
Abstract
In the evolutionary process, the random transmission and mutation of genes provide biological diversities for natural selection. In order to preserve functional phenotypes between generations, gene networks need to evolve robustly under the influence of random perturbations. Therefore, the robustness of the phenotype, in the evolutionary process, exerts a selection force on gene networks to keep network functions. However, gene networks need to adjust, by variations in genetic content, to generate phenotypes for new challenges in the network's evolution, ie, the evolvability. Hence, there should be some interplay between the evolvability and network robustness in evolutionary gene networks. In this study, the interplay between the evolvability and network robustness of a gene network and a biochemical network is discussed from a nonlinear stochastic system point of view. It was found that if the genetic robustness plus environmental robustness is less than the network robustness, the phenotype of the biological network is robust in evolution. The tradeoff between the genetic robustness and environmental robustness in evolution is discussed from the stochastic stability robustness and sensitivity of the nonlinear stochastic biological network, which may be relevant to the statistical tradeoff between bias and variance, the so-called bias/variance dilemma. Further, the tradeoff could be considered as an antagonistic pleiotropic action of a gene network and discussed from the systems biology perspective.
Keywords
Background
For a long time, biologists have known that many biological functions and diseases cannot be explained by the function of an individual gene or protein. Instead, they are the result of an interactive network between a protein and another protein, or protein with other molecules.1,2 It has also been found that some particular characteristics of biological systems evolve under robust regulation, despite large changes in the environment or alternations to the internal parameters of the system.3,4 Robustness is a ubiquitously observed property of biological systems, at many levels, from biochemical to ecological systems.5–9 For example, a genome may be robust, because it encodes proofreading and repairing systems that can reduce replication errors; a metabolic function may be robust, when it encounters stresses, like a temperature change, because some enzymes can maintain their shapes and specificity, or the interconnected network of their reactions can sustain the supply of product, even when they fail; an ecosystem might be robust, if it resists the extinction of some species because of the compensatory effect of other surviving species. 10 In the genetic robustness, since the random variations, due to mutation, deletion, duplication, insertion, inversion, translation of a chromosome in germ cell or the random transmissions of genes from parent to offspring, arise constantly in the evolutionary process, it implies that evolution, as a genetic process, must allow for this randomness, because it provides the raw material for evolutionary adaptation. The mechanisms of genetic robustness, such as genetic redundancy, proofreading and repairing systems, are considered to be the fundamental features that balance the randomness of a complex evolvable gene network. 10 However, too much robustness is not conductive to a biological system to adapt to environmental changes, which play an important role in the natural selection.
Robustness, undoubtedly, is the result of an organism's ability to adapt to the environmental changes, through long-term evolution. In individual level, robustness has important implications for both disease and evolution, as it reduces the phenotypic expression of genetic changes. For evolutionary biologist, the concept of phenotype robustness (or phenotypic stability, sometimes referred to as ‘canalization’) 11 is viewed as genetic buffering that has evolved under natural selection in order to stabilize the phenotype. 12 Concerning the interplay between the evolvability and robustness, some important issues exist and claim that robustness can either increase or decrease the evolvability.10,13 On one hand, high robustness implies low production of heritable phenotypic variation. On the other hand, both experimental and computational analyses of neutral networks indicate that robustness enhances evolvability. There should exist some delicate balance between the evolvability and network robustness, in an evolutionary biological network. The paradoxical tension between genetic robustness and evolvability has been confirmed by studying RNA genotypes and their secondary structure phenotypes. 14 A high level of correlation between genetic robustness and environmental robustness was found from the biophysics of RNA folding that the genetic robustness observed in miRNAis considered as the byproduct of selection for environmental robustness. 15 Based on the yeast gene deletion collection, the effects of mutations on environmental robustness,16,17 genetic robustness18,19 and phenotypic robustness20,21 can be measured and compared on a genome-wide scale. Using global quantitative genetic datasets in yeast, 22 the phenomenon, that the coupled genetic and environmental robustness are both also coupled to stochastic robustness, are found and largely consistent with Waddington's intuition, ie, environmental change, phenotypic variation and genetic mutation have similar effects on an organism via the same underlying molecular processes. These brilliant research results provide the firm bases to make the global quantification of robustness become possible by recent genomic experiments. However, there still lack some appropriate mathematical methods to clarify the ambiguous relationship among these different types of robustness and the interplay between the evolvability and robustness.
It has been previously reported that robustness of biological systems may come from a precise network structure. 5 To understand the interplay between the evolvability and network robustness of biological systems, in the evolutionary process, it is necessary to integrate the information of genome sequences, mRNA expression, proteomes and so on, from the system level point of view, and to analyze the composition of biological systems, at various levels- the interactions between modules, stochastic system dynamics, underlying stability robustness and sensitivity in control theory23,24 and natural selection in evolution.7,25 In the past, most molecular biologists and biochemists assumed that biological evolution occurs, due to random variations in genes and natural selection. However, the evolution of biological networks via natural selection is still in the conceptual phase, with no dynamic system rules.8,9,25 The study of the evolvability and robustness of gene networks, or biochemical networks is still in its infancy. In fact, robustness allows changes in the structure and components of gene networks, due to intrinsic random fluctuations, while still allowing the specified network function to be maintained. Evolvability also allows some extrinsic random disturbances to influence gene networks, so that a new phenotype of the gene network may evolve by natural selection, to adapt to the new environment. 9
In this study, the interplay between the evolvability and robustness of a biological network in evolution, is considered from a systems perspective. According to the hierarchical network interplaying in Figure 1, 8 the high-level selected network will specify a selection force on lower levels. The external environment molds the evolution of the local adaptation of organisms by natural selection. Once the favored organisms are selected, the low-level biological networks have to maintain the favored physiological systems of the selected organisms. Hence, these favored organisms become selection forces, able to select their favored physiological systems.26–28 The favored physiological systems exert a selection force on the biochemical networks. The favored biochemical networks then become the selection forces for their regulatory gene networks. Finally, the low-level selected networks send feedback to influence the evolution of the high-level network. In this way, natural selection actually acts on the interplaying of multiple biological networks. The development of robust adaptive design rules, for the natural selection of biological networks, might allow more insight into the evolutionary mechanisms of biological networks, under the influence of random intrinsic noise and environmental disturbances. 5

