
Abstract
Background
The etiology and pathomechanism of chronic fatigue syndrome/myalgic encephalomyelitis (CFS/ME) are unknown. However, natural killer (NK) cell dysfunction, in particular reduced NK cytotoxic activity, is a consistent finding in CFS/ME patients. Previous research has reported significant changes in intracellular mitogen-activated protein kinase pathways from isolated NK cells. The purpose of this present investigation was to examine whether protein kinase genes have a role in abnormal NK cell intracellular signaling in CFS/ME.
Method
Messenger RNA (mRNA) expression of 528 protein kinase genes in isolated NK cells was analyzed (nCounter GX Human Kinase Kit v2 (XT); NanoString Technologies) from moderate (n = 11; age, 54.9 ± 10.3 years) and severe (n = 12; age, 47.5 ± 8.0 years) CFS/ME patients (classified by the 2011 International Consensus Criteria) and nonfatigued controls (n = 11; age, 50.0 ± 12.3 years).
Results
The expression of 92 protein kinase genes was significantly different in the severe CFS/ME group compared with nonfatigued controls. Among these, 37 genes were significantly upregulated and 55 genes were significantly downregulated in severe CFS/ME patients compared with nonfatigued controls.
Conclusions
In severe CFS/ME patients, dysfunction in protein kinase genes may contribute to impairments in NK cell intracellular signaling and effector function. Similar changes in protein kinase genes may be present in other cells, potentially contributing to the pathomechanism of this illness.
Keywords
Introduction
Calcium (Ca2+) ions play an integral role in intracellular signaling. Ca2+ controls a diverse range of cellular processes, such as gene transcription, muscle contraction, and cell proliferation. 1 The effects of Ca2+ ions are mediated by the Ca2+-binding protein calmodulin, which activates a number of different protein kinases. Ca2+/calmodulin-dependent protein (CaM) kinase includes myosin light chain kinase, which signals muscle contraction and members of the CaM kinase family, which phosphorylates a number of different proteins, including metabolic enzymes, ion channels, and transcription factors. 2
In natural killer (NK) cells, Ca2+ signaling plays an important role in the granule-dependent pathway of apoptosis. 3 Ca2+ is required for inducing cytolytic granule polarization, cytokine gene transcription, and degranulation in NK cells.4,5 Ca2+ also regulates lytic granule fusion,4–6 as well as lytic granule mobilization to the immune synapse to release perforin and granzymes to kill target cells.3,7,8 Furthermore, the downstream intracellular signals that occur in NK cells trigger lysis of target cells and are governed by the Ca2+-regulated mitogen-activated protein kinase (MAPK) pathway. 9 Intra-cellular Ca2+ concentrations can either stimulate or inhibit the MAPK cascade, and thereby play an important role in the regulation of MAPK-dependent cellular processes. The effector functions of NK cells are regulated by three specific MAPK subgroups10–12: the p38 MAPK (p38); the C-Jun terminal kinase (JNK), which regulate cytokine production; and extracellular signal regulated kinases (ERK1/2). These kinases regulate the mobilization and redistribution of cytoplasmic perforin and granzyme B toward the contact zone with target cells. 13 Interestingly, our recent findings demonstrated impairments in MAPK signaling, as well as decreased intracellular Ca2+ concentration in isolated NK and B-cells from chronic fatigue syndrome/myalgic encephalomyelitis (CFS/ME) patients.14,15 Collectively, these anomalies may contribute to NK cell dysfunction, in particular the reduced NK cell cytotoxic activity that is consistently reported in CFS/ME patients.16–24
Given the importance of Ca2+ signaling in regulating NK cell function, the present study aimed to examine the role of protein kinase genes in isolated NK cells from CFS/ME patients. The study further explores the association of functionally important NK intracellular signaling alterations with CFS/ME.
