
Abstract
In ecological networks, network robustness should be large enough to confer intrinsic robustness for tolerating intrinsic parameter fluctuations, as well as environmental robustness for resisting environmental disturbances, so that the phenotype stability of ecological networks can be maintained, thus guaranteeing phenotype robustness. However, it is difficult to analyze the network robustness of ecological systems because they are complex nonlinear partial differential stochastic systems. This paper develops a unifying mathematical framework for investigating the principles of both robust stabilization and environmental disturbance sensitivity in ecological networks. We found that the phenotype robustness criterion for ecological networks is that if intrinsic robustness + environmental robustness ≤ network robustness, then the phenotype robustness can be maintained in spite of intrinsic parameter fluctuations and environmental disturbances. These results in robust ecological networks are similar to that in robust gene regulatory networks and evolutionary networks even they have different spatial-time scales.
Keywords
Introduction
In the last two decades, robust stabilization and noise filtering theories have been developed by control engineers in order to improve the reliability and performance of control systems. In previous papers,1–3 the network robustness and network sensitivity of gene regulatory networks and evolutionary networks have been discussed from a nonlinear, stochastic systems point of view. In these nonlinear biological networks, many local stable equilibrium points (phenotypes) exist. It has been found that if a network can confer enough intrinsic robustness to tolerate intrinsic parameter fluctuations, to buffer genetic mutations and resist environmental disturbances in gene regulatory networks or evolutionary networks, then the existing phenotype of these networks can be maintained despite intrinsic parameter fluctuations, genetic mutations and environmental disturbances. In most ecosystems, phenotype changes of ecological networks caused by losses of biodiversity may come from habitat destruction, alien species introduction, climate change, and pollution.4,5 Phenotype changes are likely to reduce not only the number of species but also the complexity of the ecological network and the function of the ecosystem. To maintain a preexisting phenotype, the ecological networks, like food webs, should be more robust to bear random or selective removal of species in the trophic links. In this study, results will be extended from gene regulatory networks and evolutionary networks at the molecular level to ecological networks on a large scale, while considering the compartment effect of habitats.
Darwin used the metaphor of a ‘tangled bank’ to describe the complex interactions between species. All interactions between species can be visualized as ecological networks in which species are linked together, either directly or indirectly, through intermediate species. 6 Ecological networks, although complex, have well-defined patterns that both illuminate the ecological mechanisms underlying them and promise a better understanding of the relationship between complex and ecological stability.7,8 Ecological research has widely demonstrated that community fragility is far from being understood. It is commonly accepted that community fragility and persistence are related to the way in which ecological communities are structured, specifically to the distribution of trophic links throughout the community.8,9 Nevertheless, both the scarcity of high-quality data and lack of methods suitable for a detailed analysis of the complexity of ecological networks result in the lack of a unified picture of community fragility and persistence. 10 In recent years, some topological features, the so-called “small world” behavior and “scale-free” distributions of links have been applied to ecological networks for examining ecosystem stability against different types of species loss. 8 Although ecological networks are generally not small-world, scale-free networks, this topology is consistent with patterns found within those classes of networks. 11 In general, increasing biodiversity increases the likelihood that an ecosystem will have, first, species that will respond differently under variable environmental conditions and perturbations, and second, functional redundancy; that is, species that are capable of functionally replacing extinct species. The higher levels of biodiversity present in a non-fragmented food web might support several ecosystem functions that a fragmented food web would not manage. 