
Abstract
Gender differences in lipid metabolism are poorly understood and difficult to study using conventional approaches. Magnetic resonance spectroscopy (MRS) permits non-invasive investigation of lipid metabolism. We employed novel two-dimensional MRS techniques to quantify intramyocellular (IMCL) and extramyocellular (EMCL) lipid compartments and their degree of unsaturation in normal weight adult male and female subjects. Using muscle creatine (Cr) for normalization a statistically significant (p < 0.05) increase in IMCL/Cr (7.8 ± 1.6) and EMCL/Cr (22.5 ± 3.6) for female subjects was observed (n = 8), as compared to IMCL/Cr (5.9 ± 1.7) and EMCL/Cr (18.4 ± 2.64) for male subjects. The degree of unsaturation within IMCL and EMCL was lower in female subjects, 1.3 ± 0.075 and 1.04 ± 0.06, respectively, as compared to that observed in males (n = 8), 1.5 ± 0.08 and 1.12 ± 0.03, respectively (p < 0.05 male vs female for both comparisons). We conclude that certain salient gender differences in lipid metabolism can be assessed noninvasively by advanced MRS approaches.
Keywords
Introduction
There is a great interest in the potential physiologic differences between males and females that may affect the prevention, diagnosis and treatment of obesity and diabetes. Although males and females are both susceptible to obesity, the incidence and health consequences differ between the sexes (Power and Schulkin, 2008), as do the patterns of fat deposition, fat mobilization, utilization of fat as a metabolic fuel, and the consequences of both excess and insufficient fat stores (Power and Schulkin, 2008). Non-insulin dependent diabetes mellitus (NIDDM) and obesity are associated disorders and are increasing in incidence worldwide (Smyth and Heron, 2006). Insulin resistance, leading to impaired ability of insulin to appropriately regulate glucose and lipid metabolism, is associated with these disorders and other defects of lipid metabolism. The etiology of insulin resistance is still not completely understood, but it is now clear that a strong association exists between insulin resistance and excess lipid accumulation in nonadipose tissues, particularly muscle and liver.
Previous studies have shown that intramyocellular lipid (IMCL) in skeletal muscle is increased with obesity and in NIDDM (Dobbins et al. 2001; Goodpaster et al. 2000; Kelley et al. 1999). The correlation between the magnitude of the IMCL pool as determined by MRS studies, and insulin resistance, diabetes, and disorders of lipid metabolism, has been previously demonstrated (Boesch, 2007; Boesch et al. 2006). Nevertheless, quantification of IMCL and EMCL by MRS remains highly problematic due to the overlapping resonances from both of these lipid pools (Steidle et al. 2002). The ability to distinguish IMCL from EMCL is based on their different bulk magnetic susceptibility effects due to their geometric arrangements within muscle. This leads to a spectroscopic frequency separation between the two pools; this separation is 0.15 parts per million (ppm) in the soleus and 0.2 ppm in the tibialis muscle as validated in vivo (Szczepaniak et al. 2002). At a clinical field strength of 1.5 Tesla, these values are 9.6 Hz and 12.8 Hz respectively, while at 3 Tesla the frequency separations are twofold greater (Steidle et al. 2002). This permits the IMCL and EMCL signals to be individually identified in MRS, but is not adequate for complete separation and accurate quantification.
The degree of lipid unsaturation within the IMCL and EMCL pools is also of substantial clinical importance (The Report of the British Nutrition Foundation's Task Force, 1993; Vessby et al. 2002; Warensjo et al. 2006; Warensjo et al. 2005). In addition, differences between the lipid profiles of males and females may have clinical implications. In women, high plasma triglyceride concentrations as compared to men are an independent predictor of cardiovascular risk, and more strongly predictive than total or LDL cholesterol (Cullen, 2000; Meagher, 2004). Treatment of hypertriglyceridemia may, therefore, be of greater importance in women than in men. Also, differences in hepatic handling of fatty acids between men and women are likely responsible for the greater susceptibility of women to fatty liver disease (McCullough, 2002). In addition, the association of several gene polymorphisms with risk factors for disease and substrate kinetics is often sex specific (Galluzzi et al. 2001; Talmud et al. 2001; Halsall et al. 2000). Therefore, aside from delineation of basic metabolism, improved understanding of the influence of gender on lipid metabolism may be useful for the development of novel approaches to the treatment and prevention of disease.