Methodology
Participants
Participants were recruited from the National Centre for Neuroimmunology and Emerging Diseases research database for CFS/ME. All participants completed an online questionnaire regarding their medical history and symptoms based on the 2011 International Consensus Criteria (ICC) to determine suitability for the study. 25 This requires the presence of debilitating postexertional fatigue, accompanied further by neurological, immune, and autonomic symptoms. CFS/ME patients meeting the 2011 ICC symptoms and nonfatigued healthy controls were included in this study. Severity of CFS/ME was defined according to Dr Bell's disability scale that ranges from 100% (no symptoms) to 0% (severe symptoms). 26 Patients categorized as moderate CFS/ME scored >30%. Severe CFS/ME patients scored <30% and were considered housebound or bedridden. Patients’ disability was further characterized by self-reporting scales that included the fatigue severity scale (FSS) and the SF–36.27,28 Participants were excluded if they were previously diagnosed or had a history of any alternative disease that would explain symptoms including autoimmune disorder, multiple sclerosis, psychosis, major depression, heart disease, or thyroid-related disorders, or if they were pregnant, breast feeding, or smokers. All the participants provided written informed consent and donated within a one-month period. The study was performed during this month under the approval of the Griffith University Human Research Ethics Committee (HREC/15/QGC/63). The study was performed during this month under the approval of the Griffith University Human Research Ethics Committee (HREC/15/63) and complied with the principles of the Declaration of Helsinki.
Blood collection
A total of 40 mL of whole blood was collected from the antecubital vein of participants into EDTA blood collection tubes. All the laboratory analyses were performed within six hours of blood collection to maintain cell viability. Routine pathology testing parameters including full blood count, erythrocyte sedimentation rate, electrolytes, and high-sensitivity C-reactive protein were also assessed on each participant sample by Queensland pathology.
NK cell isolation
NK cells were isolated from 40 mL of whole blood using negative selection with RosetteSep Human Natural Killer Cell Enrichment Cocktail (STEMCELL Technologies Australia Pty. Ltd) according to manufacturer's instructions. The isolated NK cells were incubated in unsupplemented Roswell Park Memorial Institute-1640 culture media (Life Technologies) and were counted using Trypan blue. Briefly following NK cell isolation (Supplementary Fig. 1), the NK cell purity was also assessed on the LSR-Fortessa X-20 flow cytometer (BD Biosciences) after labeling with CD3 and CD56, as previously described in the study by Marshall-Gradisnik et al. 29 (BD Biosciences). The isolated NK cells were lysed at a concentration 10,000 cells/µL of RLT buffer (Cat No: 79216; QIAGEN) and were snap frozen in liquid nitrogen and stored at −80 °C until further protein kinase genes assessment.
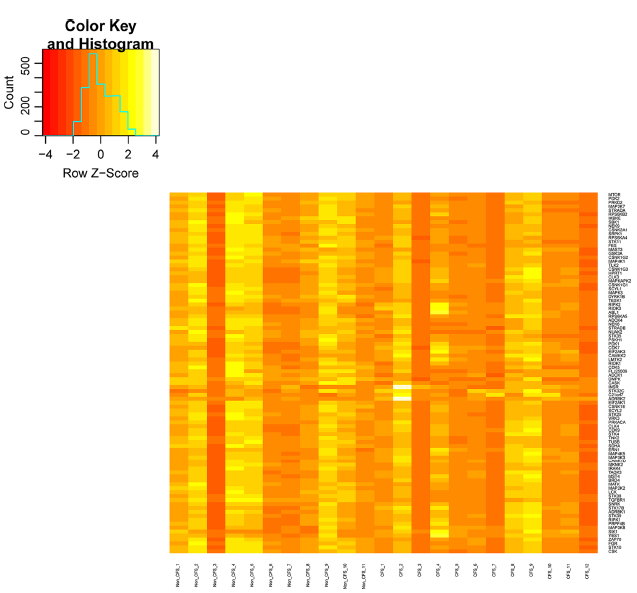
Heat map of protein kinase gene expression showing significantly upregulated and downregulated genes from severe CFS/ME patients compared with nonfatigued controls.