The natural selection process on the interplaying of hierarchical biological networks. The high-level biological network selection will become the selection force on low-level biological network. The natural selection on organisms selects its favored organisms. Once the favored organisms are selected, the low-level biological networks have to maintain the favored physiological systems of the selected organisms. Hence, these favored organisms become the selection force to shape their favored physiological systems. The favored physiological systems will lead to the selection force on biochemical networks. The favored biochemical networks by natural selection will become the selection force on genetic networks. On the other hand, the lower-level selected networks will feedback to influence the higher-level networks in evolution. Therefore, the natural selection actually acts on the interplaying of the multiple bio-networks. 8
One of the most important features of biology is that the organism's phenotypes can be robust, against either heritable genetic variations or non-heritable environmental disturbances. In the evolutionary process, an evolving gene network can be represented by a nonlinear stochastic system that is subject to heritable random genetic variations and non-heritable random environmental disturbances. The phenotype of a gene network can be represented by the equilibrium point of a nonlinear stochastic system. There are many variations of phenotypes around this equilibrium point, due to intrinsic random genetic variations and extrinsic environmental disturbances.8,9,29–32 When a variation of a phenotype is perturbed too much, from the domain of its equilibrium point into the domains of other equilibrium points, a new phenotype is generated and preserved, if it is suited to natural selection, otherwise the variation of the phenotype is terminated. A gene locus that does not help an organism to cope with the environmental stress, or to increase its fitness is neutral. These genetically varied phenotypes are heterogeneous and allow evolvability, to cope with environmental changes. 9 In evolution, a robust gene network can harbor a large number of neutral genetic variations, before the phenotype transforms to another equilibrium point. The capacity to harbor these neutral genetic variations is called the genetic robustness of the phenotype in evolution. Therefore, the phenotype robustness of a stochastic gene network at the equilibrium point (phenotype), under the influence of parametric fluctuations, due to genetic variations, will be discussed from the point of view of stochastic Lyapunov stability. 29 Furthermore, the evolvability in response to environmental changes and the genetic variations of a stochastic gene network, in the evolutionary process, can be also discussed from the point of view of stochastic H∞ filtering. 33 In this study, both the evolvability and network robustness of a nonlinear stochastic gene network, in the evolutionary process, are discussed from a nonlinear stochastic system perspective. The tradeoff between the genetic robustness and environmental robustness is also investigated. It is found that, if a gene network has enough network robustness to cover both genetic robustness and environmental robustness, to buffer the effect of genetic variations and to resist environmental stimuli respectively, then the phenotype of the gene network is maintained, in the evolutionary process. However, the accumulation of these neutral genetic variations may destroy the network robustness and provide new material for evolution, so that a phenotype transition may eventually occur by a shift to another equilibrium point, with the help of environmental stimuli. However, any study of the interplay between the evolvability, to cope with environmental disturbances, and network robustness, to allow intrinsic genetic variations of the phenotype of the gene network, must deal with a very difficult Hamilton-Jacobi inequality (HJI). At present, there exists no method to efficiently solve the HJI analytically or numerically.
This study uses a global linearization method 34 to interpolate several linearized stochastic systems, at different operation points, to approximate the nonlinear stochastic gene network, so that the interplay between the evolvability and network robustness can be discussed from a systematic viewpoint, using a set of linear matrix inequalities (LMIs), instead of the HJI. Using this method, the evolvability and network robustness are related to the locations of the eigenvalues of the linearized stochastic systems of nonlinear stochastic gene networks. If the eigenvalues of these linearized stochastic systems of a nonlinear stochastic gene network are all located in the far left hand side of the s-complex domain, then the nonlinear stochastic gene network is more robustly stable but less adaptable, in the evolutionary process. On the other hand, if these eigenvalues are closer to the imaginary axis in the s-complex domain, and are more easily perturbed to the right hand side, then the nonlinear stochastic gene network is less robustly stable but more adaptable, in the evolutionary process. The quantitative measures of the evolvability and network robustness are also investigated, from this systematic perspective. If the capacity of the neutral genetic variations tolerated by a gene network is denoted as the genetic robustness, and the evolvability is considered as the inversion of the environmental robustness, then it is also found that the phenotype of the gene network is robust in evolution, if the genetic robustness plus the environmental robustness is less than the network robustness of a gene network under the influence of genetic variation and environmental disturbance, in the evolutionary process. The correlation between the genetic robustness and environmental robustness of a biological network in evolution,15,35 can also be confirmed by the proposed network robustness, in the evolutionary process.
Since biochemical networks play the central role in cellular function, there is strong motivation to investigate the interplay between the evolvability and network robustness of evolutionary biochemical networks. As shown in Figure 1, the favored physiological systems exert a selective force on biochemical networks, in the evolutionary process. The varied phenotypes of the biochemical networks result from random changes in protein sequences, protein structure and promiscuous protein functions, due to the corresponding genetic variations, in the evolutionary process. 9 Since several biochemical networks are the backbone of physiological systems, they should have enough network robustness to tolerate intrinsic parametric variation and less evolvability, in response to external stimuli,36,37 which means that the steady state of the system variables (concentrations or rates) is invariant, under perturbations. In this study, based on the S-system model, 36 the evolvability and the network robustness of nonlinear stochastic biochemical networks are measured from the steady state perspective. A formula is also derived, to define their interplay, in order to discuss their relationship, in the evolutionary process. Finally, the interplay between the evolvability and network robustness of high-level biological networks, for example, physiological systems and bio-ecological systems, is also discussed in the same vein. The interplay between the evolvability and network robustness of a biological network in evolution is related to evolutionary antagonistic pleiotropy. 38 Based on the evolutionary theory of aging, 38 antagonistic pleiotropy holds that what is good for an organism when it is young can be bad for it when it is old. This study discusses some evolutionary antagonistic pleiotropic actions of a biological network, from the viewpoint of the tradeoff between the genetic robustness and environmental robustness in evolution. In this situation, the evolutionary antagonistic pleiotropic effect is extended, from the individual gene level to biological network level, for the evolutionary process. Furthermore, based on stability robustness,23,39 noise filtering theory33,40 and systems biology,5,41 the interplay between the evolvability and network robustness in evolution, can be applied to a synthetic gene network to model its progress, using a fast evolutionary computation method (eg, genetic algorithm and evolution algorithm) to select adequate circuit components for a synthetic gene network, in order to achieve a desired behavior (ie, robustness and filtering ability).42–49
In the following, the interplay between the evolvability and network robustness in a linear gene network is firstly discussed from the point of view of systems biology and then the results are extended to the evolutionary nonlinear gene network.
On the Interplay between the Evolvability and Network Robustness of Linear Stochastic Gene Regulatory Network
Network robustness of linear gene regulatory networks in evolution
For initial simplicity of analysis, we consider a linear dynamic gene regulatory network as follows (see Fig. 2).

where x(t) = [x1(t) x2(t) … x n (t)] T denotes the gene expressions of n genes in the network and

denotes the regulatory interaction matrix between these genes, for example a ij denotes the regulatory interaction from gene j to gene i.
Remark 1
The linear gene regulatory system in (1) can be considered as a linearized system of a nonlinear gene network
In the evolutionary process, the interaction a ij may suffer from random parametric fluctuations, due to heritable DNA mutation, genetic variations and recombinations in the germ cell (see Fig. 2).
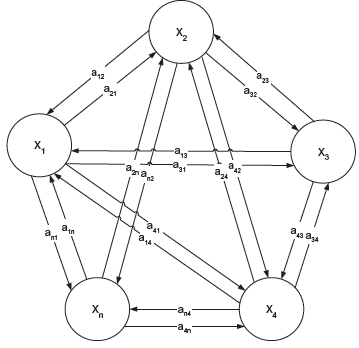
A linear n-gene regulatory network. a ij denotes the interaction from gene j to gene i and x i (t) denotes the gene expression of gene i.

where

In this equation, δ(y,τ) denotes the delta function, ie, δ(t,τ) = 1, if t = τ and δ(t,τ) = 0, if t ≠ &tau. In other words,
Suppose the gene regulatory network in (1) suffers from the following genetic parameter fluctuations in evolution

where
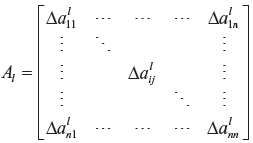
denotes the parametric fluctuations, due to the l-th random source of n
l
(t). If
The stochastic gene regulatory network in (4) is termed stochastically stable, if there exists a Lyapunov (power) function V(x) = x
T
(t)Px(t) for a positive definite matrix P such that the expectation of the derivative
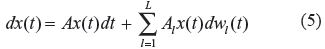
where dw l (t) = n l (t)dt and w l (t) denote the corresponding Wiener process or Brownian motion of random parametric fluctuations in evolution, ie, the stochastic system in (5) denotes the phenotypic heterogeneity in the gene network population, in evolution.
Proposition 1
The stochastic gene network in (5) is said to be robustly stable in evolution, with its phenotype x e = 0, if the following phenotype robustness inequality holds.

