
Abstract
The main cells of the adipose tissue of animals, adipocytes, are characterized by the presence of large cytosolic lipid droplets (LDs) that store triglyceride (TG) and cholesterol. However, most cells have LDs and the ability to store lipids. LDs have a well-known central role in storage and provision of fatty acids and cholesterol. However, the complexity of the regulation of lipid metabolism on the surface of the LDs is still a matter of intense study. Beyond this role, a number of recent studies have suggested that LDs have major functions in other cellular processes, such as protein storage and degradation, infection, and immunity. Thus, our perception of LDs has been radically transformed from simple globules of fat to highly dynamic organelles of unexpected complexity. Here, we compiled some recent evidence supporting the emerging view that LDs act as platforms connecting a number of relevant metabolic and cellular functions.
Introduction
The major metabolic fates of fatty acids (FA) entering the cell are esterification into triglycerides (TG), synthesis of phospholipids for membrane biosynthesis, and beta-oxidation for ATP production in mitochondria. Adipocytes, however, capture and store the excess of FAs as TG in cytosolic lipid droplets (LDs), which are large quasi-spherical structures readily visualized by light microscopy.1–3 LDs are in physical contact with other LDs and also with other cellular structures such as endoplasmic reticulum (ER), mitochondria, peroxisomes, and endosomes. These interactions are frequently observed by electron microscopy
4
(Fig. 1). Interestingly, live imaging revealed that LDs also undergo intracellular movements in the confines of the cytoplasm.5,6 LDs can be isolated from cell homogenates by density gradient ultracentrifugation7–10 and purified LDs retain their shape and functions, such as the ability to store TG
11
or serve as substrate for lipases
12
when tested in vitro.
Interaction among LDs, mitochondria, and ER. The transmission electron micrographs of 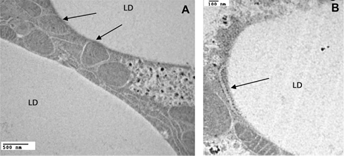
The structure of LDs is commonly described as formed by a core of neutral lipids, predominantly TG and sterol esters, surrounded by a monolayer of phospholipids and a variety of proteins.13–15 TGs are the main lipids found in LDs. Other lipids are diglycerides (DG), retinyl- and cholesteryl esters (CEs), and ether lipids.16–20 A high content of CEs is found in LDs from macrophages and adrenal cells.21–24 The LD surface consists of phospholipids, lysophospholipids, and cholesterol.
16
The phospholipid monolayer is the boundary that separates the hydrophobic core of LDs from the aqueous environment.25,26 It could also be involved in the differential recruitment of lipid droplet proteins.
25
The protein coat is composed of numerous proteins; it can comprise up to hundreds of proteins.
27
A distinctive group of proteins, the perilipins or PAT proteins, are always associated with LDs as shown in all LD proteomes.7–10,28–38 This is a small group of proteins composed by perilipin (PLIN1), ADRP (PLIN2), TIP47 (PLIN3), S3–12 (PLIN4), and OXPAT (PLIN5).39,40 The PAT family of proteins is evolutionary conserved and relevant studies on the biology of LDs and the function of PLIN proteins are being conducted not only in vertebrates but also in other eukaryotic systems such as insects6,10,12,41–49 and yeast.50,51 Early studies on TG metabolism in adipocytes have shown significant conservation between mammals and insects.
52
PLIN1 and PLIN2 occur in insects and were originally named LSD1 and LSD2.
53
PLIN proteins have in common a conserved region of ~100 amino acids toward the N-terminal called the PAT domain.
54
The function of this region remains unknown but it is not required for binding to LD.
52
PLIN proteins play important roles in the processes of lipid storage and mobilization, however the underlying mechanisms of function are not completely known in any system, yet. The occurrence of PLIN proteins depends on a variety of conditions such as type of tissue and the developmental and metabolic stage of the cell. PLIN proteins may also differ in the subcellular localization; while PLIN1 is always found associated with LDs for which they are classified as constitutive LD proteins, PLIN3–5, which are also found in the cytosol, are considered exchangeable LD-proteins.
52
PLIN1 plays major roles in the storage and degradation of TG in adipocytes as indicated by studies in vertebrates55,56 and in insects.52,57 PLIN1 blocks lipolysis under basal conditions, but enables the action of lipases under hormone stimulated lipolysis.
