
Abstract
We investigated a gene expression imaging method to examine the level of therapeutic gene expression in the cerebellum. Using a human immunodeficiency virus derived lentivial vector, we expressed the dopamine D2 receptor (D2R) as a reporter protein to mouse cerebellar Purkinje cells. Biodistribution and ex vivo autoradiography studies were performed by giving [125I]5-iodo-7-N-[(1-ethyl-2-pyrrolidinyl)methyl]carboxamide-2,3-dihydrobenzofuran ([125I]IBF) (1.85MBq), as a radioactive D2R ligand, to model mice expressing the D2R with an HA tag (HA-D2R) in the cerebellum. In this study, [125I]IBF was bound to the D2R expressed in the cerebellum of the model mice selectively. Immunostaining was performed to confirm the HA-D2R expression in the cerebellum of the model mice. A significant correlation (r = 0.900, P< 0.001) between areas that expressed HA-D2R by immunostaining and areas in which [125I]IBF accumulated by the ex vivo autoradiograms was found. These results indicated that radioiodinated IBF is useful as a reporter probe to detect D2R reporter gene expression, which can be used for monitoring therapeutic gene expression in the cerebellum.
Introduction
The cerebellum is important in motor coordination and motor learning (Ito, 2002). Damage to the cerebellum causes numerous motor coordination problems, including oculomotor disturbances, postural instability, and gait and limb ataxia. Spinocerebellar ataxias (SCAs), having ataxy as the main symptom, is one of the more serious cerebellar neurodegenerative diseases.
Recently, many genes possessing theraputic potential against the SCAs have been identified (Cummings et al, 1998; Kobayashi et al, 2002; Matsumoto et al, 2004; Miller et al, 2005). To express these genes, using viral vectors, to cerebellar neurons such as Purkinje cells is anticipated to be an effective gene therapy for cerebellar disorders such as SCAs. However, viral vectors have not been used fully in the study of the cerebellum because adenoviral vectors, which are most commonly used for in vivo gene transfer, have limited potential to transduce Purkinje cells (Hashimoto et al, 1996; Terashima et al, 1997). Recently, Torashima et al (2006a) demonstrated a novel method to transduce cerebellar Purkinje cells using human immunodeficiency virus (HIV)-derived lentiviral vectors.
In gene therapy, assessment of levels and area of therapeutic gene expression is of major importance. Therefore, the expression levels and area of any therapeutic gene have to be monitored via imaging of a co-expressed reporter gene with a suitable radiolabeled reporter probe.
In this study, we investigated the usefulness of radioiodinated 5-iodo-7-N-[(1-ethyl-2-pyrrolidinyl)- methyl]carboxamide-2,3-dihydrobenzofuran (IBF) as a reporter probe in monitoring therapeutic gene expression in the cerebellum using mice expressing the recombinant dopamine D2 receptor (D2R) gene as a reporter gene in cerebellar Purkinje cells.
Materials and methods
[125I]-sodium iodide (> 600 GBq/mg) was purchased from Dupont-NEN products. Tributylstannyl-IBF precursor was prepared by Nihon Medi-Physics Co (Nishinomiya, Japan). Other drugs and reagents were purchased from Sigma Chemical Co (St Louis, MO, USA). All procedures for the care and treatment of animals were performed in compliance with the Guidelines for the Care and Use of Laboratory Animals at the Takara-machi Campus of Kanazawa University. In this study, we used a D2R gene with an HA tag (HA-D2R gene) to perform the immunostaining of D2R using a mouse monoclonal antibody against the HA tag (anti-HA Ms Ab; InvivoGen, San Diego, CA, USA).
Virus Preparation
VSV-G pseudotyped lentiviral vectors provided by St Jude Children's Research Hospital (Hanawa et al, 2002) were used in this study. The vectors were designed to express green fluorescent protein or D2R under the control of the murine embryonic stem cell virus (MSCV) promoter (Hawley et al, 1994). The virus vector was produced by cotransfection of human embryonic kidney 293 T cells (2–6×106/a dish) with a mixture of four plasmids (6 µg of pCAGkGP1R, 2 µg of pCAG4RTR2, 2 µg of pCAG-VSV-G, and 10 µg of vector plasmid pCL20c MSCV-green fluorescent protein or -HAD2R) using a calcium phosphate precipitation method as reported previously (Torashima et al, 2006a). Briefly, cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 50 U/mL penicillin G, and 50 µg/mL streptomycin (pH 7.35) at 37°C in a 5% CO2 atmosphere. After the four plasmids were added, the cells were cultured for 48 h, during which they were washed twice with phosphate-buffered saline. The medium samples were filtered through 0.22 µm membranes and centrifuged at 25,000 r.p.m. for 90 mins. The virus samples were finally suspended in 45 µL of phosphate-buffered saline (pH 7.4), frozen in aliquots and stored at −80°C.