Proof
See
Remark 2
In the case of the nominal gene regulatory network in (1), the robust stability condition is reduced to the following stability inequality with
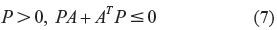
ie, the stability condition for a gene network free of parametric fluctuations in (1). The stability inequality in (7) implies that all of the eigenvalues of A should be on the left hand side of the s-domain, ie, the real parts of all eigenvalues of A should be negative or zero.
23
However, for the stochastic gene regulatory network in (5), the eigenvalues of A should be more further to the left hand side of the s-domain, so that the phenotype robustness inequality in (6) can hold, ie, the eigenvalues of A should be more negative (robust), in order to tolerate the positive term

Since the right hand side of (8) can be considered as an indication of the network robustness of the gene regulatory network, if the effects of these stochastic parametric fluctuations on the gene network, due to genetic variations, in evolution, can be overridden by the network robustness of the gene network, then they will be tolerated, without changing the pleiotropic phenotype of the gene network, in evolution. In this situation, the parametric fluctuations
Evolvability of a linear gene regulatory network in evolution
Suppose the gene regulatory network in (1) only suffers from stochastic environmental stimuli as follows.33,39

where v(t) = [v11(t) v2(t) … v
n
(t)]
T
denotes the external stimuli or molecular noises, due to environmental changes or stresses such as temperature or salinity, and B denotes the coupling matrix between external stimuli and the gene network. Environmental stimuli can perturb x(t) away from its phenotype x
e
= 0. Let us denote the phenotype variation as

The numerator in (10) denotes the total average phenotype variations to present time t p . The response r indicates the effect of environmental stimuli on the phenotype variations from a total energy point of view. The fitness function f for the phenotype x e in evolution is inversely proportional to the response r of the gene regulatory network

ie, a gene network with less (more) response to environmental stimuli exhibits more (less) fitness to the phenotype x e = 0. Therefore, the maximization of the fitness function for phenotype x e = 0 in evolution is equivalent to the minimization of the response r in (10). We solve for the maximum fitness of phenotypex e under the influence of environmental stimuli, using the following equivalent minimization problem for phenotype variation in response to environmental stimuli, in evolution

We call r0 the evolvability of the stochastic gene regulatory network in (9), in response to environmental stimuli, and the maximum fitness
However, it is still very difficult to solve the minimization problem (12) for the evolvability r0. A suboptimal method is employed to solve it, by minimizing its upper bound. Let us denote the upper bound of r in (10) as follows

where the evolution level ρ denotes the upper bound of the evolvability, ie, the upper bound of the influence of environmental stimuli, in the evolutionary process, on the phenotype variation of the gene regulatory network, from the average energy point of view. t
p
denotes the present time. If ρ in (13) is large, then the influence of environmental stimuli (or disturbances) on the phenotype variation of the gene network is large, ie, the gene network is more evolvable to the environmental stimuli and vice versa. In the following, the upper bound ρ is minimized, until its minimum value approaches the evolvability r0 in (12), ie, solving for r0 in (12) by the so-called suboptimal method.
33
If the initial
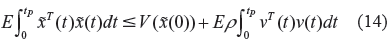
then we get the following result.
Proposition 2
For the gene regulatory network with environmental stimuli (or disturbances) in (9), in the evolutionary process, if the following inequality holds for P > 0.

then the evolutionary inequality in (13) or (14) holds, ie, the gene regulatory network in (9) has an evolution level ρ.
Proof
See
Since the evolution level ρ is the upper bound of the evolvability of the gene regulatory network, the evolvability r0 of the gene regulatory network is obtained as follows

subject to (15).
The constrained optimization problem for solving the evolvability r0 for the linear gene regulatory network in (9) can be achieved by decreasing ρ until no solution P > 0 exists in (15). By Schur complement, 34 the quadratic inequality in (15) can be transformed into the following equivalent linear matrix inequality (LMI).

Therefore, the evolvability measure in (16) is equivalent to solving the following constrained optimization problem

subject to (17).
The evolvability r0 in (18) is measured by decreasing ρ until the LMI in (17) has no solution P > 0. The solution P > 0 of the LMI in (17) is easily obtained with the help of the LMI toolbox in Matlab.
34
After obtaining r0 by solving the constrained optimization in (18), by the fact
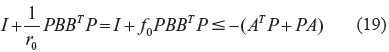
From the above, it is seen that if the network robustness –(A T P + PA) on the right hand side of (19) is larger (ie, with more negative eigenvalues of A), then the gene network has a small evolvability r0 or large environmental robustness f0 to resist external stimuli. In this situation, the gene network is less adaptive. Taken in a broad sense, f0 is related to the environmental robustness or the buffering ability against environmental stimuli, in evolution, to resist these environmental stimuli, 35 ie, the evolvability r0 is inversely proportional to the environmental robustness indicator f0. Small r0 indicates that the phenotype x e has good environmental robustness and can resist environmental stimuli and large r0 indicates that the phenotype x e has less environmental robustness to environmental stimuli. If the network robustness of the gene network is large, or r0 is small, evolutionary adaptation may be promoted, because of the increased likelihood of having a small subpopulation that can cope with the environmental stimuli. Conversely, if the network robustness of the gene regulatory network is small, the evolvability r0 must be large enough to respond to environmental stimuli, in order to guarantee the evolution inequality in (19). In this case, the population becomes increasingly heterogeneous in evolution. It can be seen that the evolvability and network robustness are two antagonistic characteristics of the gene regulatory network, in the evolutionary process. If one is large, then the other must be small and vice versa.
Tradeoff between environmental robustness to respond to environmental stimuli and genetic robustness to tolerate parametric fluctuations in evolution
If the gene regulatory network in (1) suffers from simultaneous intrinsic parametric fluctuations and environmental stimuli in evolution, then we get

Then we get the following result.
Proposition 3
If the following phenotype robustness criterion has a solution P > 0

then the stochastic gene network in (20) has enough network robustness to tolerate genetic parametric fluctuations and an evolutionary level ρ that allows it to respond to environmental stimuli, in the evolutionary process.
Proof
See
Then the evolvability of the stochastic gene network in (20) can be measured by solving the following constrained optimization problem.
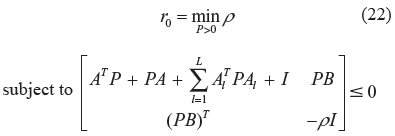
After solving r0 from (22), the phenotype robustness criterion in (21) can be modified as
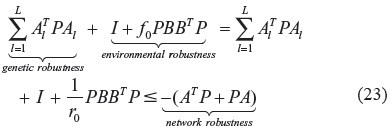
From (23), it is seen that the phenotype of the gene network can be maintained, if the network robustness can take over the genetic robustness to tolerate parametric fluctuations, and the environmental robustness to resist environmental stimuli, in evolution. The interplay between the evolvability and network robustness of a gene regulatory network in evolution, is discussed in the following.
For the gene network with only environmental stimuli v(t) in (9) and with the network robustness in (19), if the eigenvalues of A are more negative, then the gene network is more robust and should be less evolvable (small evolvability r0 or large environmental robustness indicator f0) to resist the influence of environmental disturbance v(t). However, for the gene network in (20) with the same network robustness as in (23), ie, with the same network robustness –(A
T
P + PA) as (19), the tradeoff between the evolvability and genetic robustness is discussed in the following. Since an extra genetic robustness

