
Abstract
Oxysterols are naturally occurring oxidized derivatives of cholesterol, or by-products of cholesterol biosynthesis, with multiple biologic functions. These compounds display cytotoxic, pro-apoptotic, and pro-inflammatory activities and may play a role in the pathology of atherosclerosis. Their functions as intermediates in the synthesis of bile acids and steroid hormones, and as readily transportable forms of sterol are well established. During the past decade, however, novel physiologic activities of oxysterols have emerged. They are now thought to act as endogenous regulators of gene expression in lipid metabolism. Recently, new intracellular oxysterol receptors have been identified and novel functions of oxysterols in cell signaling discovered, evoking novel interest in these compounds in several branches of biomedical research.
Introduction
Oxysterols are 27-carbon oxidized derivatives of cholesterol or by-products of the cholesterol biosynthetic pathway with multiple biological activities (Björkhem and Diczfalusy, 2002; Javitt, 2008; Schroepfer, 2000). The common modifications of cholesterol that occur in oxysterols are hydroxyl, keto, hydroperoxy, epoxy, and carboxyl moieties. The most important oxysterols are generated in cells catalysed by mitochondrial or endoplasmic reticulum cholesterol hydroxylases belonging to the cytochrome P450 family (Russell, 2000; Luoma, 2007). Of these species, the most abundant in humans are 27-, 24(S)-, 7α-, and 4β-hydroxycholesterol (OHC). Oxysterols also arise in vivo or during food processing through non-enzymatic, free radical, lipid peroxide, or divalent cation-induced oxidative processes, often termed cholesterol autoxidation. The most common oxysterols generated through autoxidation are modified at the 7-position. These include 7-ketocholesterol (7-KC) and 7β-OHC with prominent cytotoxic and pro-apoptotic properties (Lee et al. 2007; Lordan et al. 2007, 2008; Luthra et al. 2008; Martinet et al. 2008). Furthermore, the 7-modified oxysterols as well as 25-OHC have been shown to display pro-inflammatory activities (Erridge et al. 2007; Lemaire-Ewing et al. 2008).
Oxysterols are present in healthy human or animal tissues at very low concentrations as compared to cholesterol, but are found enriched in pathologic cells/tissues such as macrophage foam cells, atherosclerotic lesions, and cataracts (Brown and Jessup, 1999; Olkkonen and Lehto, 2004; Javitt, 2008). They have been studied intensively in the context of the oxidative hypothesis of atherosclerosis. Futhermore, their functions as intermediates in the synthesis of bile acids and steroid hormones, and as readily transportable forms of sterol are well established. During the past decade, however, novel physiologic activities of these compounds have emerged. Oxysterols are now thought to act as endogenous regulators of gene expression in lipid metabolism. Recently, new intracellular oxysterols receptors have been identified and novel functions of oxysterols in cell signaling discovered, evoking novel interest in these compounds in several branches of biomedical research.
Are Oxysterols in vivo Ligands for the Liver X Receptors?
Oxysterols have been shown to act in vitro as ligands of liver X receptors (LXR), nuclear receptors with central roles in sterol absorption in the intestine, the reverse cholesterol transport process, bile acid synthesis, biliary neutral sterol secretion, hepatic lipogenesis, and synthesis of nascent high-density lipoproteins (Tontonoz and Mangelsdorf, 2003; Zelcer and Tontonoz, 2006). While there has been some controversy as to whether oxysterols also are the endogenous in vivo ligands of the LXR (Björkhem and Diczfalusy, 2002), the recent work by Chen et al. (2007) provided convincing evidence that this indeed is the case: Overexpression of an oxysterol catabolic enzyme, cholesterol sulfotransferase, was shown to inactivate LXR signaling in several cultured mammalian cell lines but did not alter receptor response to the nonsterol LXR agonist T0901317. Moreover, triple-knockout mice deficient in the biosynthesis of three oxysterol ligands of LXRs, 24(S)-, 25-, and 27-OHC, responded to dietary T0901317 by inducing LXR target genes in the liver but showed impaired responses to dietary cholesterol, strongly suggesting that conversion of cholesterol to oxysterols is important for LXR activation in vivo.