Cerebellar Injection
Lentiviral vector inoculation to the mouse cerebellum was performed by the method reported previously (Torashima et al, 2006a). Briefly, a C57Bl/6 mouse under sodium pentobarbital anesthesia (40 mg/kg body weight) was mounted in a stereotactic frame. A burr hole was drilled 5 to 6 mm caudal from the bregma. The tip of a Hamilton syringe attached to a micropump was placed in the molecular layer of the cerebellar vermis (lobule 6), and 6 µL of the virus solution (> 1×1010 TU/mL) was injected at a rate of 200 nL/mins. The mice were used for experiments after breeding for 7 to 14 days.
Radiolabeling
[125I]5-iodo-7-N-[(1-ethyl-2-pyrrolidinyl)methyl]carboxamide-2,3-dihydrobenzofuran ([125I]IBF) was prepared by the iodo-destannylation reaction (Murphy et al, 1990). Briefly, to a solution of sodium [125I]iodide (37 MBq) was added 3 mol/L formic acid (50 µL) followed by a solution of the tributylstannyl precursor (50 µg) in ethanol (50 µL) and H2O2 (3%, 50 µL). The resulting solution was agitated and left for 20 mins. The mixture was quenched with aqueous sodium hydroxide solution (10 N, 30 µL) and then purified by high-performance liquid chromatography using a Zorbax RX-18 column (4.6 mm inner diameter×240 mm length) and acetonitrile/water/ethanolamine (80/20/0.2, v/v/v) as the mobile phase at a flow rate of 0.7 mL/mins. The radiochemical yield was 66.5%. The radiochemical purity was > 99%.
Biodistribution
Three groups of C57Bl/6 mice (n = 4) were anesthetized with ether and administered an intravenous injection of [125I]IBF (111–185 kBq) in 0.2mL saline. At 30, 60, and 120 mins after injection, the animals were killed immediately by decapitation. The organs of interest were dissected, weighed, and the radioactivity levels were measured in a γ scintillation counter (Aloka, ARC-1000, Tokyo). The accumulation of radiotracer was expressed as a percentage of the injected dose per gram of tissue (%ID/g).
In Vitro Autoradiography
To assess binding characteristics of D2R expressed in the cerebellum, the immunostained cerebellar sections were incubated in 50 mmol/L Tris-HCl buffer (pH 7.4) with [125I]IBF (14.8 kBq/mL) in the presence or absence of 10 µmol/L spiperone, which binds to D2R with high affinity. Excess spiperone, which masks D2R expressed in the cerebellum, was used to confirm if the radioactivity was attributed to the specific binding of [125I]IBF for D2R. After 30 mins of incubation at 37°C, the sections were washed three times for 5 mins each time in ice-cold buffer and dried. The sections were apposed to an imaging plate BAS-TR2025 (Fuji Photo Film, Tokyo). After 2 days, the radioactivity distribution was visualized by a Fuji Bio-Imaging Analyzer BAS5000 (Fuji Photo Film).
Ex Vivo Autoradiography
Two weeks after the injection of lentiviral vectors carrying the HA-D2R gene, five model mice expressing HA-D2R and five control mice were injected intravenously with [125I]IBF (1.85 MBq). At 60 mins after injection of the radiotracer, the mice were anesthetized with ether vapor and perfused transcardially with 4% paraformaldehyde plus 2% picric acid in 0.1 mol/L sodium phosphate buffer (pH 7.4). The cerebellum was removed and postfixed overnight at 4°C followed by cryoprotection in 30% sucrose. The cerebellum was cut into 50 µm sagittal sections using a cryostat microtome and consecutive sections were subjected to ex vivo autoradiograms or immunostaining. To perform ex vivo autoradiography, the sections were apposed to an imaging plate BAS-TR2025 (Fuji Photo Film). After 2 days, the radioactivity distribution was visualized by a Fuji Bio-Imaging Analyzer BAS5000 (Fuji Photo Film).