Due to the destruction of network robustness by parametric fluctuations, the evolvability r0 in (24) is increased, or the environmental robustness indicator f0 is decreased, in order to guarantee the above inequality, because of the diminution of the network robustness, on the right hand side, ie, the effect of parametric fluctuations, due to genetic variations in evolution, may make the gene network more responsive (or evolvable) to environmental stimuli. 50 Similarly, the inequality can be changed to

Due to the diminution of network robustness by the evolvability r0 to the response to environmental stimuli in (25) or by environmental robustness, the tolerable amount of genetic parametric fluctuations
Let us denote the tolerable value

ie, if the genetic robustness plus environmental robustness is less than the network robustness, in the evolutionary process, the phenotype of the stochastic gene network in (20) is robust in evolution.
In, 35 the evolutionary causes of genetic robustness were discussed, using different evolutionary scenarios. The correlation with environmental robustness is considered to be the most probable cause of genetic robustness in evolution. Based on the congruence scenario, the genetic robustness of a gene network in evolution is a by-product of the environmental robustness of a gene network to resist the environmental stimuli (or perturbations), because environmental stimuli (perturbations) are more frequent than the genetic perturbations in evolution, which have been confirmed in RNA folding and heat-shock proteins.15,35 This correlation between the genetic robustness and environmental robustness is obvious in (23) and (26). In order to provide a buffer against environmental disturbances, the gene network may develop feedback, hub and redundant mechanisms, to improve its network robustness, resulting in a large –(A T P + PA). This greater network robustness can also provide a buffer against large amounts of neutral genetic variations simultaneously in evolution. However, this large amount of genetic variations may provide raw material for new evolutionary possibilities. Therefore, the phenotype robustness criterion in (23), gives more insight into the interplay between the evolvability and network robustness.
On the Interplay between the Evolvability and Network Robustness of a Nonlinear Stochastic Gene Regulatory Network in Evolution
In real biological systems, the gene regulatory networks are always nonlinear. In the case of a nonlinear gene regulatory, the dynamic system in (1) is modified as

where f(x) denotes the nonlinear interactive function vector among these regulatory genes in the gene network.
Network robustness of a nonlinear gene regulatory network in evolution
For a nonlinear stochastic gene regulatory network with stochastic parametric fluctuations in evolution, the stochastic system in (5) is modified as follows29,42

where f l (x) denotes the nonlinear stochastic parametric fluctuation, due to the l-th random genetic fluctuation source w l (t), in the evolutionary process. Sources of these random genetic fluctuations, in the evolutionary process, include DNA mutation, deletion, duplication, inversion and translocation of chromosome. The nonlinear stochastic system in (28) represents the phenotypic heterogeneity in a gene network population, due to random genetic variations in evolution.
Consider the nonlinear system in (27) or (28). Many equilibrium points (phenotypes) exist (see Fig. 3). Suppose a phenotype near a stable equilibrium point x
e
is of interest. For convenience in analysis, the origin of the nonlinear stochastic gene network in (28) is shifted to the equilibrium point (phenotype) x
e
. In this case, if the shifted nonlinear stochastic system is robustly stable at the origin, then the equilibrium point (phenotype) of interest is also equally stable. Analysis of the procedure of the evolvability and network robustness of a nonlinear gene regulatory network in evolution, is thus simplified. Let us denote

The stochastic nonlinear gene network has many local stable equilibrium points (phenotypes). The landscape of three stable equilibrium points is shown with vertical scale illustrating the relative network robustness of the equilibrium point (phenotype) of the nonlinear gene network. Suppose x e is the equilibrium point (phenotype) of interest.

ie, the origin
Then, let us consider the robust stabilization of the nonlinear stochastic gene network in (29) in evolution. Based on the stochastic Lyapunov stability,33,39 the following result, for phenotype robustness, is obtained.
Proposition 4
The phenotype x
e
of the stochastic nonlinear gene network in (29) exhibits phenotype robustness in evolution, if the following Hamilton Jacobi inequality (HJI) holds for some positive function

ie, the nonlinear fluctuating gene network in (28) or (29) is robustly stable, at the equilibrium point x
e
, under the influence of random genetic fluctuations in evolution, if the above HJI holds for a positive function
Proof
See
Remark 3
(i) If a nonlinear gene regulatory network is free of parametric fluctuations, then from (30), the stability of the nonlinear gene network in (27), at the equilibrium point (phenotype), is guaranteed by
Evolvability of a nonlinear gene regulatory network in evolution
In the nonlinear gene regulatory network with only stochastic environmental stimuli, eq. (29) is modified as

Based on the nonlinear stochastic gene network in (31) and the evolution level ρ of the gene regulatory network defined in (13), the following result is obtained.
Proposition 5
For the nonlinear gene regulatory network with external stimuli in (31), if the following HJI holds for some positive function

then the evolution level ρ in (13) holds for the gene regulatory network with a phenotype at the equilibrium point x e .
Proof
See
In this case, the evolvability measure of the gene regulatory network in (31) in evolution, can be obtained as follows

subject to (32).
Remark 4
(i) Comparing (33) with (16), it is seen that the evolvability inequality in (15), for the linear gene network, is replaced by the above HJI in (32), for the nonlinear gene network, (ii) If the nonlinear gene network is more robustly stable, ie, if
Based on evolutionary aging theory in, 38 if the gene network exhibits greater environmental robustness (ie, large f0 or smaller r0 in (32)) at an early stage, resulting in a diminution of the external stimuli and genetic variations, so that the phenotype always remains in the basin of an equilibrium point in Fig. 3, then the effects of pathogen infections and genetic variation can be easily attenuated by the gene network, and disease and cancer can be efficiently suppressed, at an early stage. However, at a more mature stage, the gene network is less robust to genetic variations, or more responsive to environmental changes, so that the phenotype will be easier to transit from one equilibrium to another and aging (senescence) of the gene network will occur. Therefore, the interplay between the evolvability and network robustness is inherent to the antagonistic pleiotropic effect of the gene network, in evolutionary process.
Tradeoff between the environmental robustness and genetic robustness of a gene regulatory network in the evolutionary process
If the nonlinear gene regulatory network (27) suffers from simultaneous parametric fluctuations and environmental stimuli in evolution, the result is the following nonlinear stochastic system

near the equilibrium point (phenotype) xe.
Then we get the following result.
Proposition 6
If the following HJI has a positive solution
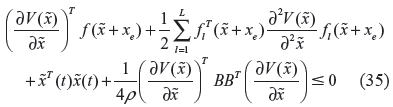
then the nonlinear stochastic gene regulatory network in (34) has enough network robustness to tolerate genetic parameter fluctuations and a sufficient an evolution level ρ to respond to environmental stimuli, in the evolutionary process.
Proof
See
Based on the HJI in (35), the evolvability of the nonlinear stochastic gene network, under the influence of both parameteric fluctuations and environmental stimuli, can be measured by solving the following constrained optimization