Oxysterols Regulate SREBP Function via Binding to the Insig Proteins
The cellular machinery for cholesterol biosynthesis and uptake, as well as for fatty acid biosynthesis, is controlled by transcription factors designated sterol regulatory element binding proteins (SREBPs; Eberle et al. 2004; Goldstein et al. 2006). The intracellular localization and proteolytic maturation of SREBPs is regulated by a cholesterol-sensing accessory protein called the SREBP cleavage activating protein (SCAP). The SREBPs are synthesized as precursors anchored to ER membranes and form complexes with SCAP. Under low-cholesterol conditions, SREBP-SCAP complexes are transported by a COPII-dependent mechanism to the Golgi complex, where SREBPs are proteolytically processed to release a basic helix-loop-helix leucine zipper transcription factor, which enters the nucleus and binds to sterol regulatory elements (SRE) in the promoter regions of target genes. When sterol builds up in cells, SCAP senses cholesterol in the ER membranes and interacts with Insig (Insulin-induced gene) proteins, and as a result the SREBP-SCAP complex is retained in the ER. The transport of SREBP to the Golgi complex is sensitive to both cholesterol and several oxysterols, of which 25-OHC is most commonly used in experimental set-ups. Intriguingly, SCAP was not found to bind 25-OHC (Adams et al. 2004; Brown et al. 2002), raising the question of how the inhibitory effect of oxysterols could be mediated. Recent reports by the group of M.Brown and J.Goldstein provide a plausible solution to this dilemma: The Insig proteins were found to directly bind 25-OHC and to mediate the regulatory effect of this oxysterol on SREBP processing. Binding of the Insig-25-OHC complex to SCAP elicits a conformational change similar to that induced by cholesterol binding to SCAP. This conformational change precludes the interaction of SCAP with COPII proteins and thereby the transport and proteolytic activation of SREBP (Radhakrishnan et al. 2007; Sun et al. 2007). Interestingly, Lange et al. (2008) found that the mitochondrial cholesterol-27-hydoxylase is required for the rapid inactivation of 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGCoAR) in response to high levels of cholesterol. This may be due to 27-OHC interaction with Insig, resulting in the ubiquitination and degradation of HMGCoAR (see Goldstein et al. 2006). Vertebrate cells thus have two sterol sensors that control SREBP activity, enabling cells to downregulate their sterol biosynthesis upon the build-up of either cholesterol or its oxidized derivatives.
24(S), 25-Epoxycholesterol, a Signal for Sterol Biosynthetic Pathway Activity
24(S), 25-epoxycholesterol [24(S),25-EPOX] is generated as a side product of the cholesterol biosynthetic process by the same enzymes that catalyze the synthesis of cholesterol (Nelson et al. 1981). 24(S),25-EPOX is a potent feedback regulator of cholesterol biosynthesis, suppressing SREBP-2 processing (Janowski et al. 2001) and the cellular HMGCoAR activity (Mark et al. 1996). It is also a highly potent oxysterol activator of the LXRs (Chen et al. 2007; Janowski et al. 1999; Lehmann et al. 1997). The cellular level of 24(S), 25-EPOX can be manipulated either by using inhibitors of the cholesterol biosynthetic pathway enzyme 2,3-oxidosqualene cyclase (OSC), resulting in elevation of cellular 24(S), 25-EPOX levels, or by overexpressing this enzyme to reduce the cellular content of the oxysterol. Using these tools and other approaches the groups of A. Brown and M. Huff have significantly added to our understanding of the physiologic role of 24(S), 25-EPOX. Beyea et al. (2007) demonstrated that partial inhibition of OSC in THP-1 macrophages reduced cholesterol synthesis and increased the expression of several LXR target genes, ABCA1, ABCG1, and APOE (apolipoprotein E). Importantly, OSC inhibition did not stimulate lipoprotein lipase (LPL) or fatty acid synthase (FAS), and the observed induction of the lipogenic transcription factor SREBP-1c was counteracted by a block in its conversion to the active nuclear, supporting the notion that OSC inhibition might have therapeutic potential (Rowe et al. 2003). However, this idea is undermined by the finding that, even though 24(S), 25-EPOX facilitates ABCA1 and G1 expression and cholesterol efflux from normal macrophages in culture, it strongly inhibits cholesterol efflux from macrophage foam cells, possibly due to inhibition of cholesterol ester hydrolase function (Ouimet et al. 2008). The findings of Wong et al. (2007a, 2008) suggest that synthesis of 24(S), 25-EPOX parallels that of cholesterol and acts as a signal that fine-tunes the acute control of cellular cholesterol homeostasis and protects cells against accumulation of newly synthesized cholesterol. This occurs via impacts of the oxysterol on both the LXRs and the SREBPs.