12 High biodiversity also reduces the probability of secondary extinctions. 13 An analysis considering interaction strengths among species would be very useful in confirming whether highly connected species play a stabilizing role in ecosystems, but early studies provide little or no information on the strength of trophic links because identifying strength of trophic links is difficult. Recently, some analyses based on predator-to-prey interactions propose that body size ratio can predict trophic strength. The larger the ratio, the stronger the interaction,14,15 which can link the food web to the relationship between body size and numerical abundance for ecological network analysis. 16 As we know, ecological networks can be very robust against random removals of species, but can be extremely fragile when selective attacks are used.5,17 These further models involving species removal according to criteria other than the most-connected species would help establish a more global view of the effects of human-driven disturbances. 12 In this study, nonlinear stochastic partial differential systems will be employed to describe ecological networks under intrinsic perturbations and environmental disturbances. The robust stabilization problem of large-scale ecological networks will be discussed with consideration of the compartment effect of habitat. Robust stabilization (persistence) and environmental sensitivity (community fragility) will be investigated from a systematic perspective. The finite difference scheme will be employed to treat the species diffusion operation for easy estimation of network sensitivity and network robustness of ecological networks in a habitat.
To measure the network sensitivity of an ecological network relies on solving a Hamilton Jacobin integral inequality (HJII)-constrained optimization problem. Based on the HJII, we could find that if the network robustness can confer intrinsic robustness to tolerate intrinsic parameter fluctuations and environmental robustness to resist the environmental disturbances simultaneously, then the phenotype of ecological network could be maintained in spite of intrinsic parameter fluctuations and environmental disturbances. The framework of phenotype robustness criterion in an ecological network is similar to that of the gene regulatory network and evolutionary network at the molecular level. Moreover, the tradeoff between network robustness, intrinsic robustness and environmental robustness are also found to be the same at different biological levels from gene regulatory networks and evolutionary networks to ecological networks.
However, it is still very difficult to solve HJII for phenotype robustness criterion in order to discuss the tradeoff between network robustness, intrinsic robustness and environmental robustness in ecological networks. In this study, the global linearization technique, 1 finite difference scheme and spatial state space method18–20 are employed to transform the HJII-constrained optimization problem for network sensitivity to an equivalent linear matrix inequalities (LMIs)-constrained optimization problem, which can be easily solved with the help of LMI toolbox in Matlab. 21 Then the phenotype robustness criterion of the ecological system can be discussed from the local linearized system point of view. If the eigenvalues of local linearized ecological systems are in the farther left hand complex domain (more stable), then the ecological network will have more network robustness. Finally, to verify the existence of phenotype robustness criterion and to estimate the network sensitivity of a nonlinear stochastic partial differential ecosystem, an in silico example is given to illustrate the tradeoff between the network robustness, intrinsic robustness and environmental robustness in an ecological network.
Tradeoff between Intrinsic Robustness, Environmental Robustness and Network Robustness in Ecological Biology
Examples of robust biological systems are found on many scales, from biological genetic networks to ecological networks. In previous papers,1–3,22 we have discussed the robust stabilization problem of gene regulatory networks and evolutionary networks at the molecular level. In this section, the robust stabilization problem of a large-scale ecological network will be discussed with consideration of the compartment effect of habitat.
Consider the following ecological network in the spatial domain:8–10,23
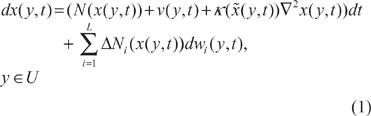

