
Abstract
Proteases play important roles during tumor angiogenesis, invasion, and metastasis. Various molecular imaging techniques have been employed for protease imaging: optical (both fluorescence and bioluminescence), magnetic resonance imaging (MRI), single-photon emission computed tomography (SPECT), and positron emission tomography (PET). In this review, we will summarize the current status of imaging proteases in cancer with these techniques. Optical imaging of proteases, in particular with fluorescence, is the most intensively validated and many of the imaging probes are already commercially available. It is generally agreed that the use of activatable probes is the most accurate and appropriate means for measuring protease activity. Molecular imaging of proteases with other techniques (i.e. MRI, SPECT, and PET) has not been well-documented in the literature which certainly deserves much future effort. Optical imaging and molecular MRI of protease activity has very limited potential for clinical investigation. PET/SPECT imaging is suitable for clinical investigation; however the optimal probes for PET/SPECT imaging of proteases in cancer have yet to be developed. Successful development of protease imaging probes with optimal in vivo stability, tumor targeting efficacy, and desirable pharmacokinetics for clinical translation will eventually improve cancer patient management. Not limited to cancer, these protease-targeted imaging probes will also have broad applications in other diseases such as arthritis, atherosclerosis, and myocardial infarction.
Introduction
Over the last decade, considerable advances have been made to further our understanding of cancer invasion and metastasis. Although surgery and radiation therapy can effectively control many cancers at their primary sites, metastatic diseases still have very poor prognosis. 1 Metastasis, the spread of cancer from its primary location to a distant organ, is the main cause of death in cancer patients. Cancer metastasis occurs as a series of sequential and interrelated processes which include dissociation of cancer cells in the primary tumor, local invasion, angiogenesis, intravasation of cancer cells into the vasculature or lymphatic systems, survival in these channels, extravasation, and proliferation at a distant site.1,2 Successful inhibition of tumor metastases is a promising approach for cancer therapy and/or cancer control.
Proteases belong to a group of evolutionarily conserved enzymes which are activated in response to the stimuli in normal cells. They can regulate a variety of different cellular processes such as gene expression, differentiation, and cell death. 3 Briefly, proteases can be classified into six groups by their mechanisms of action: serine, cysteine, threonine, aspartate, glutamic acid proteases, as well as metalloproteases. The threonine and glutamic acid proteases were not discovered until 1995 and 2004, respectively, and glutamic acid protease is the only subtype not found in mammals so far. 4 The mechanism for cleaving a peptide bond with a protease typically involves a serine, cysteine, or threonine residue (typically a histidine is close-by which can activate these residues) or a water molecule (in aspartate, metallo- and glutamic acid proteases) as the nucleophile in the active site. 5
Tumor progression and metastases are highly dependent on oxygen and nutrient supply, which are stimulated by multiple proteases in the tumor and/or surrounding tissues. Many tumors have been shown to have elevated levels of proteases at an early stage and these proteases are thought to be crucial for tumor angiogenesis, invasion, and metastasis. 6 Further understanding of the roles of proteases in tumor progression will guide the development of novel therapeutic strategies against cancer.
Although the clinical significance of detecting protease activity in vivo has been well recognized, it is quite difficult to achieve. Early detection of small primary tumors is crucial for successful cancer therapy and patient management. Molecular imaging, defined as the in vivo characterization and measurement of biologic processes at the cellular and molecular level,7,8 can fulfill this goal. Different from traditional diagnostic imaging techniques such as computed tomography (CT), ultrasound, and magnetic resonance imaging (MRI), molecular imaging aims to detect the molecular abnormalities that are the basis of the disease.