Interestingly, 24(S), 25-EPOX is also suggested to have a specific role in CNS sterol homeostasis. The data by Wong et al. (2007b) demonstrate that this oxysterol is produced by astrocytes in vitro and is taken up by neurons in which it impacts gene expression. The authors suggested that 24(S), 25-EPOX may act as a signal from astrocytes that reduces the energetically costly cholesterol biosynthesis by neurons, enabling the neurons to divert resources into other cellular processes.
27-Hydroxycholesterol Acts as a Selective Estrogen Receptor Modulator
Estrogen receptors are nuclear receptors that, in addition to central reproductive functions, mediate estrogen regulation of a number of other physiologic processes (Deroo and Korach, 2006). Much of the cardiovascular protection observed in premenopausal females has been attributed to beneficial effects of estrogen on endothelial function and the lipid profile (Mendelsohn and Karas, 1999; Murphy and Steenbergen, 2007). Recently, it was discovered that 27-OHC antagonizes the estrogen-dependent production of NO by vascular cells, resulting in reduced vasorelaxation of rat aorta, a potentially deleterious effect (Umetani et al. 2007). Moreover, increasing 27-OHC levels repressed carotid artery re-endothelialization. Also cell type-specific pro-estrogenic actions of 27-OHC were reported (DuSell et al. 2008; Umetani et al. 2007), indicating that this oxysterol acts as an endogenous selective estrogen receptor modulator (SERM). The findings suggest that 27-OHC may counteract the estrogen protection from vascular disease, thus revealing a novel, potentially harmful physiologic oxysterol function.
Oxysterols Regulate Developmental Processes via Impacts on Hedgehog Signaling
The Hedgehog (Hh) signaling pathway plays a key role in the patterning of metazoan embryos, in post-embryonic development, as well as in the homeostasis of adult tissues and stem cell physiology (Jacob and Lum, 2007; Varjosalo and Taipale, 2007). Corcoran and Scott (2006) reported evidence that cholesterol or certain oxysterols are required for Sonic hedgehog pathway signal transduction and proliferation of medulloblastoma cells. Further, Dwyer et al. (2007) demonstrated that naturally occurring oxysterols exert osteoinductive effects through activation of the Hh signaling pathway. Consistently, Kim et al. (2007) provided evidence that the inhibition of bone marrow stromal cell differentiation into adipocytes by 20(S)-OHC occurs through a Hh-dependent mechanism. These findings introduce a novel role of oxysterols as regulators of mammalian development. Interestingly, they may be connected with the observation that defects in the cholesterol biosynthetic pathway, such as those in Smith-Lemli-Opitz syndrome, can result in severe congenital abnormalities (Yu and Patel, 2005). The pathology of SLOS has been suggested to involve the Hh signaling pathway: The active Hh proteins contain a covalently linked cholesterol molecule (Jeong and McMahon, 2002). Hence, disturbances in the biosynthesis of cholesterol and of oxysterol derived therefrom could lead to defective Hh signaling during embryonic development, potentially causing congenital defects.
The Niemann-Pick C1 Protein Binds Both Cholesterol and Oxysterols
Niemann-Pick C (NPC) disease is characterized by accumulation of free cholesterol and sphingolipids within late endocytic compartments of cells (Sturley et al. 2004). The disease is caused by mutations in either of two proteins, NPC1 or NPC2. NPC1 is a multi-spanning membrane protein localized in late endosomes and has a consensus cholesterol binding motif consisting of five trans-membrane helices. The mechanism by which NPC1 facilitates egress of lipids from late endocytic compartments is poorly understood. Recently, NPC1 was found to bind not only cholesterol but also 25-OHC, the binding of which was efficiently competed by 24(S)-OHC and 27-OHC (Infante et al. 2008a). The authors found that cholesterol and 25-OHC bind not to the trans-membrane helix motif but to a luminal loop of NPC1 (Infante et al. 2008b). The relevance of this sterol binding for the function of NPC1 remained unclear in experiments employing cultured fibroblasts, and NPC1 was shown to be dispensable for the ability of 25-OHC to regulate SREBP processing. However, one can envision that oxysterol binding by NPC1 could fine-tune the function of the protein in routing lipids out of late endosomes.