39
So far, only
Since the hallmark discovery that PLIN1 regulates the storage of lipids, 59 the study of LDs has seen a steady increase in attention by the scientific community. In the past decade, LDs have emerged as heterogeneous organelles with great dynamics in size, abundance, and composition in regard to both proteins and lipids. 60 Recent studies have provided more details on LD structure, biogenesis, function, and molecular regulation.3,13,15,61,62 Given the conservation among the basic mechanisms of control of lipid metabolism, 63 studies in insects 41 and other systems have been instrumental to the advancement of this field. LDs are currently associated with multiple cellular roles beyond the storage of lipids. 27 Here we compiled some of the most recent studies with the goal of presenting the LDs as a major platform for the regulation and execution of multiple cellular functions.
Heterogeneity, A Feature of LDs
With the exception of mature white adipocytes from mammalian cells in which a single supersized LD (≫100 μm)
64
is present, LDs of a broad range of sizes (1–100 μm) are observed in both vertebrate and invertebrate adipocytes.
15
Almost all types of cells are able to store TG in LDs, and in general, droplets in non-adipocyte cells are smaller.
65
LDs are subjected to fast and substantial variations in size and number in response to hormonal changes and the availability of nutrients. For instance, a thirty-fold increase in LD-volume is observed within hours when
Enzymes of lipid synthesis are mostly integral membrane proteins of ER. 74 Therefore, the site of the biogenesis of LDs is assumed to be the ER membrane, which harbors the diacylglycerol acyltransferases (DGAT) that are needed to catalyze the last reaction of TG synthesis. The hydrophobicity and low solubility of TG in phospholipids indicate that they would start accumulating in the bilayer core promoting the formation of intramembrane lipid globules that would eventually bud off from the ER to the cytoplasm.62,75 Once the LD is formed, it could accept additional lipids. Live-cell microscopy studies suggested that at least part of TG synthesis takes place in the immediate vicinity of LDs, 11 and that intracellular lipids continuously exit and re-enter LDs. 76
Some studies suggest that the newly formed LDs that appear in adipocytes incubated with FA localize on the cytosolic side of the plasma membrane. These LDs are smaller and coated with different types of perilipins unlike the larger and older cytosolic droplets that are coated with perilipin 1.77–79 Likewise, studies in
LDs are Organelles
The quasi-spherical geometry and the presence of a large lipid core that characterizes both lipoproteins and LDs suggest that they could be compared. In fact, until not long ago, LDs were seen as simple intracellular lipoproteins. It may then be appropriate to point out some of the differences between these two structures. Lipoproteins are assembled in the ER lumen and secreted by the secretory pathway into the circulatory system. 85 LDs also originate from the ER, and although the process of LD assembly is not understood yet, it is clear that nascent LD segregates from the cytoplasmic face of the ER. The synthesis of lipoproteins is restricted to certain types of cells (eg hepatocytes, enterocytes). 86 However, LDs are common organelles present in most types of cells. Some tissues, such as the liver in vertebrates and the fat body of insects store lipids as cytoplasmic LDs and also synthesize and secrete lipoproteins to circulation. 87 Lipoproteins transport lipids among tissues, whereas LDs hold lipids in the cytoplasm. The function of lipoproteins requires a structure that limits their interactions with many cellular and extracellular components, and thus, allows the particle to reach the target tissues. Conversely, LDs serve as a scaffold that recruits proteins involved in lipid metabolism and other metabolic processes. LDs are in contact with elements of cytoskeleton as shown in all LD proteomes.7–10,28–38 The intermediate filament vimentin was one of the first LD-associated proteins identified. 88 Early studies showed a tight association between LDs and vimentin filaments enduring extensive washes with 1% Triton X-100. 89 It has been shown that vimentin is required for proper lipolysis90,91 and for the provision of LD-cholesterol to the mitochondria for steroidogenesis. 92 Cholesterol stored in the LDs of adrenal cells must be delivered to mitochondria, where the first step of steroid synthesis takes place. Genetic ablation of vimentin in mice resulted in a marked defect in steroidogenesis. 92 Vimentin binds to LDs and proteins that are involved in LD metabolism, such as the hormone sensitive lipase, HSL, 91 and the steroidogenic acute regulatory protein that targets mitochondria. 93 Vimentin is considered an important component of the network that facilitates lipolysis and the movement of LDs toward mitochondria. 93
In
The number of proteins associated with the surface of LDs is remarkably large, whereas circulating lipoproteins only contain a discrete number of specific proteins (structural and exchangeable apolipoproteins). Given the variety of proteins associated with LDs, different types of interactions and mechanisms of association of proteins with the LD surface are expected. PLIN1, the best characterized LD-associated protein so far, is synthesized in free ribosomes and it is supposed to localize to LD surface from the cytosol.97–99 More recent information showed that PLIN1 also can relocate to the ER from where the protein moves back and forth between ER and LD.