Immunohistochemistry and Acquisition of Brain Images
Other consecutive cerebellar sections used with ex vivo autoradiography were subjected to immunostaining with a mouse monoclonal antibody against the HA tag (anti-HA Ms Ab; InvivoGen). Immunostaining of the cerebellar sections, which were used in in vitro autoradiography, was performed before in vitro autoradiography. The primary antibodies against HA tag were used at dilutions of 1:500. Sections were incubated overnight at room temperature, rinsed twice in phosphate-buffered saline, and then incubated with Alexa fluoro-488-conjugated secondary antibodies (1:1000; Invitrogen, Paisley, UK) for 2 h at room temperature. Immunofluorescence was evaluated by a confocal laser-scanning microscope (LSM 5 Pascal; Zeiss, Oberkochen, Germany).
Correlation Between HA-D2R Expression and [125I]IBF Accumulation in the Cerebellum
Three model mice-expressing HA-D2R were used to assess the correlation between [125I]IBF accumulation and HAD2R expression in the cerebellum. Ten points of regions of interest (ROI) were drawn on cerebellum sections of autoradiographic and immunostaining images. [125I]IBF accumulation was expressed as photo-stimulated luminescence per mm2. HA-D2R expression was expressed as intensity of fluorescence. Photo-stimulated luminescence per mm2 in cerebellum regions was determined by analyzing 10 ROIs on the autoradiographic