subject to (35).
If we substitute r0 into (35), the following HJI is obtained, for phenotype robustness in evolution.
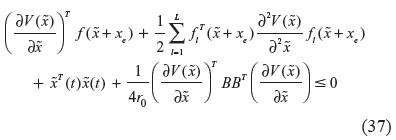
Obviously, from (35), it is seen that a nonlinear gene network requires more network robustness in (37), than in (30) or (32), when it suffers from simultaneous genetic parametric fluctuations and environmental stimuli in evolution, ie,
If a gene regulatory network selects a strategy with enough genetic variation tolerance and stimuli resistance,
50
the basin of the equilibrium x
e
in Figure 3 must be deeper and wider, so that the genetic variations and environmental stimuli are inhibited and the phenotype can be securely retained within the basin. In this case, the term
When the phenotype robustness inequality (37) holds, the accumulated genetic variations
In general, it is still very difficult to solve the HJI in (30) directly, in order to determine the extent to which stochastic parametric fluctuations can be tolerated by the gene network, or to solve the unconstrained optimization in (33) or (36), in order to measure the evolvability of the gene network, to gain a greater insight into the dynamic mechanism of a nonlinear stochastic gene network, in the evolutionary process. At present, there is no good method to solve the nonlinear partial differential HJI either analytically or numerically.33,39,40 In this case, the global linearization technique34,40 is used, to transform the nonlinear stochastic gene network (34) into an interpolation of a set of locally linearized gene networks, in order to simplify the analysis of the evolvability and network robustness of a stochastic gene network in evolution.
Using the global linearization method, if all the global linearizations are bounded by a polytope, consisting of M vertices, as34,40

where Co denotes the convex hull of the polytope with M vertices, defined in (38), ie, if the linearized systems at all
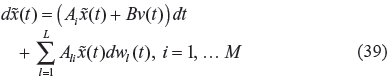
Based on the global linearization theory, 34 if (38) holds, then every trajectory of the nonlinear stochastic gene network in (31) can be represented by a convex combination of the M linearized gene networks in (39). Therefore, if we can prove that the convex combination of the M linearized gene networks in (39) has the evolvability r0 to respond to environmental stimuli and the network robustness to tolerate stochastic parametric fluctuations, due to genetic variations, then the original nonlinear stochastic gene network in (34) will also have the same evolvability and network robustness. The convex combination of the M linearized gene networks in (39) can be represented by34,40

where the interpolation function
In the case of free of environmental stimuli, the global linearization approach in (40) is reduced to

which is the global linearization system for a nonlinear stochastic system (28). For the global linearization stochastic system in (41) with only stochastic parametric fluctuations, due to genetic variations in evolution, we get the following result.
Proposition 7
If the following linear matrix inequalities (LMIs) have a positive symmetrical solution P > 0.

then the stochastic parametric fluctuations in (41) or (28) can be tolerated by the stochastic gene network in the evolution process, ie, the nonlinear stochastic gene network is robustly stable for the phenotype (ie, with phenotype robustness) at x e (the equilibrium point of interest) under the influence of parametric fluctuations in evolution.
Proof
See
Actually, the LMIs in (42) can be considered as a set of linearizations of the HJI in (30), ie, we could replace a nonlinear HJI by a set of LMIs, using the global linearization technique. If the stochastic gene network in (40) is free of genetic parametric fluctuations and is subject to only environmental stimuli in evolutionary process, then we get

which is the global linearization system of the nonlinear gene network in (31).
In this case, the evolutionary inequality in
Proposition 8
If the following evolutionary inequality holds with P > 0 for a nonlinear gene network (31) or (43)
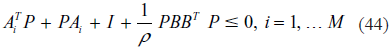
then the evolutionary inequality in (13) or (14) holds, ie, the nonlinear stochastic gene network in (31) or (43) has an evolution level ρ.
Proof
See
Since ρ in (13) or (14) is the upper bound of the evolvability for the nonlinear gene network in (31) or (43), the evolvability of a nonlinear gene network is obtained as follows

subject to (44).
The constrained optimization problem in (45), to solve the evolvability of nonlinear gene network in (31) or (43) can be achieved by decreasing ρ until no solution P > 0 exists for the quadratic inequalities in (44). By Schur complement, 34 the quadratic inequalities in (44) can be transformed to the following equivalent LMIs

Therefore, the evolvability measure in (45) is equivalent to solving the following constrained optimization problem

subject to (46).
Remark 5
(i) The LMIs in (46) or the LMI-constrained optimization problem in (47) can be efficiently solved, using the LMI toolbox in Matlab. (ii) r0 is obtained by decreasing ρ until no solution P > 0 exists for LMIs in (46) with the help of the LMI toolbox in Matlab. (iii) Based on the global linearization, the measure of the evolvability, from the HJI-constrained optimization in (33), can be replaced by the LMI-constrained optimization in (47). 52 After solving the evolvability r0 from (47), the inequalities in (44) can be modified as.

From (48), it is seen that if the eigenvlaues of A i for all linearized gene networks are more negative (ie, more robust), 23 r0 can be smaller to better resist the environmental stimuli, ie, there is a larger environmental robustness f0. In this case, the gene network tends toward stasis, under the influence of extreme environments, a phenomenon that is observed in “living fossils”. 50 Conversely, if network robustness is smaller, the evolvability r0 of a gene regulatory network must be larger, to ensure a response to environmental stimuli, in order to guarantee the inequality in (48), ie, there will be a smaller environmental robustness. This leads to the rapid evolutionary change of the gene regulatory network. Therefore, the interplay between the evolvability and network robustness of a nonlinear stochastic gene regulatory network in evolution, can be also investigated using the linear theory method. The phenotype robustness in (48) shows that if the network robustness of every local linearized gene network is larger than the environmental robustness, then the phenotype of a nonlinear gene network is robust, under the influence of environmental stimuli, in the evolutionary process.
If a nonlinear regulatory gene network is subject to genetic parametric fluctuations and environmental stimuli simultaneously, as in (34) or (40), then the following result, is obtained.
Proposition 9
If the following quadratic inequalities have a solution P > 0,
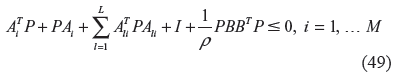
then the nonlinear stochastic gene network in (34) or (40) has sufficient network robustness to tolerate random genetic parametric fluctuations and with an evolution level ρ sufficient to respond to environmental stimuli in the evolutionary process.
Proof
See
Based on

subject to
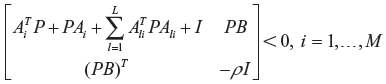
After solving the evolvability r0 from (50), then the phenotype robustness inequality in (49) can be modified as.

In (51),
From (51), it can be seen that the tradeoff between the environmental robustness to resist the environmental stimuli, and genetic robustness to resist the stochastic parametric fluctuations in evolution. A more detailed discussion of the interplay between the evolvability and network robustness of the nonlinear stochastic gene network in (34) or (40) is undertaken in the following. Firstly, the phenotype robustness inequalities in (51) can be changed to the following equivalent inequalities.