100
These observations are in accordance with the tight association between LD and ER observed by electron microscopy,51,101,102 and the functional association between LDs and ER that has been demonstrated in yeast
Underlining the dynamic nature of these organelles is the fact that the protein coat of LD undergoes major changes in response to the metabolic state of the cell.8,10 As an illustration, Figure 2 shows protein profiles of LDs isolated from Changes in the protein composition of LDs from the same tissue. Protein profiles of purified LDs from the fat bodies of 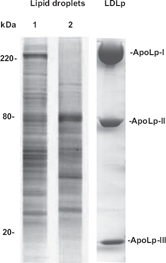
LD Proteomes: First Look at the LD Network
Proteomic studies of purified LD have shown that, in addition to the PLIN proteins, the protein coat of LDs includes a large number of significantly abundant proteins. A complex mixture of proteins from multiple subcellular compartments including cytosol, mitochondria, lysosomes, and ER are found associated with purified LDs. Proteins of the PLIN family, lipid metabolism, nucleus, ribosomes, chaperones, cytoskeleton, ER, membrane trafficking, and cell signaling are commonly identified in LD proteomes.7–10,28–38 The possibility of contamination of the LD preparations is often raised.3,13,52 However, the consistency on these findings in preparations of LDs from different systems clearly argues against that possibility. Electron micrographs from many systems have demonstrated tight physical contact of LDs with other subcellular compartments.10,101,106,107 Most recently, reports of functional interaction between LDs and ER, 103 and LDs and mitochondria, 108 and LDs and peroxisomes 61 add support that these proteomes for the most part are a reflection of the complexity of LDs. Interestingly, LD proteomes show proteins that are known to possess integral membrane regions. This common observation suggests that the LD envelope may have bilayer domains enriched in ER or mitochondrial proteins or, alternatively, may contain domains that are fragments of ER or mitochondrial membranes. It seems obvious now that LDs cannot be simply described as a fat globule surrounded by a monolayer of phospholipid and a discrete number of different proteins anchored to its surface. Electron microscopy has provided evidence consistent with the presence of monolayer structure.2,109 However, that evidence may not apply to some LD population. The complexity of the protein composition of LDs that has been provided by the proteomic studies suggests that LDs have an ample spectrum of interactions with other organelles and a number of cytosolic proteins and protein complexes. This picture of LDs is in line with the increasing evidence supporting a role of LDs in a number of cellular functions. Recent studies showing that structural and functional interactions between LDs and other organelles are of physiological relevance support this notion. 50
LDs as Signaling Platforms Involved in Multiple Cellular Functions
The array of functions related to LDs can be separated into classical and non-classical functions. The former are directly or indirectly linked to lipid storage and mobilization, whereas the second group of functions involving “refugee” proteins are related to vesicular trafficking, protein folding, protein storage, autophagy, immunity, and virus replication. 44 “Refugee” or non-resident proteins are proteins characteristic of other cellular compartment that nonetheless associate with LDs. Numerous refugee proteins have been identified in independent biochemical and proteomic studies.7–10,28–38 An increasing number of mostly recent studies are shedding light into why those proteins are present in LDs and what metabolic and/or cellular conditions cause their association with the organelle.