Molecular imaging of cancer can be a highly useful tool in clinical medical practice. 9 Systematic efforts are under way to detect both primary tumors and metastatic diseases at their early stages. The insights gained into the biological or molecular events in living subjects will accelerate the development of novel therapeutics agents and provide more accurate means to monitor the effects of these agents. 10 Various molecular imaging modalities have been employed for protease imaging: optical (both fluorescence and bioluminescence), MRI, single-photon emission computed tomography (SPECT), and positron emission tomography (PET). The probes used in these studies can be briefly categorized into two major types: substrate-based and activity-based.11–13 In this review, we will summarize the current state-of-the-art of protease imaging in cancer.
Fluorescence Imaging of Protease Activity
Optical imaging is less expensive and more convenient than the other imaging modalities such as MRI and PET. 14 The most widely used optical imaging techniques are fluorescence imaging and bioluminescence imaging (BLI). Many other optical imaging techniques are also under active development, such as Raman spectroscopy and photoacoustic imaging.15,16 Fluorescence imaging is by far the most widely used technique for imaging protease activity in vivo and the proteases that have been imaged include the cathepsin family, the matrix metalloprotease (MMP) family, and the urokinase plasminogen activator (uPA).
Most of the probes used in these studies are called “smart probes” or “activatable probes”, which can change their optical properties after protease cleavage. Typically, fluorescently labeled substrates are designed to be maximally quenched by a quencher (in some cases the fluorescent dye itself) in close proximity because of fluorescence resonance energy transfer (FRET).17,18 Upon cleavage, the fluorophore and the quencher are separated which resulted in an enhanced fluorescence signal.
Imaging of cathepsins
During tumor metastasis, proteases play an important role in mediating the passage of tumor cells between tissue and vessels via degradation of the basement membrane. Cathepsin is a lysosomal cysteine protease family involved in cellular protein degradation. 19 It is over-expressed in many tumor cells as well as the host cells associated with the tumor. 20 The ubiquity of cathepsins makes them very attractive targets for tumor detection.
There are five major types of cathepsins: cathepsin B, D, S, K, and L. Cathepsins B and L are major cysteine proteases involved in protein degradation within the lysosome. 21 Cathepsin D is a proteolytic enzyme that has been implicated in the degradation of basement membranes.22,23 Cathepsin S belongs to the papain superfamily and a difference in cathepsin S level was found between the tumor and adjacent control tissue in lung cancer patients, which suggested that cathepsin S may be involved in cancer progression. 24 Cathepsin K has strong collagenolytic activity and mediates matrix degradation, a pivotal step in tumor invasion and metastasis. 25 To date, optical imaging studies have been reported for cathepsins B, D, and S.
The first in vivo optical imaging of protease activity was demonstrated in a xenograft lung carcinoma model a decade ago (Fig. 1). 26 Near-infrared fluorescent (NIRF) fluorophores were bound to a long circulating graft copolymer consisting of poly-L-lysine and methoxy-polyethylene glycol (PEG) succinate. Following intravenous injection, the NIRF probe accumulated in solid tumors due to its long circulation life-time and leakage through the tumor neovasculature, termed the “enhanced permeability and retention (EPR)” effect.27,28 Intratumoral NIRF signal was generated by proteolytic cleavage of the macromolecule which released the fluorescence signal of the previously quenched fluorochrome.
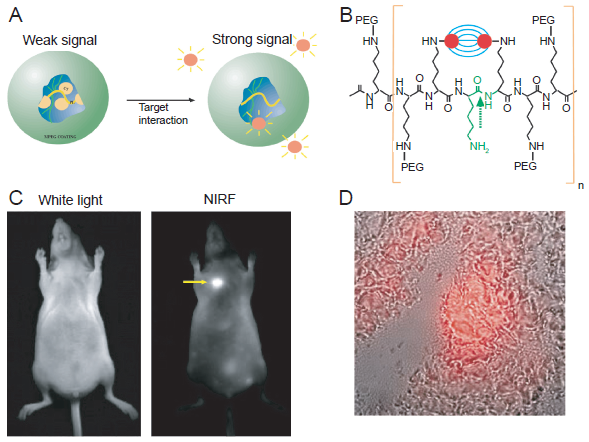
Fluorescence imaging of tumors with a protease-activated NIRF probe.