Compared with (48), the term

Due to adaptation to environmental stimuli, the network robustness of each linearized gene network is diminished, on the right hand side of (53), so that its tolerance of genetic parametric fluctuations is decreased, in the evolutionary process, ie, the tolerance to
On the Interplay between the Evolvability and Robustness of Biochemical Networks in Evolution
Since the biochemical network plays the central role in cellular function, there is strong motivation to study for the underlying principles of the adaptive evolution of biochemical networks. As seen in Figure 1, the favored physiological systems will lead to the exertion of a selection force on biochemical networks, in the evolutionary process. Since the biochemical networks are the backbone of physiological systems, they have less evolvability to allow them to respond to external stimuli and enough network robustness to resist parametric fluctuations, so that the favored physiological functions can be preserved, in the evolutionary process.36,37 In order to test whether a physiological function would prevails, in a new environment, the network robustness and the sensitivity measure the ability to tolerate the variations in the metabolite concentrations of a biochemical network, in the face of environmental changes. 6 The mechanisms of the selection forces in biochemical networks, in the evolutionary process, were also discussed in. 49
In this study, the interplay between the evolvability and network robustness of a biochemical network will be discussed, based on the S-system model. 36 Actually, it can be extended to other dynamic models of biochemical networks. Using the S-system model, the dynamic system of a biochemical network, in the evolutionary process, can be represented by 36
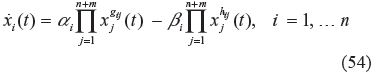
where x1(t), … x n+m (t) are metabolites, such as substrates, enzymes, factors or products of a biochemical network in which x1)(t),…x n (t) denote the n-dependent variables (intermediate metabolites and products), and xn+1(t), … x n+m (t) denote the m-independent variables (initial reactants and enzymes), α i and β i denote the rate constants, and g ij and h ij represent the kinetic parameters of the biochemical network. These parameters can be estimated using experimental data or microarray data.53,54 Suppose that transient time is neglected, in the evolutionary process and, for simplicity, we focus on the evolvability and network robustness of a biochemical network in the steady state near the equilibrium point of interest (ie, the phenotype of the biochemical network). Consider the steady state of the biochemical network in (54), we get
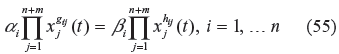
Taking the logarithm of both sides of (55), and introducing new variables y j (t) = ln[x j (t)],a ij =g ij –h ij and b i = ln(β i /α), after some rearrangement, we get 36

The above equations can be represented by the following steady state equation, 55

where Y D (t) = [y1(t) … y n (t)] T , b = [b1 … b n ] T , Y I (t) = [yn+1(t) … ym+1(t)] T ,
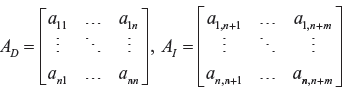
In which A D denotes the system matrix of the catalytic interactions between the dependent variables y1(t) … y n (t) in Y D (t) and A I indicates the catalytic interactions between dependent variables y1(t) … y n (t) in Y D (t) and the independent variables Yn+1(t) … Y n+m (t) in Y I (t) (ie, the environmental medium of the metabolic system). From the simple algebraic steady state equation in (57), obviously, the S-system in (54) is a useful model to describe the phenotype of the biochemical network. 36 If the inverse of A D exists, the steady state (or phenotype) of the biochemical network is solved by. 36
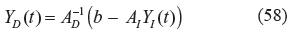
The steady state (phenotype) Y D (t) in (58) is one of the equilibrium points of the nonlinear biochemical network in (54). Actually, there are many equilibrium points for (54), which represent different phenotypes. Only the equilibrium point (or phenotype) in (58) is favored by natural selection in evolution.
Suppose that there exist some random parametric variations Δα i (t), Δβ i (t), Δg ij (t), Δh ij (t), due to random genetic mutations in protein sequences, structure and function, and ΔY I (t), due to environmental random disturbances, in the evolutionary process, ie,

These random parametric variations, due to phenotypic noise and genetic variations, can be considered as the design parameters of the biochemical network, in the evolutionary process. Then the corresponding heterogeneous steady state (phenotypic heterogeneity) of the biochemical network in evolution is given by

Because biochemical networks are the backbone of the physiological systems of organisms, a biochemical network must be sufficiently robust to tolerate the random parametric variations and environmental changes, due to phenotypic variation and noise, in order to maintain its functional properties, in the evolutionary process (see Fig. 1). We can find the following phenotype robustness of biochemical network, in the evolutionary process.6,56
Proposition 10
If the following phenotype robustness condition holds

then the phenotype of a stochastically perturbed biochemical network exists, in the evolutionary process, as follows:
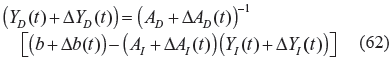
ie, if the covariance
Proof
Similar to ref 6 6 except the consideration of random variations.
If the phenotype robustness condition in (61) is violated, in the evolutionary process, the matrix inverse on the right hand side of (62) may cease to exist or move to another equilibrium point with a change of phenotype. The random variations, Δb(t) and ΔA I (t), due to the mutations of protein sequences, and ΔY I (t), due to the environmental change of the biochemical network, will influence the phenotypic variations ΔY D (t) in (62), in the evolutionary process. Their effects on the phenotype can be measured from (62) by the following evolutionary analysis of the biochemical network. 36
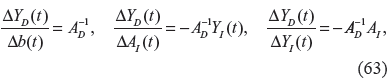
In order to tolerate random variations, Δb(t),ΔY I (t) and ΔA I (t), in the evolutionary process, to preserve the phenotype of a biochemical network, the evolvabilities in (63) must remain below some values as follows
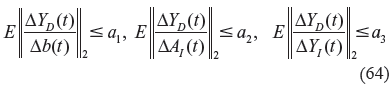
or equivalently, from (63) and (64), we get

where a1, a2 and a3 are some small evolvability values, such that the disturbed phenotype ΔY D + ΔY D (t) of the stochastic biochemical networks in (62) cannot change too much, in comparison with the nominal phenotype in (58), ie, ΔY D (t) in (62) must still be in the basin of the phenotype in evolution. The evolvability criteria in (64) or (65) determine the ranges of the sensitivities of the phenotypic changes ΔY D (t), due to random parametric and environmental fluctuations.
Based on the above analysis of the evolvability and network robustness of biochemical networks, the perturbed biochemical networks should satisfy the phenotype robustness criterion in (61), in order to prevent them being perturbed too far from their equilibrium points (phenotypes), in the evolutionary process. Because the violation of (61) generally implies a lethal phenotype mutation, the phenotype robustness criterion in (61) is the necessary condition for the survival of a phenotype of a biochemical networks, in the evolutionary process.
49
From the phenotype robustness criterion in (61), natural selection favors the perturbed biochemical networks with small-variance random phenotypic mutations, ie, with small
On the other hand, the evolvability criteria in (64) or (65) determine the range of the adaptability of the phenotypic change ΔY D (t) to random parametric variations, due to genetic and phenotypic mutations and random environmental changes, in the evolutionary process. For a functional biochemical network, the evolvability criteria should prevent the metabolic concentration from being changed, too much, by random genetic and phenotypic mutations and environmental changes. Hence, the steady state (phenotype) of a biochemical network can be preserved, while exposing the random genetic and phenotypic mutations and environmental changes to natural selection, in the evolutionary process.
When the network robustness in (61) is compared with the evolvability in (65), the interplay between evolvability and network robustness in biochemical networks is obvious in evolution. From (61), it is seen that a large
On the Interplay between the Evolvability and Network Robustness of High-Level Biological Networks in Evolution
In this section, the interplay between the evolvability and network robustness of high-level bio-networks, such as physiological systems and ecological networks (see Fig. 1) will be discussed in a similar vein. In the evolutionary process, we assume a high-level biological network can be represented by the following nonlinear stochastic system
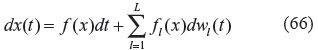
where x(t) = [x1(t),…,x
n
(t)]
T
denote n components of the high-level biological network such as organs or species, f(x) denotes the interactions between the components in the high-level biological network, the term
Suppose we only discuss the biological phenomenon near an equilibrium point x
e
, in which we are interested, ie, x(t) =