Protein kinase gene expression
Gene expression was directly measured via counts of corresponding messenger RNA (mRNA) in each sample using an nCounter (NanoString Technologies) GX human kinase kit v2 (XT), which is a multiplex assay for 528 protein kinase genes known to be differentially expressed in the human kinome. 30 The nCounter system allows for direct detection and counting of nucleic acid via reporter probes appended with multiple fluorophore barcodes and biotinylated capture probes that attach to microscopic beads. These are affixed to lanes in a translucent cartridge and read in an optical scanner. Batches of 12 separate samples at one time were prepared as per manufacturer's instructions. NK cell lysate was hybridized with probes at 65 °C for 16–18 hours before being placed into the automated nCounter Prep Station (NanoString Technologies), in which samples were affixed to cartridges. Cartridges were immediately placed into the nCounter Digital Analyzer (NanoString Technologies) optical scanner and read at a goal resolution of 550 fields of view, which is the maximum resolution for this instrument.
Statistical analysis
Statistical analysis was performed using SPSS software version 22 and GraphPad Prism version 6. Data were compared among the three groups (control, moderate CFS/ME, and severe CFS/ME) with statistical analysis performed based on the distribution. Shapiro–Wilk normality tests were performed on all the data sets to test for Gaussian distribution. Analysis of variance was used to examine parametric data and the Kruskal–Wallis test of independence was performed for nonparametric data when appropriate, with statistical significance set at an alpha criterion at a P-value of <0.05. Gene expression was directly measured via counts of corresponding mRNA in each sample using DESeq R package software, 31 where alpha level of significance was set at a P-value of <0.05.
Results
Participants
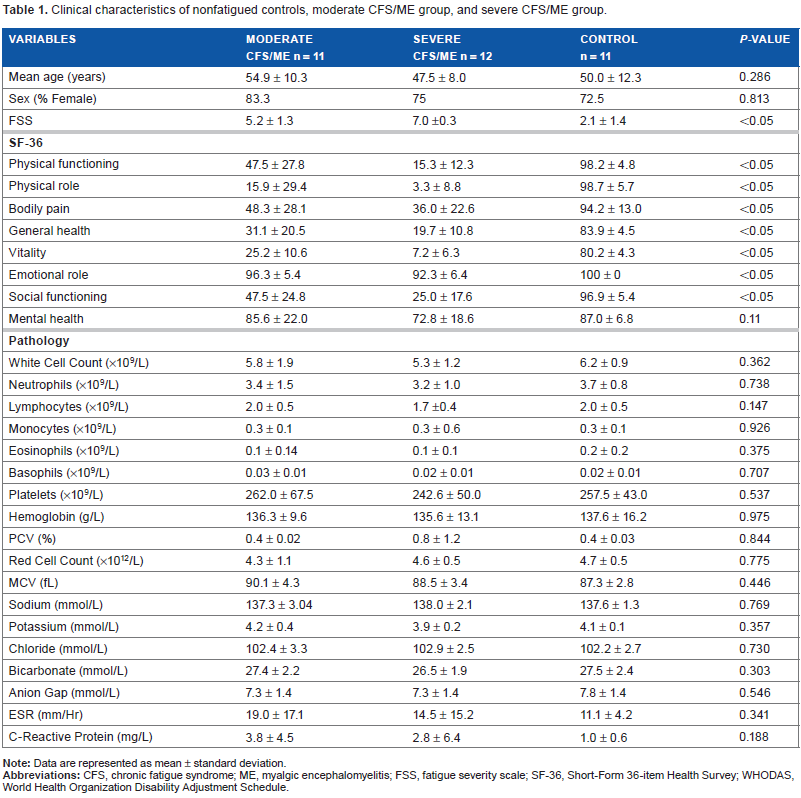
Table 1 summarizes the clinical characteristics of the participants. The study included 11 moderate CFS/ME (age, 54.9 ± 10.3 years; 83.3% female) and 12 severe CFS/ME (age, 47.5 ± 8.0 years; 75.0% female) patients and 11 nonfatigued controls (age, 50.0 ± 12.3 years; 72.5% female). There was no significant difference between mean age and sex between groups. All participants in the study were of European decent and were residents of Australia at the time of blood collection. Fatigue severity was highest among severe CFS/ME patients, compared with moderate CFS/ME patients and nonfatigued controls (P < 0.05). Furthermore, severe CFS/ME reported significantly greater impairment across all SF-36 scales compared with moderate CFS/ME patients and nonfatigued controls (P < 0.05), except for general mental health (P = 0.11). Comparison of the group age and blood parameters including erythrocyte sedimentation rate, high-sensitivity C-reactive protein, and full blood counts for white and red blood cells between participant groups showed no significant difference.