images using Multi Gauge V3.0 software (Fuji Photo Film). Intensity of fluorescence in cerebellum regions were determined by analyzing 10 ROIs on the immunostained images using Images 1.37v software (National Institutes of Health, MD, USA). GraphPad Prism 4.00 software was used for statistical analysis.
Results
Table 1 demostrates the biodistribution of [125I]IBF in mice. [125I]IBF demonstrated higher accumulation in the striatum with the high-density D2R than the cerebellum as nonspecific binding. The [125I]IBF accumulation ratio of striatum-to-cerebellum ratio were 14.2 ± 2.80, 24.9 ± 3.50, and 48.5 ± 7.90 at 30, 60, and 120 mins after injection, respectively. Thus, [125I]IBF demonstrated a significantly high accumulation striatum-to-cerebellum ratio. [125I]IBF demonstrated low accumulation in blood.
Human embryonic kidney 293 T cells were infected with lentiviral vectors carrying the HA-D2R gene (Figure 1A). The cells were immunostained with an anti-HA antibody 2 days after the injection, demonstrating the efficient expression of D2R (Figure 1B). Expression of D2R protein was confirmed by western blot analysis (Figure 1C).
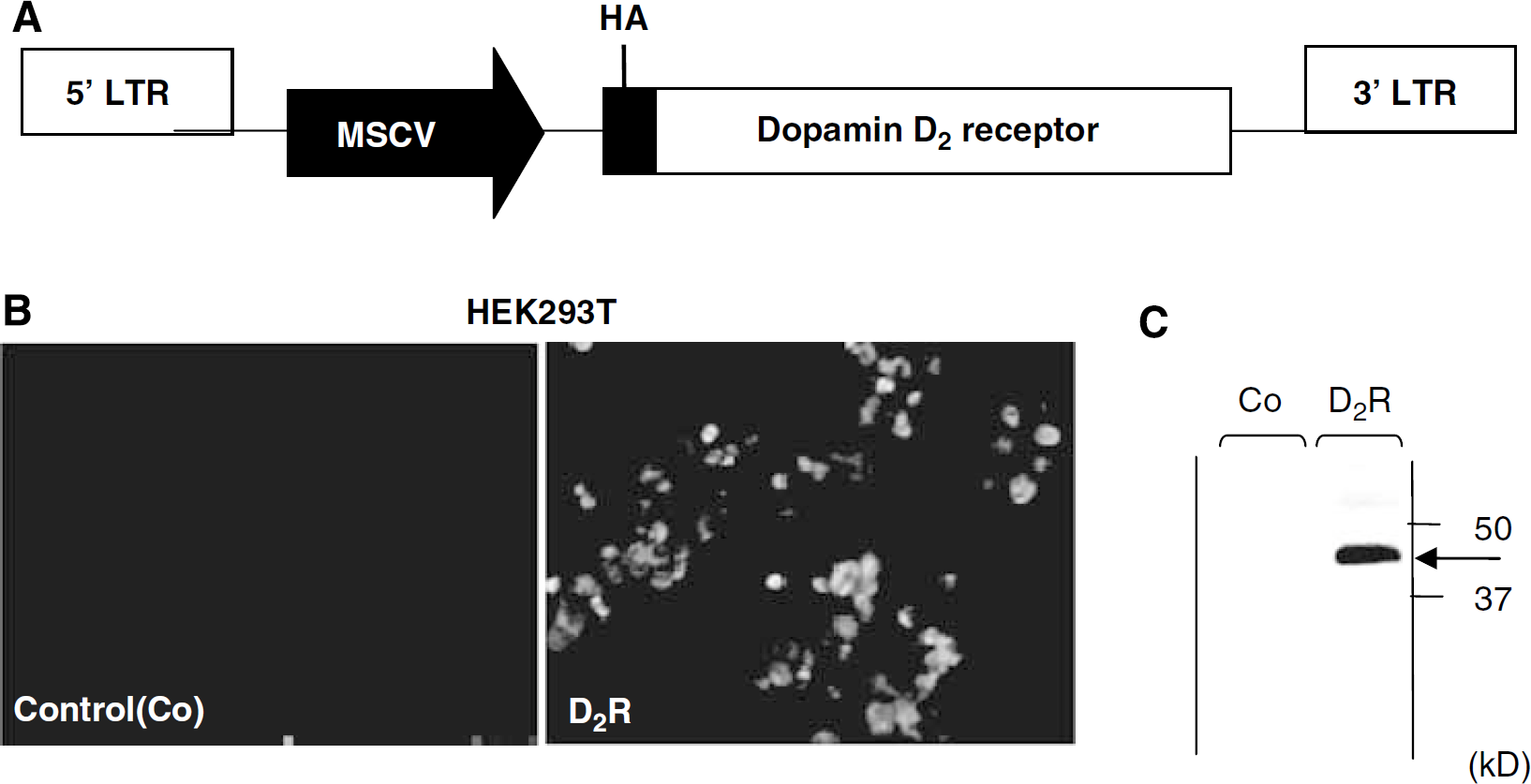
Expression of the HA-D2R gene by lentiviral vectors. (
Biodistribution of [125I]IBF in mice
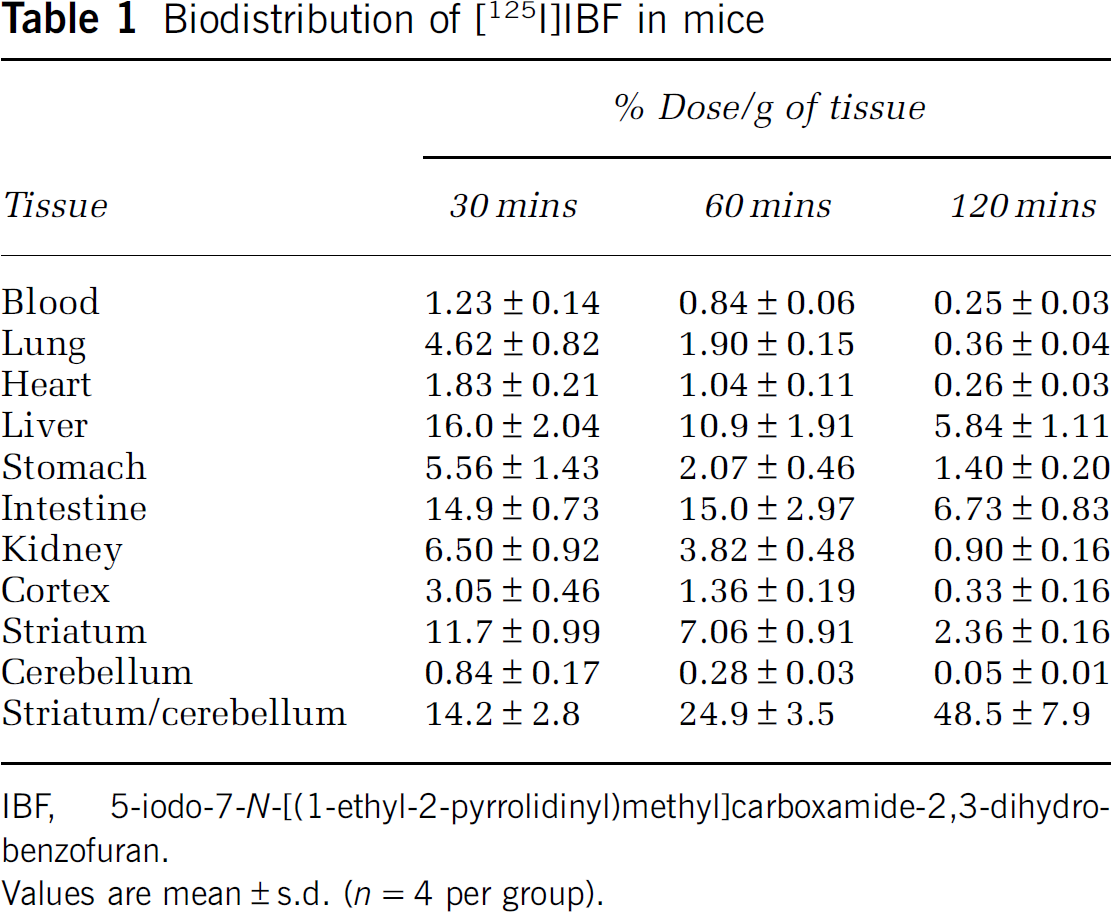
IBF, 5-iodo-7-N-[(1-ethyl-2-pyrrolidinyl)methyl]carboxamide-2,3-dihydrobenzofuran.