Accordingly, our laboratory has developed novel MRS approaches that permit improved spectral resolution for detecting both saturated and unsaturated components of lipids (Velan et al. 2007a; Velan et al. 2007b). The purpose of the present study was to assess gender differences in IMCL and EMCL, along with the degree of unsaturation within these lipid pools. In this study we have applied these novel MRS techniques to the skeletal muscle of normal weight adult males and females.
Materials and Methods
Subjects
A total of 16 normal weight subjects (8 males and 8 premenopausal females; BMI range 20–24.5 kg/m2, matched between males and females; age range 25 ± 5 years) were studied. All subjects were healthy and without any diagnosed or suspected metabolic disorders, and gave informed consent. The experimental protocol was approved by the local institutional review board. Subjects were not participants in high levels of exercise training, and were instructed to avoid fatty foods for at least three days prior to data acquisition.
MRI and MRS Experiments
All experiments were performed using a 3T MRI/MRS Excite HD scanner (GE Healthcare Technologies, Waukesha, WI) with self-shielded gradients (40 mT/m). At the time of MRS data acquisition, subjects were positioned supine feet first within the magnet, with their right calf placed inside a vendor-supplied quadrature transmit/receive coil. All data were acquired from the soleus muscle. The soleus muscle was selected for this study since its location permitted optimal Bo homogeneity in the volume of interest (VOI) and it has a substantially larger volume compared to other muscle compartments. In order to standardize the position and alignment of the subject's leg, a point 1/3 of the distance from the medial femoral condyle to the medial malleolus was marked and used for MRS data acquisition.
Gradient echo scout images were acquired in the axial, coronal and sagittal planes with 5 mm slice thickness, in-plane spatial resolution of 976 µm x 976 µm, repetition time (TR) = 14.7 ms, number of excitations (NEX) = 1, and echo time (TE) = 2.25 ms. Using these images to localize the target volume, MRS data was then collected from a 3 x 3 x 3 cm3 (27 ml) voxel within the soleus using two protocols as described below.
Localized 2D correlation spectroscopy (L-COSY) (Velan et al. 2007a; Thomas et al. 2001), in which cross-peaks are formed from indirect spin-spin couplings, was performed on all subjects. Parameters included a minimal TE of 30 ms and 64 incremented time steps t1, with increment Δt1 = 0.8 ms, resulting in a spectral bandwidth of 1250 Hz along the second, indirectly detected, spectral dimension. The signals for each value of t1 were averaged over 8 scans, resulting in a total acquisition time of approximately 17 minutes. The directly detected dimension (t2) had a bandwidth of 5000 Hz with 2048 complex points sampled.
Spectral Processing
Post processing was performed using FELIX software (Accelrys Inc, San Diego, CA). Prior to Fourier transformation, the time domain data matrix was apodized using shifted sine squared filters in the t1 and t2 dimensions. The length of the windows was adjusted so that the filters reached zero at the last experimental data point in both dimensions. All data sets were linear predicted from 50 t1 points to 100 t1 points along the F1 dimension and zero filled to 256 points. The diagonal and cross peaks were assigned and the volume integrals were quantified including corrections for T1 relaxation as described previously (Velan et al. 2007a). The diagonal peak volumes from (CH2)n protons within the IMCL [1.2,1.2 ppm] and EMCL [1.35,1.35 ppm], and the creatine signal at 3.03 ppm, were estimated. The volumes of the cross peaks between olefinic (-CH=CH-) and allylic methylene protons (CH2CH=CH) within IMCL and EMCL at [5.3,2.0 ppm] and [5.45,2.15 ppm], respectively, reflect the size of the monounsaturated fatty acid component of the two lipid pools. Similarly, the cross peak volumes between olefinic and diallylic methylene protons (-CH=CH-CH2-CH=CH-) within IMCL and EMCL at [5.3,2.7 ppm] and [5.45,2.85 ppm], respectively, reflect polyunsaturated fatty acids. Ratios of these cross peak volumes define the degree of unsaturation (Velan et al. 2007a; Velan et al. 2007b).