Then we want to discuss the robust stabilization problem of the nonlinear stochastic partial differential ecological system in (1). Suppose the origin is shifted to the equilibrium point (phenotype) of interest, then (1) is modified as follows:18,24

In general, it is still very difficult to estimate the network sensitivity of nonlinear stochastic partial differential ecological networks in (2) to environmental disturbances directly. We will estimate its upper bound, ie, its network sensitivity level, and then decrease this upper bound to approach the network sensitivity indirectly. In this situation, the environmental disturbance sensitivity level for the nonlinear stochastic partial differential ecological network in (1) is to consider the total multi-species interaction domain 

According to the spatial-temporal environmental disturbance sensitivity in (3), we get the following phenotype robustness (community stability) criterion for the ecological system in (1).

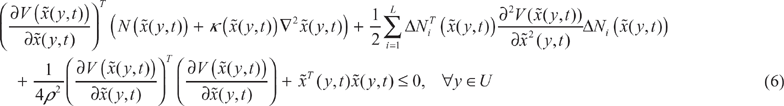
The network sensitivity 
According to the above network sensitivity
To maintain proper functions of an ecosystem that can produce enough foods or services for species, enough network robustness should be ensured for the ecological network in face of intrinsic parameter fluctuations, such as changes of species from trophic links, and environmental disturbances, such as the invasion of new species or pollution from human activities. In order to increase the network robustness on the

If the ecological network in (1) does not have enough network robustness to guarantee the phenotype robustness criterion in (8), the community stability of ecological system cannot be maintained. In this situation, in order to maintain the phenotype at 

In general, it is not easy to solve HJII in (5) or (8) for the phenotype robustness criterion of the nonlinear stochastic ecological network in (1). The nonlinear stochastic ecological network in (1) can be represented by the interpolation of local linearized systems according to the global linearization method as follows:
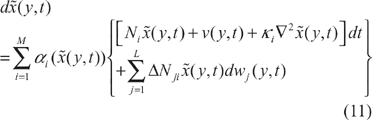
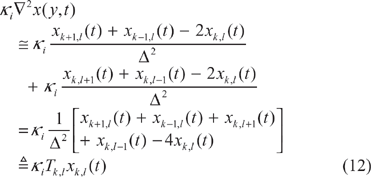

Finite difference grids of a two-dimensional space
Following this, according to the finite difference approximation in (12), the nonlinear stochastic partial differential ecological network in (11) can be represented by the following stochastic finite difference ecological system:
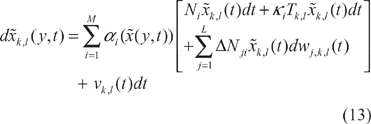
To simplify the representation of stochastic finite difference ecological system in (13), we definite a spatial state vector 


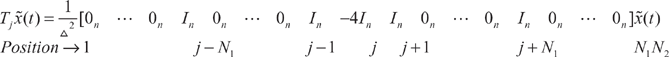
Let us denote
Using the properties of Kronecker product, we have the following properties:
By using the Kronecker product in Lemma 1, the stochastic finite difference ecological systems in (14) at all grid points in habitat U can be integrated as the following stochastic spatial space system:

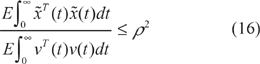
Then, based on the stochastic spatial space dynamic equation of ecological system in (15), we get the following phenotype robustness (community stability) criterion for ecological systems.

Similarly, the network sensitivity 
Therefore, after replacing ρ in (17) with the above network sensitivity p0, the phenotype robustness (community stability) criterion of ecological network in (17) could be modified as the following form:

It is seen that if each local linearized ecological partial differential system has enough local network robustness to confer local intrinsic robustness and local environmental robustness, then the phenotype of nonlinear stochastic partial differential ecological systems is robust under intrinsic parameter fluctuations and environmental disturbances. The tradeoff between local intrinsic robustness and local environmental robustness can be seen from (19). If the eigenvalues of each local system matrix [

The smaller distance between the locations of eigenvalues of [
State feedback control to improve the network robustness of ecological networks in (9) can also be approximated by the global linearization and finite difference scheme as follows:


