue, whereas cerebral blood flow showed a delayed increase to finally meet the oxygen demands. In congruence with this finding and our hypothesis, we found a very strong Hbα signal in the cerebellar cortex and in climbing fibers of the mouse brain. Neuronal Hb as an oxygen buffer could be involved in a phenomenon generally refered to as ‘initial dip’ of the blood oxygen level-dependent effect in brain imaging. Drawing oxygen from the buffer upon activation could explain the initial generation of deoxy-Hb that precedes the increase in blood flow (Buxton, 2001; Mintun et al, 2001).
At the moment, the relationship between Hb and Ngb is still unclear. Their expression patterns are not concordant and Ngb does not react to EPO. It is therefore unlikely that they have a tightly linked function, e.g. in facilitated oxygen transport. In this study, we show that Hbα and Ngb are expressed in the same nerve cells. The question remains whether Hb and Ngb interact or fulfill independent tasks in neurons. The presence of two distinct globins in rodent neurons is supported by the finding that different invertebrate nerve globin lineages of polyphyletic origin exist in invertebrates (Dewilde et al, 2006). They are likely to fulfill independent functions, either acting in an Hb- or Mb-like fashion that involves oxygen transport and storage, or in a Ngb-like fashion.
For a functional role of neuronal Hb, it is essential that its expression can be regulated. We found that the hypoxia-induced growth factor EPO stimulates Hb expression in the mouse brain. Thus, the neuroprotective effect of EPO in ischemia and hypoxia that has been reported in many studies may partly be explained by an enhanced Hb expression in neurons. The finding that Hb is inducible in murine neurons by EPO provides a potential new opportunity for the development of neuroprotective drugs which stabilize or enhance neuronal Hb with regard to oxygen consumption and storage in the brain.
Footnotes
Acknowledgements
We thank M Harlacher and T Lorenz for technical assistance; DC Górecki, University of Portsmouth, UK, for discussing and reading the paper; H Schröck for technical advice; C Ackermann and U Engel, Nikon Imaging Centre, University of Heidelberg, for technical help with confocal microscopy.
Several of the authors (AS, WK, AB, MHM) applied for a patent incorporating some of the findings of this study. Several of the authors (AS, DW, CK, TD, AB, MHM) are employed by the assignee of the above-mentioned patent application.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.