Statistical Analysis
ANOVA analysis was used to compare the IMCL/Cr, EMCL/Cr and degree of unsaturation among normal weight male and female subjects. The post-hoc Bonferroni test was employed to test for pairwise differences. Data are presented as mean ± SD, and statistical significance was assigned at the P < 0.05 level.
Results
Figure 1 shows a typical 2D L-COSY spectrum obtained from a normal weight male subject. Resonance assignments were based on our earlier work (Velan et al. 2007a). In addition to the CH3 and (CH2)n groups of IMCL and EMCL, olefinic, allylic methylene, and diallylic methylene groups from the IMCL and EMCL lipid pools can be identified by the cross peaks C1, C2, C3 and C4. The glycerol backbone protons (cross peaks labeled as C5) and the imidazole protons of carnosine are also evident (cross peaks labeled as C6). The cross peaks labeled C1 and C3 arise from the indirect spin-spin coupling between olefinic (-CH=CH-) and allylic methylene protons CH2CH=CH of IMCL and EMCL, respectively. They appear if the methylene protons are adjacent to only one unsaturated site. Cross peaks C2 and C4 arise from the indirect spin-spin coupling between olefinic (-CH=CH-) and diallylic methylene protons (-CH=CH-CH2-CH=CH-) of IMCL and EMCL, respectively. These peaks appear if there are atleast two olefinic groups that appear symmetric to methylene protons. The volumes of C2 and C4 increase as the number of unsaturated sites increases, so that the degree of unsaturation can be estimated from the volume ratios C2/C1 and C4/C3 within IMCL and EMCL, respectively (Velan et al. 2007a). The cross peaks labeled C5 are due to the J coupling between CH2 and CH groups of the glycerol backbone protons, while cross peaks C6 are due to the residual dipolar coupling between CH groups from the imidazole ring protons of carnosine (Velan et al. 2007a; Velan et al. 2007b). The diagonal resonance frequencies (denoted [F1, F2]) from the n-methylene protons resonating at [1.2,1.2] ppm for IMCL and at [1.35,1.35] ppm for EMCL, and the signal of the methylene group of creatine at [3.03,3.03] ppm, were used to calculate the ratios IMCL/Cr and EMCL/Cr.
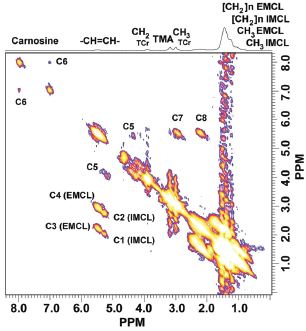
Localized 2D L-COSY spectrum from the soleus muscle of a normal weight subject acquired with minimal TE of 30ms, 8 averages for each t1, 50 t1 increments, and voxel size = 27ml. 2048 complex points were acquired with a spectral width of 5 kHz.
Figures 2 and 3 show mean IMCL/Cr, EMCL/Cr and degree of unsaturation for male and female subjects. Average IMCL/Cr and corresponding coefficients of variation (CV) in the male subjects were 5.9 ± 1.7 (28.8%) and 7.8 ± 1.6 (20.5%) for the normal weight male and female subjects, respectively (Fig. 2); this difference was statistically significant.
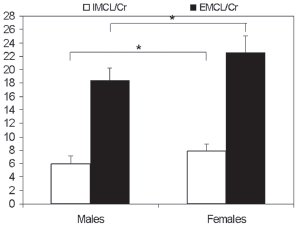
Mean IMCL/Cr and EMCL/Cr for normal weight males and females (n = 8 per group). The vertical axis represents the mean values of IMCL/Cr and EMCL/Cr ratios from the two groups of subjects.
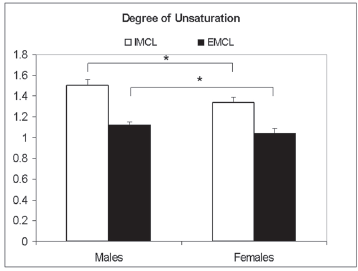
Mean degree of unsaturation for males and females (n = 8 per group). The vertical axis represents the mean degree of unsaturation within IMCL and EMCL from the two groups of subjects.