Values are mean±s.d. (n = 4 per group).
The mice received injection of lentiviral vectors expressing HA-D2R. They were used for the analysis 7 days after the injection.
Figure 2 demonstrates in vitro [125I]IBF autoradiogram and the immunostaining images in a sagittal cerebellar section of the model mice expressing HAD2R. Both images (Figure 2A and 2C) visualized the area in which HA-D2R was expressed. The specific high density in the cerebellum in in vitro [125I]IBF autoradiograms was dramatically reduced by the administration of spiperone such as the dopamine D2R ligand (Figure 2B).
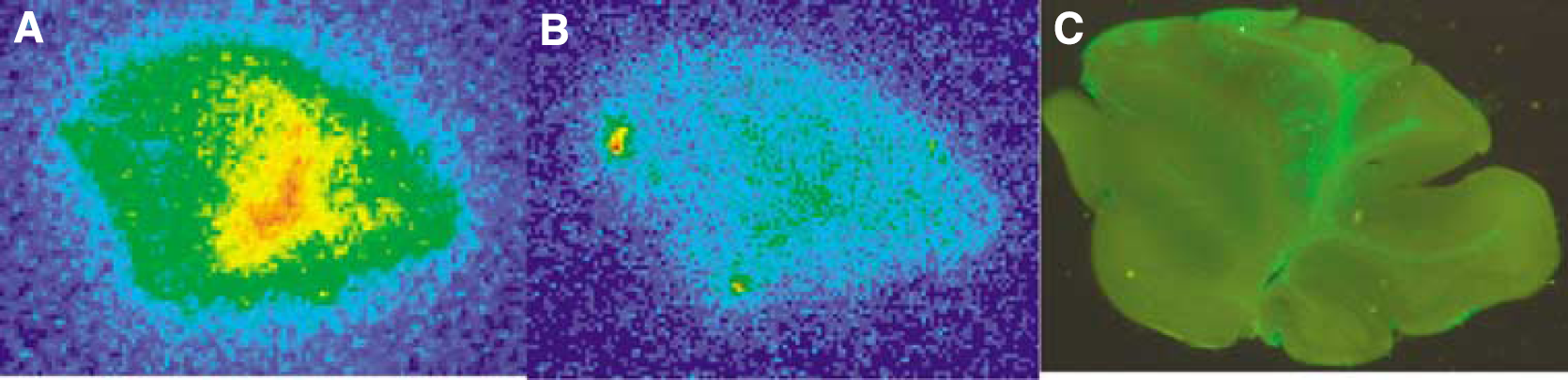
In vitro [125I]IBF autoradiograms of a sagittal section of lentivirus-treated cerebella in the absence (
In ex vivo autoradiograms of the sagittal cerebellar sections, [125I]IBF demonstrated high accumulation in a large part of the cerebellum of the model mice expressing HA-D2R (Figure 3B–3E). In contrast, [125I]IBF demonstrated no significant accumulation in the cerebellum of the control mice (Figure 3A and 3I). Immunohistochemical analysis with an anti-HA antibody demonstrated that HA-D2R was efficiently and selectively expressed in the cerebellum of the model mice (Figure 3F–3H), whereas it was not expressed in the cerebellum of the control mice (Figure 3J). Magnified images demonstrate that Purkinje cells were effectively transduced by lentiviral vectors and HA-D2R was detected in the cell bodies and dendritic arbors of Purkinje cells (Figure 3K). Ten points of ROIs were drawn on cerebellar sections of the autoradiogram (Figure 4E’) and the immunostained images (Figure 4H’) to assess the correlation between [125I]IBF accumulation and HA-D2R expression in the cerebellum. The correlation between [125I]IBF accumulation and HA-D2R expression in the cerebellum is shown in Figure 4. The accumulated area of [125I]IBF on the cerebellar section was well correlated with the area of HA-D2R expression (r = 0.900 ± 0.038 (P < 0.001) (n = 3)).