LDs and whole-body energy homeostasis
The adipose tissue in vertebrates and the fat body in insects play a major role in maintaining the whole body energy homeostasis. Under an energy demand, FA are mobilized from these tissues to supply fuel for other organs. Mammalian adipocytes secrete free FA, whereas insects mobilize FA as DG, which are transported in circulation by lipophorin. 110 On the surface of the lipid droplet is PLIN1, which serves as a gatekeeper controlling the access of lipases to the lipids inside the droplet. Catecholamines in vertebrates 111 and adipokinetic hormones in insects 8 7,11 2 control the mobilization of FA through a cAMP/PKA signaling cascade. PKA-mediated phosphorylation of PLIN1 in vertebrates 113 and LSD1 in insects 12 activates TG hydrolysis. In vertebrates, lipolysis is catalyzed by the concerted action of at least PLIN1 proteins, three lipases (ATGL, HSL, MGL), a co-lipase (CGI58), and two separate pools of PKA.24,114 PKA proximal to plasma membrane phosphorylates HSL triggering the translocation to the surface of LD.22,115 The second PKA seems to be on the surface of LD116,117 apparently recruited by the protein OPA1. 118 A new study showed that this protein, which is known to be a mitochondrial protein, is a putative A-kinase anchoring protein (AKAP) on LDs. 119 Under stimulated conditions, OPA1 seems to operate on the surface of LDs recruiting PKA in connection to PLIN1 phosphorylation and activation of lipolysis. 119
Other kinases such as ERK, p38 MAP, and AMP kinases are also activated by catecholamines120–122 and adipokinetic hormones. 123 For example, HSL can be phosphorylated by ERK which increases its activity,124,125 whereas phosphorylation by AMP kinase is inhibitory.126,127 PLIN1 and HSL have multiple phosphorylation sites. 56 The modulation of the lipolytic response may rely on differential phosphorylation levels of these proteins. 128
The lipolytic activity of a cell must respond to short and long term requirements of its own metabolism, such as FA oxidation and membrane synthesis, but also of the whole body metabolism, for instance for the synthesis of steroid hormones, for the synthesis of lipoproteins, and for the provision of FA to other tissues. Since FA and cholesterol are mobilized from the surface of the LDs, it is not surprising that the LDs harbor a number of lipases, kinases, and regulatory proteins. This complexity is needed to allow the coordination of multiple signals originated from the metabolic requirements of both the cell and the whole organism.
LDs as transient platforms for protein storage
LDs are highly abundant in the oocytes of insects and play an essential role in the reproduction of insects by providing fuel and FA for the development of the embryo. The accumulation of histones in LDs has been detected in proteomic study in early
It has been proposed that other proteins target LDs for temporary shelter. The common presence of ATP-synthase subunits in LDs is intriguing.7–10,132,133 A new study showed that three ATP-synthase subunits interact with invadolysin, 134 which also is a LD-associated protein. 135 Invadolysin is a conserved metalloproteinase required for proper mitochondrial function. 134 It is possible that following their synthesis in the cytoplasm, the inactive ATP-synthase subunits are recruited by the LDs. The interaction with invadolysin on the LD surface could be needed as part of a mechanism of folding and activation of ATP-synthase subunits.
LDs as transient platforms in the path of protein degradation
Misfolded proteins in the ER lumen are removed from the ER via dislocation to the cytosol, where they are degraded by the ubiquitin/proteasome system. This process, which is called ER-associated degradation (ERAD), covers a range of different mechanisms that include protein adaptors whose function is to recruit misfolded proteins to the ubiquitination machinery.30,33 Proteasomes are distributed in the vicinity of LDs, 136 and LDs are involved in the degradation of at least two substrates: lipidated ApoB100 137 and 3-hydroxy-3-methylglutaryl CoA (HMG CoA) reductase, 138 the rate-limiting enzyme of cholesterol synthesis. A role for LDs in ERAD was originally suggested when proteasome inhibition caused the accumulation of ApoB100 at the surface of LDs. 137 Furthermore, LDs might play a broader role in the degradation of misfolded ER proteins since pharmacological inhibition of LD formation impaired the dislocation of other proteins. 145 To eliminate lipidated ApoB100 from the ER lumen, the lipidated particle is transported to the LD surface in a process that involves the proteins Derlin-1 and Ubxd8. 139 It has been suggested that Derlin-1 in the ER membrane and Ubxd8 on the LD surface interact in an ER–LD juncture allowing the passage of ApoB100 to the LD surface for ubiquitination. 139 Ubxd8, a LD-associated protein140–142 recruits VCP/p97, the ATP driven chaperone required to process ubiquitin-labeled proteins. 143 The ubiquitination machinery also involves E3 ubiquitin ligases that use adaptor proteins for substrate recognition. Ancient ubiquitous protein 1 (AUP1), a highly conserved protein that localizes to ER and LDs34,38 and is found in LD proteoms, 8 acts as an adaptor recruiting different ubiquitin ligases such as E2 conjugase Ube2g2, 144 an E3 ligase AMFR/gp78.138,145,146
AUP1 also mediates the sterol-induced ubiquitination of HMG-CoA reductase. 146 AUP1 recruits the ubiquitin-conjugating enzyme Ubc7 to LDs and facilitates its binding to both gp78 and Trc8. The LD mediated degradation of the rate-limiting HMG CoA reductase is important for the control of cholesterol synthesis in mammalian cells. 138
Apart from degradation of ERAD proteins, the abundance of LD-associated proteins PLIN1 and 2 are also controlled by the ubiquitin/proteasome system. 147 In this regard, it has been shown that the multifunctional protein spartin acts as an adaptor needed for the degradation of ADRP that takes place during adipocyte maturation. 148 Housekeeping chaperones such as Hsp70, which are found in all LD proteomes,8–10,31,149,150 may also serve as adaptors to recruit misfolded proteins for proteasomal degradation.