Then we get the following result.
Proposition 11
For the high-level biological network with intrinsic random fluctuations in (66), if the following HJI has a positive solution V(

then the nonlinear stochastic biological network is robustly stable around the equilibrium point in evolution, ie, the system's random fluctuation can be tolerated by the biological network, in the evolutionary process.
Proof
Similar to that for
If the stochastic high-level biological network in (67) is also subject to environmental disturbance, in the evolutionary process, as follows
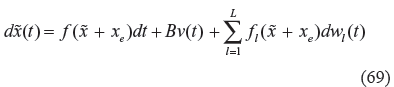
where v(t) denotes the environmental disturbances to the high-level biological network, such as impacts from environmental hormones or climatic anomalies, then we get the following result.
Proposition 12
For the nonlinear stochastic biological network in (69), under the influence of random parametric fluctuations and environmental disturbances in evolution, if the following HJI has a positive solution, V(

then the nonlinear stochastic biological network can tolerate the intrinsic random fluctuations and also has a sufficient evolution level ρ to respond to environmental disturbances, in the evolutionary process.
Proof
Similar to Proposition 6.
The evolvability of the nonlinear stochastic high-level biological network in (69) can also be measured, using the following constrained optimization method

subject to HJI in (70).
Since it is very difficult to solve the HJI in (70), or to solve the HJI-constrained optimization in (71), for the evolvability and network robustness problem of the nonlinear stochastic high-level biological networks in (69), global linearization techniques can be also employed as with
The interplay between the evolvability and network robustness of a biological network is related to the evolutionary antagonistic pleiotropy, 38 which can extend the antagonistic pleiotropy from the gene level to the biological network level, so that the antagonistic pleiotropy can be explained from a systematic perspective. For example, based on the evolutionary theory of aging,35,38 a biological network can more robustly attenuate the effect of genetic variations and extrinsic pathogens to efficiently depress cancer, at an early stage, but the biological network must be less adaptive in its response to other environmental stresses, in old age, so that senescence occurs. 38
Discussion and Conclusion
In the evolutionary process of stochastic biological systems, phenotypic, environmental and genetic variations are systematically correlated, which accounts for their evolvability and network robustness. The robustness of phenotypes evolves in response to the need to cope with stochastic genetic and environmental variations. Native functions or phenotypes that play a permanent physiological role and have therefore been constantly under natural selection, must become more robust with respect to genetic, phenotypic and environmental variations. In contrast, latent, promiscuous, or coincidental phenotypes are easily perturbed by stochastic genetic, phenotypic and environmental variations, in evolution, and thereby provide the raw material for evolutionary novelty. 61
Here, the living organism was considered as a robustness system under stochastic perturbations during the evolution. Therefore, the interplay of robustness and evolvability can be analyzed by using the well-developed system theories. From the system perspective, the phenotype robustness of the organism's system in evolution can be seen as the network robustness needs to obey a phenotype robustness criterion (23) or (26), ie, “genetic robustness + environmental robustness ≤ network robustness”, which means the network robustness needs to be strong enough to tolerate either heritable perturbations (ie, intrinsic parameter fluctuations) or non-heritable perturbations (ie, external environmental disturbances) between generations so that phenotype of the network can be maintained in the evolutionary process. For the reason that the phenotype robustness of a stochastic gene regulatory network is completely consistent with the idea of Waddington. 11 Based on the criterion, the correlation among genetic robustness, environmental robustness and the phenotype robustness by recent genomic experiments in yeast, ie, genes confer similar robustness to genetic, environmental and phenotypic robustness, 22 can be rationally explained. That is if the network robustness of a gene network is large enough then the genetic perturbations or environmental disturbances can be took over respectively or simultaneously to maintain the functional phenotype during the evolutionary process. The environmental robustness indicator f0 reversely proportioned to the evolvability r0 can also largely illustrate the antagonistic relationship between the robustness and evolvability found in the RNA secondary structure phenotypes.10,14 Further, due to the hierarchical structure in biological networks, these results may not only suit for genetic network but also suit for high-level biological network in metabolic pathway or ecological system during evolution.62,64
The dynamics of evolutionary change at molecular level, ie, the various molecular mechanisms in the long-term evolution of genomes, genes and their products, have been studied extensively.61,65 In this study, based on the nonlinear stochastic model, the interplay between the evolvability and network robustness of biological networks is discussed from the perspective of system robustness and sensitivity theory. Using the global linearization technique, the evolvability and network robustness of biological network can be measured, from the locations of the eigenvalues of linearized systems and by solving a LMI-constrained optimization method.
Genetic, phenotypic and environmental random variations are inevitable in evolution, rather than desirable features of biological networks.56,66 These random variations arise from the complexity and evolutionary history of biological systems. However, there is numerous evidence for the evolution of high fidelity and minimal noise, including the proof editing of DNA replication and protein translation. Enzymes have evolved toward high specificity. Gene expression is regulated by elaborate mechanisms, and stochastic variations seem to have been minimized, in the evolutionary process. For example, gene networks within bacterial operons may have evolved to minimize genetic and phenotypic noise and infidelity. However, chemophysical constraints regulate the specificity of biological networks and fidelity in biological networks is costly, so there is a tradeoff between the benefits of fidelity and its costs. Therefore, if biological networks want to retain enough network robustness and reduce their evolvability (small r0) to genetic, phenotypic and environmental variations, in order to keep their functional accuracy (ie, phenotype robustness), much effort is needed and a high cost must be paid. In general, random genetic and phenotypic variation and heterogeneity are neither a desired nor a deliberate outcome of evolution. 66 Heterogeneity and diversity form the very basis of evolution, not only within genetically diverse populations but also within the same allele or genome.55,65 Thus, random genetic, phenotypic and environmental fluctuations are inherent features of biological components and networks. Random perturbative biological networks may contain more widely connected and more interconnected components and may provide multifunctionality of the biological network. This multifunctionality results in an increased robustness and a capacity to cope with diverse challenges, including those that have never been encountered in evolution.41,67 However, multi-functionality also increases the complexity and the variations in the biological network, which might increase adaptive potential. Thus, behind the façade of perfection and optimality, lies the messy biology that originates from evolution. The balance of the evolvability and network robustness, in stochastic biological networks, sheds light on the mechanisms that govern the exploitation and toleration of the messiness of the euolutionary process, from the systematic perspective. In the evolutionary scenario, network robustness is an intrinsic property of the evolvability and might in the long term, improve the evolution of all levels of biological networks.5,35