Corresponding EMCL/Cr values were 18.4 ± 2.64 (14.3%) and 22.5 ± 3.6 (16%), (Fig. 2), which again was a statistically significant difference.
The average lipid unsaturation in the IMCL pool and the corresponding CV in the male and female subjects were 1.51 ± 0.08 (5.2%) and 1.3 ± 0.075 (5.7%), respectively (Fig. 3). The gender difference was statistically significant here as well.
Corresponding values for the degree of unsaturation in the EMCL pool were 1.12 ± 0.037 (3.3%) and 1.04 ± 0.06 (5.7%) and again, the difference was statistically significant.
Discussion
Excess accumulation of lipid in tissue can occur as a result of increased fatty acid uptake, a decreased rate of fatty acid utilization, or a combination of these factors. There is clear evidence of gender-related differences in whole-body lipid metabolism (Power and Schulkin, 2008; Blaak, 2001; Williams, 2004), with marked differences seen in the amount and distribution of excess body fat between men and women. As an example, in the basal condition, women are physiologically adapted to store fat to a greater extent than are men (Power and Schulkin, 2008; Blaak, 2001; Williams, 2004). Such differences arise from metabolic and hormonal differences between males and females, and also contribute to differences in their health risks attributable to obesity. However, the mechanisms responsible for these gender-related effects are largely unknown.
Adipocytes take up circulating fat via lipoprotein lipase (LPL) as well as through direct uptake and storage of circulating free fatty acids (Santosa and Jensen, 2008). The transport of fat in the blood stream is approximately twice as fast in women as men (Knopp et al. 2005). Further, obesity and diabetes are associated with greater lipoprotein abnormalities in women as compared to men, with a greater increment in cardiovascular disease risk in women being linked to these abnormalities. A greater change in triglyceride level and a lesser change in low-density lipoprotein level is observed in women as compared to men with high-carbohydrate or high-fat feeding, while changes in high-density lipoprotein (HDL), HDL2, and apolipoprotein A-I levels with such diets are less gender-specific. Dietary fat restriction appears to exhibit a blunted effect on lipoproteins in women as compared to men, so that this intervention may be less efficacious for women in reduction of cardiovascular risk. In addition, estrogens and androgens have opposite effects on fat distribution, storage, mobilization and transport in all three major lipoprotein pathways (Knopp et al. 2005). Lipoprotein metabolism is approximately two fold more rapid in women as compared with men due to the stimulatory effects of estrogen and the inhibitory effects of androgens. Overall, then, gender differences in IMCL levels would be expected. Body fat distribution, which is know to differ between males and females, has also been established as an important predictor of metabolic abnormalities in obese humans. Recent studies have been performed to investigate gender-specific usage of intramyocellular lipids and found significant differences (Zehnder et al. 2005; Larson-Meyer et al. 2002; White et al. 2003).
In this study we documented higher IMCL levels in female subjects, in agreement with the known higher rates of fat uptake into lower extremity lipid depots in women than in men (Votruba and Jensen, 2006; Horton et al. 2002) as well as with the increased circulation of fatty acids in females (Knopp et al. 2005). We also found a reduced degree of unsaturation in female subjects, in agreement with earlier biochemical studies of serum fatty acids (Warensjo et al. 2006); this investigation also found that men had a higher content of 18:3 (n-6), 20:3 (n-6) and 20:5 (n-3) as compared to females, in agreement with our findings.
In conclusion, the present study and our earlier work (Velan et al. 2007a; Velan et al. 2007b) demonstrate the ability of non-invasive 2-dimensional MR spectroscopy to delineate the saturated and unsaturated triglyceride components within IMCL and EMCL in skeletal muscle. Additionally, we found that these results are sensitive to gender status, consistent with known facts of lipid metabolism. The mechanisms accounting for lower degree of unsaturation within IMCL and EMCL pools of female subjects and the significance of this finding requires further investigation.
Footnotes
Acknowledgements
This work was supported by a Health Sciences Center Grant from West Virginia University and in part by the Intramural Research Program of the National Institutes of Health, National Institute on Aging (RGS) and National Institute of Diabetes and Digestive and Kidney Diseases Research grant DK018777 (VMR).