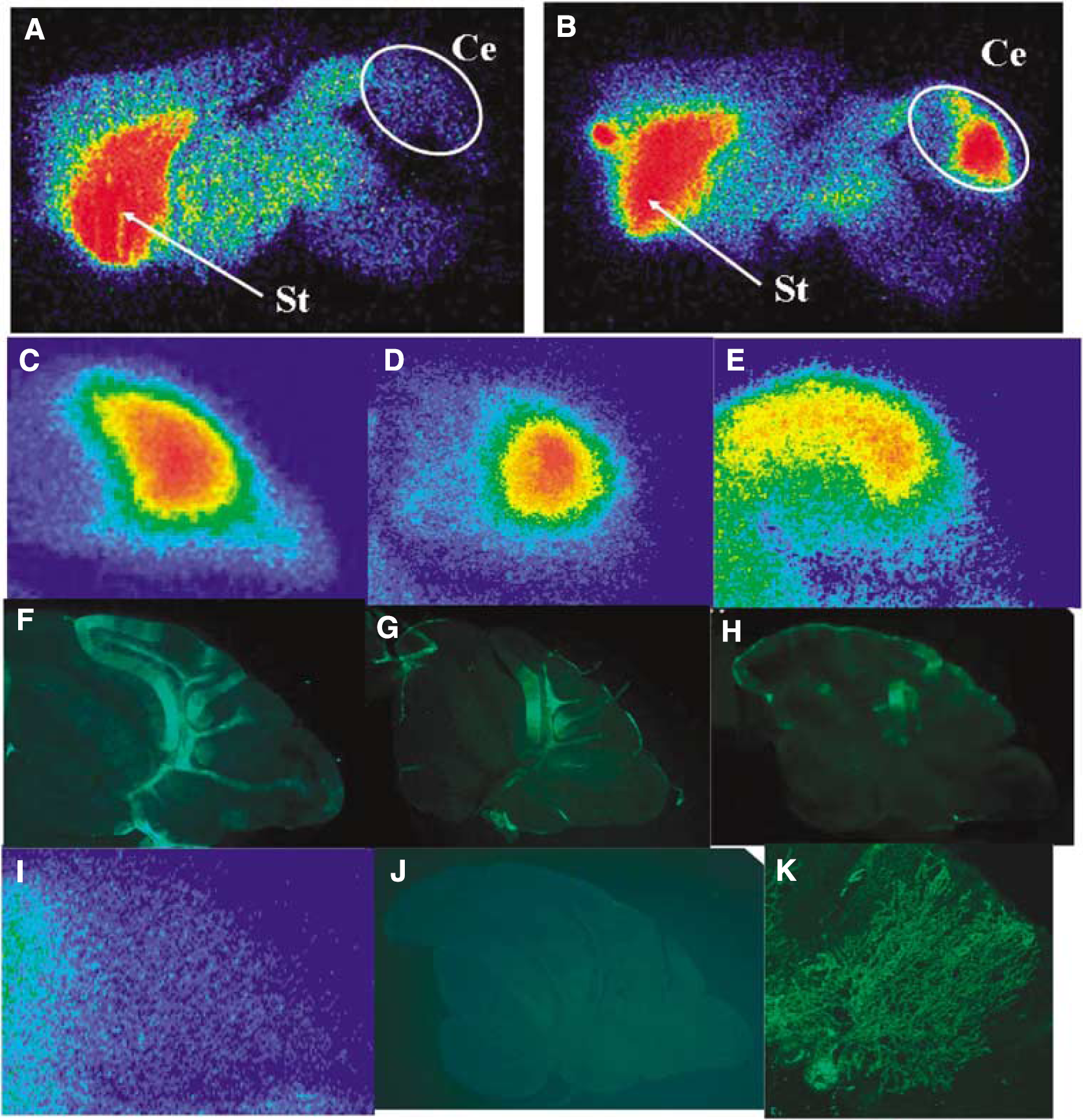
Ex vivo autoradiograms of [125I]IBF in sagittal sections of brains of non-injected control (
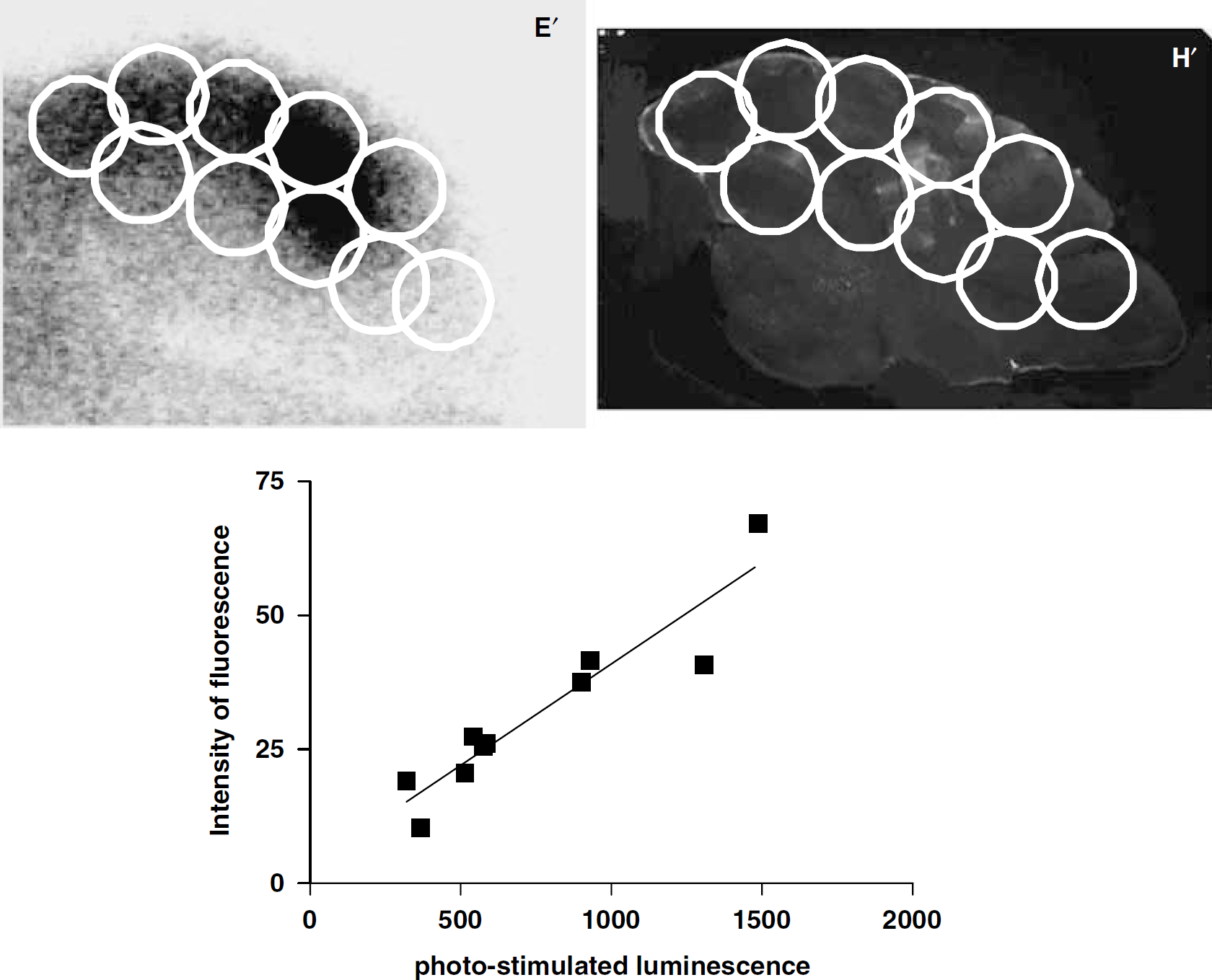
Correlation between [125I]IBF accumulation and HA-D2R expression in the cerebellum. Ten points of regions of interest (ROI) were drawn on cerebellum sections of the autoradiographic image (
Discussion
Human immunodeficiency virus derived lentivial vectors have been increasingly used for gene transfer into neurons. However, there have been no reports testing their potential to transduce Purkinje cells. Recently, Torashima et al (2006a) revealed that lentiviral vectors injected into the cerebellar cortex efficiently transduced Purkinje cells without significantly influencing their viability or synaptic functions. However, lentivector-mediated green fluorescent protein expression in the mouse cerebellum is influenced by some factors such as pH of the culture medium (Torashima et al, 2006b), the lot quality of the serum supplemented in the culture medium (Torashima et al, 2007), and the depth of the viral injection site. Monitoring of the expression of a therapeutic gene in the cerebellum is important in the case of gene therapy for serious cerebellar neurodegenerative diseases. Imaging of the linked reporter gene with a suitable radiolabeled reporter probe is thought to be a useful method to monitor the gene expression. In this study, we selected D2R gene as the reporter gene for the following two reasons: (1) D2R is absent in the cerebellum, which was the target organ. (2) Radioiodinated IBF has already been developed as an excellent D2R radioligand (Buck et al, 1995; Ichise et al, 1997).