Clinical characteristics of nonfatigued controls, moderate CFS/ME group, and severe CFS/ME group.
Ca2+-dependent protein kinase gene expression
Microarray analysis of the 528 protein kinase genes revealed that there were 92 genes, which were significantly associated with severe CFS/ME patients compared with nonfatigued controls. Of the 92 genes, 37 genes were significantly upregulated (Table 2) and 55 genes were significantly downregulated (Table 3) in severe CFS/ME patients compared with nonfatigued controls. A heat map of gene expression with clustering using Spearman's correlation in severe CFS/ME patients and nonfatigued controls is shown in Figure 1. There was no significant alteration in the expression of protein kinase genes in moderate CFS/ME patients compared with nonfatigued controls.
List of protein kinase genes significantly upregulated in severe CFS/ME group compared with nonfatigued controls.
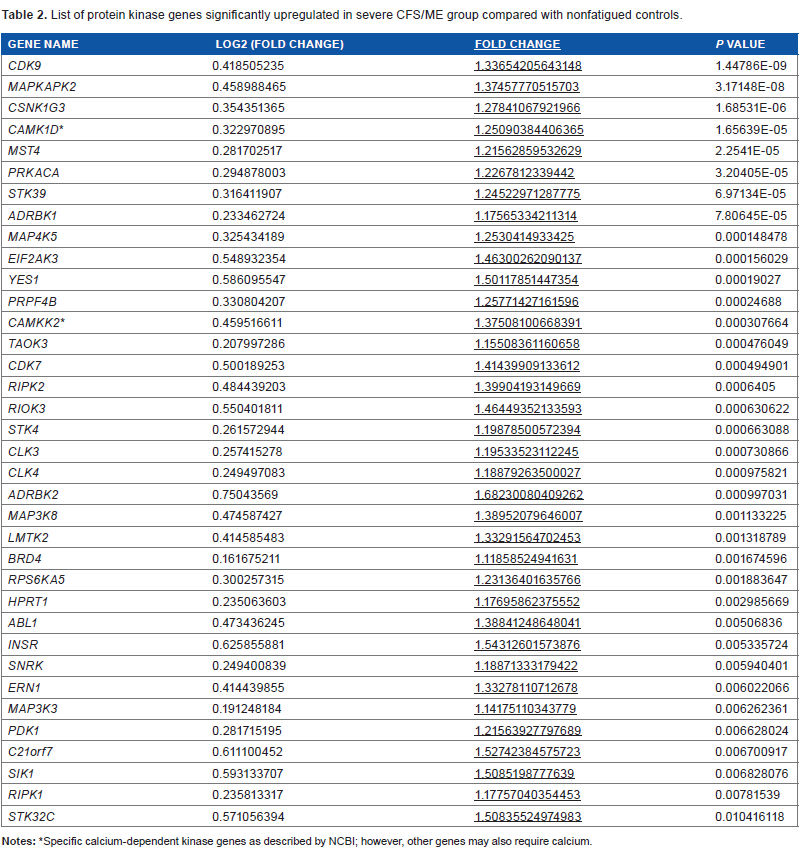
Specific calcium-dependent kinase genes as described by NCBI; however, other genes may also require calcium.
List of protein kinase genes significantly downregulated in severe CFS/ME group compared with nonfatigued controls.

Specific calcium-dependent kinase genes as described by NCBI; however, other genes may also require calcium.