Then, the robust ecological network design must specify an adequate
Computer Simulation
In the real world, most phenomena are governed by partial differential equations such as population dynamics in ecology, chemical dispersion in chemistry and heat flow or wave propagation in physics.31,32 In this section, a simple ecological example for nonlinear stochastic partial differential system is given to illustrate and to confirm the phenotype robustness criterion of the ecological network. This mono-species population system is used to describe the dynamical interaction between the native species and the environmental disturbances in a two-dimensional spatial domain.
23
The individual's spatial dispersal in the field of ecological biology can be seen as the heat transfer or fluid diffusion in physics. Suppose the mono-species population system with stochastic intrinsic fluctuation and environmental disturbance could be formulated as follows:
20
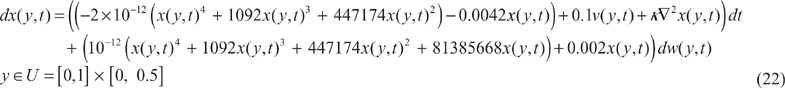
The spatial-time profiles of the nonlinear partial differential system is approximated by global linearization scheme with vertices 
Discussion
There are many examples of robust biological systems found at many scales, from biochemical networks to ecological networks. At each scale, robustness may reflect the properties of individual elements or, alternatively, the dynamic feedback between interacting elements. For example, the expression of some metabolic functions may be robust in the face of temperature changes, because an enzyme maintains its shape and specificity across a range of temperatures or because an interconnected network of reactions sustains the supply of product, even when some enzymes fail. A genome may be robust because it encodes proofreading and repair systems that reduce replication errors or because it gets organized such that many mutations have little effect on its phenotype. An ecosystem might be robust if it resists the extinction of some keystone species or, if extinction does occur, because surviving species can compensate over physiological, demographic, or evolutionary time scales.33,34
Ecological networks may pose a challenge when one considers severe changes such as the addition or removal of species. 17 By topological approaches (without population dynamics), it is seen that the network will appear ‘species deletion unstable’. 35 ie, the network quickly breaks into many disconnected subnetworks when the most connected species are successively removed from the network. Dynamical models of species removals also show that the more polyphagous the predators, the less the effect caused by removing one of its prey species. 35 Thus, whether simple or highly connected model food webs are robust to the loss of species depends entirely on whether one looks at top predators or plant species. 36 By contrast, when disasters ruin species randomly, these networks will be more robust, showing both little fragmentation and few secondary extinctions.5,7–12 But a lack of integrated concepts about how an ecological network can maintain its phenotype robustness under random or selective changes and how to evaluate the network sensitivity still exists. This study has developed a unifying mathematical framework for investigating the principles of both robust stabilization and environmental disturbance sensitivity in ecological networks from a systematic perspective.
From the gene regulatory network to the ecological network, it can be seen that, for different scales of biological systems, their phenotype robustness criteria according to their global linearization models can be demonstrated within a unifying mathematical framework, even when the partial differential diffusion operator is considered in the ecological network. From the systems theory perspective, the phenotype robustness of nonlinear stochastic gene networks in systems, evolutionary 1 and ecological biology needs to obey a similar phenotype robustness criterion, ie, “intrinsic robustness + genetic robustness + environmental robustness ≤ network robustness”. This means that network robustness needs to be strong enough to tolerate either heritable perturbations (genetic variations) or non-heritable perturbations (ie, random molecular fluctuations and environmental disturbances) so that the phenotype of biological networks can be maintained in systems, evolutionary and ecological biology using a similar mathematical framework.
In general, random genetic variations, phenotype perturbations and heterogeneity are neither desired nor deliberate outcomes of systems, evolutionary and ecological biology. However, heterogeneity and diversity form the very basis of evolutionary biology, not only within genetically diverse populations but also within the same allele or genome. Thus, random genetic variations, environmental disturbances and phenotypic perturbations are inherent features of biological systems and networks. Random perturbative biological networks may contain more connected and interconnected systems, which may provide multifunctionality of the biological network. This multifunctionality may result in increased robustness and a capacity to cope with diverse challenges. However, multifunctionality also increases the complexity and variations of the biological network, which may increase adaptive potential. Thus, behind the façade of perfection and optimality of systems, evolutionary and ecological biology lies the messy biology that originates from the genetic variations and environmental disturbances in evolution. There exists the tradeoff among intrinsic robustness, genetic robustness, environmental robustness and network robustness in the phenotype robustness of stochastic biological networks. That is, if intrinsic robustness + genetic robustness + environmental robustness ≤ network robustness, then the phenotype of the biological network is maintained. This sheds light on the mechanisms that govern the exploitation and toleration of the messiness of biological networks in systems, evolutionary and ecological biology, from a systematic perspective. Obviously, network robustness needs to be strong enough to tolerate either heritable perturbations (genetic variations) or non-heritable perturbations (random molecular fluctuations and environmental disturbances) so that the phenotype can be maintained in biological networks at different levels.