The biodistribution study demonstrated that [125I]IBF was superior as a radiolabeled D2R reporter gene probe in comparison with [125I]iodobenzamide reported by Kung et al (1988) because of its much higher ratio of specific binding to nonspecific binding (Table 1). The rapid clearance of [125I]IBF from the mouse body results in low radiation exposure owing to [125I]IBF. The D2R with an HA tag, as well as green fluorescent protein, was expressed in human embryonic kidney 293 T cells transfected with HIV-derived lentivial vectors carrying the HA-D2R gene. Immunostaining with an HA antibody demonstrated that most of the HA-D2R-expressing cells were localized in the Purkinje cell bodies through the dendritic arbors to the spines in the cerebellum of the model mice expressing HA-D2R. Furthermore, [125I]IBF, a selective D2R ligand, bound to HA-D2R expressed in the cerebellum and the binding area of [125I]IBF was significantly correlated with the area of HA-D2R expressed in the cerebellum of the model mice in vivo (Figures 3 and 4) (r = 0.900, P < 0.001), which demonstrated that transduced HA-D2R, a membrane receptor in the cerebellum, maintained the function of the binding to D2R ligands; thus, [125I]IBF is a suitable radio D2R ligand for imaging of transduced HA-D2R in the cerebellum. In in vitro autoradiography, [125I]IBF accumulation was not much matched with HA-D2R expression in the cerebellum of the model mice because the immunostained cerebellar sections might have already been damaged before in vitro autoradiography was performed. However, the decrease in [125I]IBF accumulation in the cerebellum of the model mice expressing HA-D2R in the presence of 10 µmol/L spiperone indicated that [125I]IBF selectively bound to HA-D2R expressed in the cerebellum.
It is usually necessary to develop indirect imaging strategies by using a reporter gene in combination with a therapeutic gene. Several approaches may be useful for indirect imaging of a therapeutic gene in the cerebellum: (a) a bicistronic approach to express two different genes from the same vector construct inserts a linker such as an internal ribosomal entry site sequence between two different genes; (b) a bidirectional promoter approach expresses two different genes from a bidirectional promoter within a single vector; and (c) a double promotor approach expresses two different genes from distinct promoters within a single vector.
The D2R gene has been reported to be useful as a reporter gene for monitoring the expression of a therapeutic gene (MacLaren et al, 1999). These approaches using two genes including a D2R gene have been reported to demonstrate a high correlation of the relative levels of expression between the two genes (Yu et al, 2000; Yaghoubi et al, 2001; Sun et al, 2001). However, the levels of the expression of two different genes, a therapeutic gene and a reporter gene, depend on the size and type of each gene when using these approaches. In the near future, when a therapeutic gene for serious cerebellar neurodegenerative diseases such as SCAs is developed, we will need to investigate the best approach for indirect quantitative imaging by using a D2R gene as the reporter gene in combination with a therapeutic gene.
A potential problem of receptor-based reporter genes such as the D2R gene is that ectopic receptor gene expression could affect the target cell physiology. For example, expression of ligand-binding D2Rs could alter the signaling pathways of target cells and lead to a reduction in the level of cyclic adenosine monophosphate. However, this problem was resolved with replacement amino acid 80 from Asp to Ala in the D2R (Neve et al, 1991; Cox et al, 1992; Woodward et al, 1996). The mutant protein lacks the ability to be regulated by cyclic adenosine monophosphate, but retains affinity for the D2R ligand (Liang et al, 2001).
In conclusion, the D2R gene as a reporter gene was expressed to Purkinje cells in the cerebellum using HIV-derived lentiviral vectors and radioiodinated IBF was found to be useful as a reporter probe of D2R reporter gene expression in the monitoring of therapeutic gene expression in the cerebellum. Further studies will be needed to develop indirect imaging strategies by using a mutant D2R reporter gene such as amino acid 80 from Asp to Ala in the D2R with radioiodinated IBF in combination with a therapeutic gene for serious cerebellar neurodegenerative diseases such as SCAs.