A total of 92 protein kinase genes associated with severe CFS/ME patients were analyzed using MetaCore pathway analysis. The gene signatures were associated with 77 significant process networks of immune cells, immune cell function, physiological processes, signal transduction, and translation in CFS/ME patients (Supplementary Table 1).
Discussion
This study reports, for the first time, the differential expression of protein kinase genes from isolated NK cells in CFS/ME patients compared with nonfatigued controls. In particular, 37 genes were significantly upregulated and 55 genes were significantly downregulated in severe CFS/ME patients compared with nonfatigued controls. As this investigation was undertaken in isolated NK cells, the protein kinase genes that are reported will be discussed in the context of intracellular pathways involved in JNK, STAT, and NF-kappa beta (NF-κβ) activity, and their role in Ca2+ regulation and NK cell lysis.
The results from the current investigation highlight significant downregulation of Ca2+-dependent protein kinases, namely, Lck and ZAP-70. NK cells contain a zeta chain, associated with the Fc receptor CD16 (FcgRIIIA), where Zap-70 phosphorylates, as well as the associated transducing gamma chain. 32 The cytoplasmic tails of adhesion molecules and activating receptors of the NK cells recruit Src family of kinases to phosphorylate immunoreceptor tyrosine-based activation motifs.33–36 Subsequently, the signaling molecules including Lck, Zap-70, linker activation for T-cells, and SH2 domain-containing leukocyte protein of 76 kDa (SLP-76) are phosphorylated. This mechanism continues to phosphorylate and mobilize multiple downstream signaling proteins, which results in the activation of NK cells and the initiation of granule-dependent exocytosis.37–39 Given the significant reduction of both ZAP-70 and Lck, these being Ca2+-dependent protein kinases, the intracellular downstream effect may be significant for effector functions of NK cells. We and others have previously described significant reduction in NK lysis, changes in cytokine production and mobilization, and redistribution of cytoplasmic perforin and granzyme B toward the contact zone with target cells in CFS/ME patients.13,17,19,40–43
The significant reduction in ERK1/2 and MEK1/2 reported in this study aligns to our previous investigation that reported a significant decrease in ERK1/2 in CD56dimCD16+ NK cells compared with nonfatigued controls. 44 A significant reduction in MEK1/2 suggests further compromise in effector NK cell function. Following the activation of triggers, a signaling cascade via sequential phosphorylation of MAPK, MEK, and ERK results in the lytic granule polarization mediated by TUBB, 45 which regulates the reorientation of the microtubule and microtubule-organizing centre (MTOC) toward target cells to release perforin and granzymes. Activation of ERK1/2 facilitates polarization of cytotoxic granules toward the MTOC.46,47 MAPK intracellular signals activate reorganization and polarization of the actin cytoskeleton, which facilitates movement of the cytotoxic granules along the MTOC microtubules toward the immune synapse.46,47 A critical threshold for signaling of MAPK, MEK, and ERK is required for NK cells to mount an effector cell response. A significant reduction in the expression of MEK2, ERK1, and TUBB may disrupt these distal events that contribute to reduced NK cytotoxicity. This reduced NK cytotoxicity may be particularly due to a reduction in the ERK1/2 phosphorylation, reducing the polarization of the secretory granule toward the immune synapse for degranulation.13,17 We and other researchers have also observed a significant reduction in lytic granules, such as granzyme B in CFS/ME patients.13,17,19,40–43
Binding of NK cells to target cells triggers phosphati-dylinositol (PI)-3 kinase (PI3 K) to be rapidly activated by Src family tyrosine kinases and/or SYK, leading to calcium influx 48 and protein kinase C (PKC) activation. In this investigation, we report significant increases in PKC-alpha in the severe CFS/ME group compared with the nonfatigued control group. PKC-alpha is a member of PKC family of Ca2+ and/or lipidactivated serine/threonine kinases, and functions downstream in many membrane-associated signal transduction pathways. 49 For example, PKC-alpha activation triggers a signaling cascade via sequential phosphorylation of MAPK, MEK, ERK, and JNK pathways. Ca2+ ions, magnesium ions, and diacylglycerols (DAGs) are the most important molecules for regulating PKC-alpha activity, as low concentrations of these molecules increase the PKC-alpha activity. Hence, the present study highlights the importance of Ca2+ transport ion channels.