In this study, a tradeoff between genetic robustness and environmental robustness in evolution is derived from the interplay between the evolvability and network robustness. Based on this tradeoff between the genetic robustness and environmental robustness of a biological network in evolution, 35 the evolutionary antagonistic pleiotropy is extended from gene level to biological level and can be discussed from the systematic perspective. In terms of gene networks, based on the evolutionary theory of aging35,38 antagonistic pleiotropy holds that what is good for an organism when it is young can be bad for it when it is old. 38 Conventionally, this discussed from the point of view of individual genes. Based on the tradeoff between the genetic robustness and environmental robustness, the antagonistic pleiotropic actions of a gene network, in the evolutionary process, can be investigated from a nonlinear stochastic system perspective. Obviously, the tradeoff between the genetic robustness and environmental robustness in the evolutionary gene network is also inherent in an evolutionary antagonistic pleiotropy.
For different levels of biological networks, two favored strategies improve the phenotype robustness in evolution. One is to improve the network robustness, to resist the genetic variations and to filter the effect of environmental stimuli. Negative feedback is the mechanism that improves network robustness and is favored by natural selection at different levels of biological networks, in the evolutionary process. Another strategy is to reduce the effect of genetic variations and external stimuli on different biological networks. Redundancies and repairs are the mechanisms of this strategy and are favored by natural selection in evolution. This is the reason why there are so many different redundancies from duplicated genes in gene networks, redundant pathways in biochemical networks and species redundancy in ecological systems. 35
Recently, synthetic biology has spawned the engineering of artificial biological systems that fulfill a particular purpose, from off-the-shelf chemical ingredients, to allow a better understanding of biological design, using synthetic circuits.41,67–73 High-throughput measurements are necessary, to determine what is evolved and how engineering genetic networks, in vivo, mimic the evolutionary process of genetic networks.74–78 An approach involving directed evolution has been used to rationally design a synthetic network, in vivo.61,66,79,80 Unfortunately, synthetic biologists are faced with the uncertainties of modeling and biochemical parametric variations, in vivo.55,65,71–73,81–85 To overcome this problem, the interplay between the evolvability and network robustness should be considered in the design procedure, so that the synthetic gene network can evolve to robustly track some desired behavior by evolution algorithm or genetic algorithm via the maximization fitness, at full speed to mimic the evolutionary process of a gene network, and then can function properly in the host cell, under the influence of parametric fluctuations and environmental disturbances.43,46,81,82,86–89 Here we provide some insights, from a stochastic system perspective, into how evolutionary biological networks sustain functions robustly, which might be useful in the design of in vivo gene networks.
In summary, this study proposes that: (i) The idea that genetic, phenotypic and environment variances pave the road for the evolvability of biological networks is an attractive one. However, their dynamic mechanisms are unclear. The proposed nonlinear stochastic systems can represent messy biological networks with intrinsic genetic and phenotypic mutations and environmental disturbances in evolution, (ii) Measures of the evolvability and the network robustness of a biological network in evolution can be obtained from the stochastical robustness theory and the H∞ filtering method, (iii) The interplay between the evolvability and network robustness of a biological network can also be discussed from a nonlinear stochastic system perspective. 52 (iv) The criterion of phenotype robustness of a biological network in evolution is found as follows: The genetic robustness plus environmental robustness have to be covered by the network robustness in (26). This is also discussed for a stochastic biological network with genetic variations and environmental stimuli, in the evolutionary process. Based on the phenotype robustness of a gene network, the evolutionary cause of genetic robustness can also be discussed with reference to the congruent mechanism in.15,35 (v) Based on the tradeoff between the genetic robustness and environmental robustness of evolutionary biological networks, the evolutionary antagonistic pleiotropy can be investigated, from the individual gene level7,10 to biological network level, from the systems biology perspective. In order to avoid solving the nonlinear HJI for the evolvability and network robustness of biological networks in evolution, the global linearization technique is proposed to simplify the analysis, so that the measures of the evolvability and network robustness in a biological network only need to solve a set of LMIs and a LMI-constrained optimization, which can be easily achieved by the help of LMI toolbox in Matlab. Furthermore, the interplay between the evolvability and network robustness can be easily discussed with reference to the stability robustness and filtering ability of a biological network, from the linear system theory point of view.
The tradeoff between the genetic robustness and environmental robustness of a stochastic evolutionary biological network is most relevant to the well-known statistical tradeoff between bias and variance.51,90 This bias/variance dilemma has been solved by some regularization method via selecting adequate regulation parameters. 51 These regularization methods may provide other insights into the tradeoff between the genetic robustness and environmental robustness of a stochastic evolutionary biological network, which will be our future works.
Disclosures
Author(s) have provided signed confirmations to the publisher of their compliance with all applicable legal and ethical obligations in respect to declaration of conflicts of interest, funding, authorship and contributorship, and compliance with ethical requirements in respect to treatment of human and animal test subjects. If this article contains identifiable human subject(s) author(s) were required to supply signed patient consent prior to publication. Author(s) have confirmed that the published article is unique and not under consideration nor published by any other publication and that they have consent to reproduce any copyrighted material. The peer reviewers declared no conflicts of interest.
Footnotes
Acknowledgements
The work was supported by the National Science Council of Taiwan under grant NSC 99-2745-E-007-001-ASP.
Supplementary Data
Appendix A: Proof of proposition 1
Appendix B: Proof of proposition 2 equation section (next)
Consider the following equality.
where
By the fact
By the fact
Then, we get
By the inequality (15), we get
This is the inequality in (14). When
Q.E.D.
Appendix C: Proof of proposition 3
For the stochastic gene regulatory network in (20), consider the following equality,
By the fact E dw
l
(t)/dt = En
l
(t) = 0, ∂V(
By the inequality in (21), we get
which is the HJI in (14). If
Appendix D: Proof of proposition 4
For the nonlinear stochastic gene network in (29), let us choose the Lyapunov function V(
(by the fact E dw l (t)/dt = En l (t) = 0).
By the inequality in (30), we get
ie, the nonlinear stochastic system in (28) is stochastically stable.
Q.E.D.
Appendix E: Proof of proposition 5
For nonlinear gene network in (31), with Lyapunov function
By Lemma 1
we get
By the inequality in (32), we get
This is the inequality in (14). If
Appendix F: Proof of proposition 6
For the nonlinear stochastic gene network in (34) with Lyapunov function
Then, by the fact
By Lemma 1, we get
Then
(by HJI in(35))
This is the inequality in (14). If
Appendix G: Proof of proposition 7
Since we replace the nonlinear stochastic gene network in (28) by the interpolation of globally linearized system in (41), then the HJI of the stochastic stability condition (30) of nonlinear stochastic gene network can be replaced by
If we choose
or
ie, if the LMIs in (42) hold, then the stochastic stability is guaranteed for the nonlinear stochastic gene network in (41).
Q.E.D.
Appendix H: Proof of proposition 8
Since we use the globally linearized system in (43) to replace the nonlinear gene network in (31), the HJI in (32) for solving the evolution level ρ could be replaced by the following inequality
If we choose
Because
The above inequality holds if the inequalities in (44) hold.
Q.E.D.
Appendix I: Proof of proposition 9
Since we use the globally linearized gene network in (40) to replace the nonlinear gene network in (34), the HJI in (35) for evolutionary evolution level ρ and network robustness in Proposition 6 could be replaced by
Therefore, if the quadratic inequalities in (49) hold, then the evolution level ρ and network robustness hold for nonlinear stochastic gene network in (34) or (40).
Q.E.D.