For gene regulatory networks, evolutionary networks 1 and ecological networks, two favored strategies can improve phenotype robustness in the network evolutionary process. One is to improve network robustness to provide enough intrinsic robustness for tolerating intrinsic parameter fluctuations, genetic robustness for buffering genetic variations and environmental robustness for resisting environmental disturbances, so that phenotype robustness of these biological networks at different scales can be maintained under these uncertain perturbations and environmental disturbances. Negative feedback is a mechanism that can improve network robustness (ie, it can make right-hand sides of (8), (19) larger) and is favored by natural selection in biological networks at different levels in the network evolutionary process. Another strategy is to reduce the effect of intrinsic parameter fluctuations, genetic variations and environmental disturbances on different biological networks (ie, it can make the left-side of (8), (19) smaller). Redundancies and repairs are the mechanisms of this strategy and are favored by natural selection in evolution. This is the reason why there are so many different redundancies from duplicated genes in gene regulatory networks, redundant pathways in biochemical networks and species redundancies in ecological systems.
From the phenotype criteria of the ecological network in (8) and (19), if the network robustness in the right-hand side is fixed, it is seen that if the intrinsic robustness becomes large to tolerate more intrinsic parameter fluctuations in the first term, it will lead to the decrease of environmental robustness or the increase of the network sensitivity
Conclusion
This paper presents a unifying mathematical framework to describe stochastic ecological networks under intrinsic parameter fluctuations and environmental disturbances. Then, according to the unifying stochastic biological systems, the phenotype robustness criteria of ecological networks are also investigated from the robust stabilization and network sensitivity perspective. It is found that if the phenotype criterion “intrinsic robustness + environmental robustness ≤ network robustness” is guaranteed, ie, network robustness can confer intrinsic robustness for tolerating intrinsic parameter fluctuations and environmental robustness for resisting the environmental disturbances, then the phenotype will be robust in ecological networks. Using the global linearization method and finite difference scheme to estimate network robustness, based on spatial state space system, we found that if the network robustness of each local linearized ecological network is greater than the total sum of intrinsic robustness and environmental robustness of each local linear ecological network, then the phenotype of the ecological network is also maintained, despite intrinsic parameter fluctuations and environmental disturbances. We also found that the phenotype robustness criterion and the tradeoff among network robustness, intrinsic robustness and environmental robustness are similar for different scales of biological networks, from gene regulatory networks and evolutionary networks to ecological networks. Finally, if the network structure is given, by taking into account different strengths of trophic links, or the removal of species in trophic links, the phenotype robustness criterion may also be employed in food webs to discuss the network robustness with random or selective removal of species in the trophic links.
Funding sources
The work was supported by the National Science Council of Taiwan under grant NSC 100-2745 -E-007-001-ASP and NSC 101-2745-E-007-001-ASP.
Competing Interests
Author(s) disclose no potential conflicts of interest.
Footnotes
Author Contributions
Conceived and designed the experiments: BSC. Analysed the data: BSC. Wrote the first draft of the manuscript: BSC. Contributed to the writing of the manuscript: BSC, YPL. Agree with manuscript results and conclusions: BSC. Jointly developed the structure and arguments for the paper: BSC, YPL. Made critical revisions and approved final version: BSC, YPL. All authors reviewed and approved of the final manuscript.
Disclosures and Ethics
As a requirement of publication author(s) have provided to the publisher signed confirmation of compliance with legal and ethical obligations including but not limited to the following: authorship and contributorship, conflicts of interest, privacy and confidentiality and (where applicable) protection of human and animal research subjects. The authors have read and confirmed their agreement with the ICMJE authorship and conflict of interest criteria. The authors have also confirmed that this article is unique and not under consideration or published in any other publication, and that they have permission from rights holders to reproduce any copyrighted material. Any disclosures are made in this section. The external blind peer reviewers report no conflicts of interest. This manuscript has been read and approved by all authors. The authors and peer reviewers report no conflicts of interest.