ATP-Binding Cassette Transporters Mediate the Cellular Egress of Oxysterols
Several oxysterols, especially 7-KC and 7β-OHC, induce apoptosis or necrosis in vascular cells, with putative adverse consequences on the development and the stability of atherosclerotic lesions (Colles et al. 2001; Olkkonen and Lehto, 2004). Terasaka et al. (2007) demonstrated that ABCG1 has a specific role in reducing the susceptibility of macrophages to oxidized LDL- or 7-KC-induced apoptosis, and in promoting the cellular efflux of 7-KC and 7β-OHC to large high-density lipoprotein (HDL) acceptor particles. Consistent findings were reported by Engel et al. (2007). ABCA1 and lipid-free apolipoprotein A-I were shown to play no role in these efflux events. However, other oxysterols such as 25-OHC can also be effluxed by ABCA1 (Tam et al. 2006; Terasaka et al. 2007). Importantly, Abcg1-/- mice fed a Western diet displayed accumulation of 7-KC in peritoneal macrophages. The findings suggest that the ability of large HDL particles to promote 7-KC efflux via ABCG1 may protect our vascular system from deleterious effects of oxysterols derived from the diet or from lipoproteins oxidized in vivo.
The Cytoplasmic Oxysterol-Binding Protein Family–-Novel Functions in Cellular Lipid Metabolism and Signaling
Families of proteins displaying sequence homology to the cytoplasmic oxysterol-binding protein, OSBP, are present throughout the eykaryotic kingdom. In humans the family consists of 12 members. These proteins, designated OSBP-related (ORP) or OSBP-like (OSBPL) proteins, have been implicated in the control of cellular lipid metabolism, vesicle transport, and cell signaling (reviewed by Yan and Olkkonen, 2008). In addition to OSBP, several other family members in both mammals (Suchanek et al. 2007) and in S. cerevisiae (Im et al. 2005; Raychaudhuri et al. 2006) have been shown to binds sterols as their ligands. However, the mechanisms of their action have remained poorly understood. In the following I summarize the most important findings on ORP function published in 2007 and 2008.
OSBP in the control of hepatic lipogenesis and cellular cholesterol efflux
OSBP, the founder member of the ORP family, acts as a sterol sensor involved in the regulation of sphingomyelin synthesis (Perry and Ridgway, 2006) and of extracellular signal regulated kinase (ERK) activity (Wang et al. 2005). Adenoviral overexpression of rabbit OSBP in mouse liver was shown to result in an increase of plasma very-low-density lipoprotein (VLDL) and liver tissue triglycerides (TG) (Yan et al. 2007b). The increase of plasma TG was attributed to an increase of hepatic TG secretion. Analysis of the underlying mechanism revealed up-regulation of SREBP-1c expression and increase of the active nuclear form of this lipogenic transcription factor in the OSBP-transduced liver. Silencing of OSBP by RNA interference in cultured hepatocytes attenuated the insulin induction of SREBP-1c and fatty acid synthetase (FAS), as well as TG synthesis. Furthermore, OSBP overexpression was shown to inhibit phosphorylation of the extracellular signal-regulated kinases (ERK; see also Wang et al. 2005). In the light of the finding that changes in ERK activity impact the stability of nuclear SREBP-1c (Botolin et al. 2006), this provides one tentative mechanistic explanation to the OSBP overexpression phenotype. The findings demonstrate a new role of OSBP as a regulator of hepatic TG metabolism and a putative function in insulin-induced signaling cascades.
A recent report by Bowden and Ridgway (2008) reported that silencing of OSBP by RNA interference resulted in increased cellular amount and cholesterol efflux activity of ABCA1, in the absence of effects on the ABCA1 mRNA level or LXR activity. OSBP knock-down was shown to increase the half-life of the ABCA1 protein, the effect being dependent on an intact OSBP sterol-binding domain. Thus, it seems that OSBP opposes the activity of LXR by destabilizing ABCA1, thereby acting in parallel with ORP8 (see below).
Related with the above report on OSBP, Yan et al. (2008) demonstrated that silencing of ORP8 expression in THP-1 macrophages induces the transcription of ABCA1 and, consequently, cholesterol efflux to apolipoprotein A-I. This effect was reproduced using a luciferase reporter assay, in which ORP8 silencing synergized with a synthetic LXR agonist and was significantly suppressed when a mutant ABCA1 promoter devoid of a functional LXR response (DR4) element was used, providing the first solid piece of evidence for a functional interplay between the LXR and the ORPs. Importantly, ORP8 was found to be abundant in the macrophages of human coronary artery lesions, bringing up the possibility that ORP8 may play a role, possibly an adverse one, in the development of atherosclerotic lesions. Why cellular oxysterol receptors (LXR vs. OSBP and ORP8) should counteract each other in ABCA1 regulation is somewhat unclear–-One possibility is that they could respond to different oxysterol signals in the nucleus and the cytoplasm, respectively, resulting in a more fine-tuned regulation of ABCA1 expression.