Although the number of examples is limited, the reported studies suggest that degradation of ER proteins on the surface of LDs could be a mechanism controlling the fate of a large number of proteins. The study reported by Klemm and coworkers 145 certainly suggest that this could be the case. As observed for HMG-CoA reductase, the abundance of other membrane enzymes, such as other enzymes involved lipid synthesis could also be controlled by ubiquitination and proteasome activity on the LD surface.
LDs recruit autophagic machinery leading to lipophagy
A number of factors, such as cellular starvation and cellular infection lead to the activation of autophagy. In this process, cytoplasmic components are first engulfed in a vesicular structure and then fused with lysosomes for degradation.70,151 LDs are targets of autophagy and one of the required structural components of the autophagosomes is the protein LC3 covalently conjugated to phosphatidylethanolamine (PE). LC3 is recruited to the surface of LDs, where it is conjugated with PE. 152 Other required proteins of the autophagic machinery, Atg2, Atg5, and Atg7, are also recruited at the LD surface to help the formation of autophagosomes.153,154 In the starved cell, small LDs or portions of larger LDs are directed to lysosomes via autophagosome vesicles in a process called lipophagy.155,156 This process may be important to eliminate misfolded proteins stored in LDs or fragments of membrane proteins that remain in the LD after the proteasomal activity. It may also be important to provide FA through the action of lysosomal acid lipases.152,157 Some studies have shown that alteration of the autophagy process through lysosomal inhibition or knockdown of the expression of the Atg5 gene decreases TG breakdown and increases LD number and size. 153 Although much remains to be known, current information suggests that LDs play an active role in the degradation of cellular components. To this end, LDs seem to provide a platform for the association of both the degradation targets and the degradation machineries of the proteasomal complex and autophagy.
On the role of LDs in cellular infection and immunity
A range of pathogens such as hepatitis C virus (HCV),
158
dengue virus,
159
bacteria
LDs play an important role in the synthesis of eicosanoids,
169
important mediators of the inflammatory response. LDs of macrophages and mast cells store arachidonic acid,170,171 which is the precursor for the synthesis of eicosanoids. LDs of these cells are considered central organelles of the inflammatory response because they are the place for storage and synthesis of prostaglandins and leukotrienes that are secreted during infective processes.
162
Interestingly, LDs of macrophages seem to be structurally different from other LDs.
172
In addition to the protein coat surrounding the droplet, these LDs appear to have internal structures consisting of membranous structures and proteins in the core of the particle.173–175 To what extent this is a unique structural feature of the LDs of macrophages, or a more general case remains to be elucidated. Several pathogens induce the accumulation of LDs in immune cells.161,172,176–179
Conclusion
From simple globules of fat to highly dynamic organelles of unexpected complexity, our perception of LDs has been radically transformed in the last years. Now we recognize LDs as organelles involved in a number of cellular processes beyond their role as site for lipid storage. The broad range of LD functions implicates specific interactions with other cellular compartments and in some conditions the concerted movement of LDs along microtubule tracks. Although the details of the mechanisms for any of the cellular functions in which LDs are involved are limited, protein–protein interactions involving LD-associated proteins emerge as vital components of LD biology. Knowing the full spectrum of LD interactions and signaling mechanisms is essential to understand LD biology and lipid metabolism.
Author Contributions
Wrote the first draft of the manuscript: FZS, ELA. Contributed to the writing of the manuscript: JLS. Made critical revisions and approved final version: ELA, JLS. All authors reviewed and approved of the final manuscript.
Footnotes
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests. Provenance: the authors were invited to submit this paper.