Moreover, JNK is activated by PKC-alpha, where JNK modifies the activity of numerous proteins located in the mitochondria or activates inflammation and pro-inflammatory cytokines such as IL-2, IL-6, and TNF-α. Increased activation of PKC-alpha may provide possible explanation for the increase in JNK along p38, resulting in pro-inflammatory cytokine production such as IFN-γ, TNF-α, IL-2, and IL-6, from NK cells. 50 The significant increase in PKC-alpha may suggest a shift toward a Th1/pro-inflammatory immune response. Previous researchers report significant increases in IFN-γ, IL-2, TNF-α, and IL-6 in CFS/ME patients.51–57 Moreover, anti-inflammatory IL-10 exerts inhibitory effects on cytokine secretion and impedes pro-inflammatory cytokine secretion by multiple cells including NK cells (IFN-γ and TNF-α). 14 A decrease in IL-10 favors an increase in pro-inflammatory responses, and this may increase the prevalence of Th1-like cytokines. Importantly, we and others have reported significant reductions in IL-10 from CFS/ME patients.58,59
CFS/ME is a heterogeneous condition in regard to symptoms and severity; hence, this study stratified findings according to moderate and severe patient groups. No difference in expression of protein kinase genes was observed between moderate CFS/ME patients and nonfatigued controls. This lack of observable difference in gene expression between moderate CFS/ME patients and nonfatigued controls may be due to clinical variability among this patient group. By comparison, salient changes in pathology may be predominant in a clinically severe, homogenous patient group, as observed in the findings of this study. The kinase genes identified in this study control a large number of process networks within cells affecting synaptic function, signal transduction, inflammation pathways, apoptosis, muscle contraction, microtubule cytoskeleton spindle assembly, circadian rhythm, calcium transport, and nitric oxide signaling. Metabolic effects, predominantly insulin gene expression pathways, were identified. Protein phosphorylation and protein modification pathways predominated in gene association analysis. Thus, this study revealed that multiple gene, metabolic and signaling pathway perturbations manifest in calcium-sensitive kinase genes. Kinase pathways control or regulate numerous physiologies including cardiovascular, urogenital, gastrointestinal, neurological, and respiratory systems. Kinase perturbations suggest the likely demonstration of an inflammatory profile along with other dysregulated physiological mechanisms, adding to widespread inflammatory mechanism dysregulation in virtually all cells. 60 Furthermore, Ca2+ dysregulation is an important consequence of altered membrane receptor signaling and likely to have effects in neuronal function, such as impulse transmission, 61 as well as muscle contraction. 62 Impaired neurological and motor control are common symptoms associated with CFS/ME. 25 Therefore, it is suggested that Ca2+ and kinase signaling dysregulation be further investigated in the central nervous system, given the high dependence on Ca2+ signaling for glial and neuronal cell functioning and their potential role in the pathomechanism of CFS/ME.
Conclusion
This study identifies, for the first time, 92 protein kinase genes differentially regulated in NK cells of CFS/ME patients compared with healthy nonfatigued controls. Specifically, 37 genes were upregulated and 55 genes were downregulated, which are involved in numerous cell signaling and metabolic pathways including inflammation. While primarily indicating functional impairment in NK cytotoxic activity and immunological dysfunction, kinases are located throughout cells in the body and may be associated with other clinical manifestations reported in CFS/ME.
Author Contributions
Conceived and designed the experiments: AC, SM-G. Analyzed the data: AC. Wrote the first draft of the manuscript: AC. Contributed to the writing of the manuscript: AC, DS, SJ, SM-G. Agree with manuscript results and conclusions: AC, DS, SJ, SM-G. Jointly developed the structure and arguments for the paper: AC, DS, SJ, SM-G. Made critical revisions and approved final version: DS, SJ, SM-G. All authors reviewed and approved of the final manuscript.