New roles for OSBP in JAK-2/STAT3 signaling and amyloid precursor protein processing
The group of R. Anderson (Wang et al. 2005, 2008) identified OSBP as a sterol-sensing scaffolding factor that regulates the dephosphorylation and hence the activity of the ERK, key components of the mitogen activated protein kinase (MAPK) signaling pathways. Another signaling function of OSBP was recently discovered by Romeo and Kazlauskas (2008), who found that up-regulation of profilin-1, an actin-binding protein implicated in endothelial dysfunction and atherosclerosis, by 7-KC is mediated by OSBP. The signaling route involves interaction of the OSBP-7-KC complex with the tyrosine kinase JAK-2, which phosphorylates Tyr394 on OSBP. This apparently leads to the activation of STAT3 responsible for the induction of profilin. An intriguing implication of these findings is that also other members of the ORP family could have lipid-specific scaffolding functions in signaling pathways. Consistent with this idea, Lessman et al. (2007) demonstrated that ORP9 contains a phosphoinositide-dependent kinase-2 (PDK-2) phosphorylation site, the phosphorylation of which was dependent on PKC-β or mTOR in bone marrow-derived mast cells or HEK293 cells, respectively. Furthermore, the authors provided evidence that ORP9 interacts with these kinases to negatively regulate phosphorylation of the PKD-2 site in Akt/protein kinase B, a major controller of cell cell survival, cell cycle progression, and glucose metabolism.
An entirely novel function of OSBP was recently reported by Zerbinatti et al. (2008). The authors demonstrated that OSBP overexpression in a neuroglioma cell line and in HEK293T cells downregulated the processing of amyloid precursor protein (APP) to β-amyloid (Aβ), while OSBP silencing had the opposite effect. Furthermore, OSBP overexpression caused the sequestration of APP-Notch2 heterodimers in the Golgi complex, an effect reversed by addition of the OSBP high-affinity ligand 25-OHC. This fits nicely with the fact that the distribution of OSBP itself between cytosol/ER and Golgi membranes is regulated by the cellular sterol status (Ridgway et al. 1992; Mohammadi et al. 2001), and suggests that OSBP has the capacity to modulate the intracellular trafficking of APP, possibly also as dimers with other interaction partners. OSBP could thus play a pivotal role in APP metabolism when cellular cholesterol is abundant, by keeping Aβ production low.
ORP1L regulates dynein/dynactin motor recruitment on late endosomes and macrophage lipid metabolism
Johansson et al. (2007) employed ORP1L knock-down with siRNA to show that the protein is required for the clustering of late endocytic compartments in the pericentriolar region. The protein was shown to form a complex with the late endosomal GTPase Rab7 and its second effector protein RILP. The tripartite complex apparently recruits dynein/dynactin motor to late endosome membranes to drive minus-end directed motility of the compartments. The role of ORP1L in this process may be to transfer the motor complex to its actual membrane receptor, αIβIII-spectrin. Even though the sterol regulation of the process remained unexplored, one can envision that ORP1L may act as a sterol sensor regulating the subcellular distribution and transport functions of late endocytic compartments. Such regulation could, for example, affect the egress of lipids from these compartments dependent on NPC1 and NPC2.
Yan et al. (2007a) showed that transgenic macrophages overexpressing ORP1L increased the size of atherosclerotic lesions in LDL receptor deficient mice. The transgenic macrophages displayed a defect in cholesterol efflux to spherical high-density lipoproteins (HDL) and reduced expression of ABCG1 and apolipoprotein E, as well as increased expression of phospholipid transfer protein (PLTP). All these genes are subject to transcriptional regulation by the LXRs. Furthermore, ORP1L overexpression in cultured macrophages was shown to attenuate the response of the ABCG1 mRNA to the LXR agonist 22(R)OHC, which is also a ligand of ORP1L. One interpretation of the results is that ORP1L could modulate the LXR ligand interactions, thereby affecting the expression of LXR target genes and the development of atherosclerosis. However, it is also possible that other, more indirect mechanisms account for the observed phenotypic effect. Whether these findings are related to the above function of ORP1L in dynein/dynactin recruitment on endosome membranes is as yet unclear, but one is tempted to speculate that ORP1L could affect sterol-regulated gene expression by impacting the transport and hydroxylation of cholesterol originating from the endosomes.
ORP3 interacts with R-Ras and regulates cell adhesion
ORP3 and ORP7 were recently found to interact physically with R-Ras, a small GTPase that regulates cell adhesion and migration (Goldfinger et al. 2007), implying a putative role of these ORPs in R-Ras signaling. Lehto et al. (2008) reported that ORP3 controls cell adhesion and spreading, organization of the actin cytoskeleton, β1-integrin activity and macrophage phagocytic function, cellular processes also subject to regulation by R-Ras. Moreover, ORP3 was shown to be a phosphoprotein, the phosphorylation of which is apparently regulated by outside-in signals mediated by integrins and cadherins. ORP3 is expressed abundantly in leukocytes and in several epithelia, and its abnormally high expression is detected in certain forms of leukemia and solid tumors, suggesting that the protein may modify cell signaling and adhesion properties in a manner that facilitates malignant growth.
S. cerevisiae Osh4p modulates the golgi phosphatidylinositol-4 phosphate pool and its accessibility for protein interactions
Deletion of the S. cerevisiae ORP OSH4/KES1 leads to by-pass of the temperature-sensitivity of mutants in SEC14, a gene encoding a phosphatidylinositol transfer protein (PITP; Sec14p) essential for secretory vesicle biogenesis (Fang et al. 1996; Li et al. 2002). Osh4p thus acts as a negative regulator of Golgi secretory function, but the underlying mechanism has remained poorly understood. A recent study by the group of C. McMaster shows that Osh4p reduces both the cellular content of phosphatidylinositol-4 phosphate (PI4P) and its availability for recognition by other proteins, which include components required for transport vesicle formation (Fairn et al. 2007). How binding of sterols by Osh4p (Im et al. 2005) and its ability to transfer them between membranes (Raychaudhuri et al. 2006) relate to the function of the protein in Golgi vesicle transport is not clear. The ability of osh4 mutants to bind sterol did not correlate with their ability to complement osh defect in vesicle transport and cell viability (Im et al. 2005), suggesting that there is no simple interdependency between the two functions.
Future Perspectives
In addition to the roles of oxysterols as potentially harmful substances that accumulate in pathophysiologic states, work carried out during the past decade has revealed their physiologic functions as signaling molecules that maintain cellular and body lipid homeostasis and determine cell fate. The discovery of oxysterol interactions with new receptors involved in lipid metabolism, such as the Insigs, the estrogen receptors, NPC1 and the OSBP-related proteins, paves the way for future work to elucidate in detail the mechanisms of oxysterol action in lipid metabolism and in diseases such as atherosclerosis and Alzheimer's disease. Discovery of oxysterols as regulators of Hedgehog signaling uncovered their role in embryonic development. This finding has opened new perspectives: We can expect a plethora of novel connections of oxysterols and their cellular receptors with differentiation and developmental processes. The currently known major functions of oxysterols are summarized in Figure 1.
A summary of the major biological functions of oxysterols.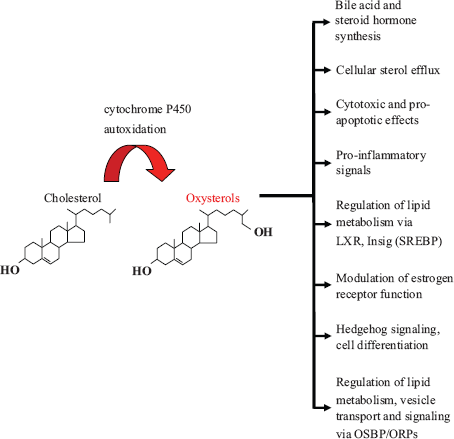
Recent studies in various organisms have uncovered new roles of ORP proteins as regulatory factors involved in with cellular lipid metabolism and signaling. The interest in this protein family is constantly expanding, and the recent groundbreaking studies have paved the way for creating new functional hypotheses. ORP gene silencing in cultured cells and live animals as well as gene-deficient mouse models will be instrumental in reaching a detailed understanding of ORP function. Even though many activities of these proteins involve cellular lipid metabolism, it is obvious that numerous new connections of the ORPs with the control of intracellular vesicle transport, cell differentiation, proliferation, polarity, adhesion, migration, and survival/death, will de discovered.
Footnotes
Acknowledgments
Work in the author's group is supported by the Academy of Finland (grant 121457), the Sigrid Juselius Foundation, the Finnish Foundation for Cardiovascular Research, and the Magnus Ehrnrooth Foundation.