In vivo imaging revealed a 12-fold increase in the NIRF signal, allowing the detection of tumors with sub-millimeter diameter. Although this probe was not specific for a single protease (it can be activated by many cysteine/serine proteases), the generation of enhanced NIRF signal as a result of lysosomal activity did open up a new research area that has been vibrant over the last decade.
Similar probes were tested in a breast cancer xenograft model in mice, 29 demonstrating that this strategy can be used to detect early stage tumors, probe for specific enzyme activity, and evaluate the therapeutic effect in vivo. Subsequently, the same type of NIRF probe was used to determine that cathepsin B activity in the tumor correlated with the aggressiveness of breast tumor phenotypes. 30 These probes have later been adopted to reveal colonic neoplasms in mice, 31 in which the signal intensity of the tumor was more than 30-fold greater than the control animals. This study demonstrated that the use of NIRF imaging microcatheters, in combination with protease-activatable probes, could delineate tumors with good contrast, which may be a potentially useful adjunct to white light colonoscopy in the clinic.
With the development of a fluorescence-mediated tomography (FMT) system, the up-regulation of cathepsin B was imaged semi-quantitatively in a HT-1080 fibrosarcoma tumor model. 32 The key feature of the FMT system is that the fluorescence signal in deep tissues can be reconstructed to give a three-dimensional image of the probe distribution in vivo.33,34 This study demonstrated that fluorescence reflectance imaging and FMT using these protease-activatable probes can allow the detection of experimental spontaneous breast tumors. 32 Because the activity levels of various proteases may correlate with the clinical outcome, this technique may help not only to detect, but also to differentiate breast cancer phenotypes non-invasively in the future. To date, the research on FMT has been limited to small animal models due to the limited light penetration even in the NIR range. It is likely that clinical FMT systems will be developed in the future which can help breast cancer patient management, as an adjunct to the other modalities such as CT and PET.
Besides in vivo imaging applications, some probes were designed to visualize the dynamic alteration of cathepsin B with multichannel microscopy. 35 A series of quenched activity-based probes (qABPs) become fluorescent upon activity-dependent, covalent modification by proteases (Figs. 2A and B). These reagents were able to freely penetrate cells and allow direct imaging of protease activity in living cells, which have been used to monitor real-time protease activity in live human cells with fluorescence microscopy. Subsequently, these qABPs were applied to determine the role of cathepsin B in invasive growth and angiogenesis during multi-stage tumorigenesis. 36 Recently, these qABPs were further optimized with NIRF fluorophores which were able to give a whole-body readout in mice (Fig. 2C). 37
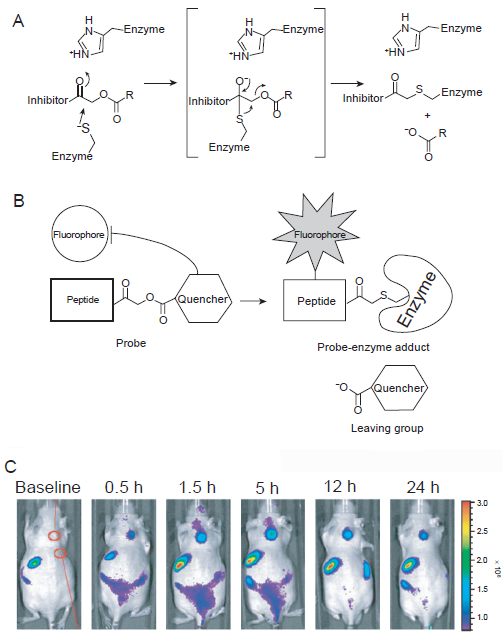
Fluorescence imaging of cathepsin activity using quenched activity-based probes (qABPs).
Other subtypes of cathepsin have also been investigated for tumor detection. A NIRF probe for cathepsin D imaging was reported a decade ago. 38 A NIRF fluorochrome was attached to the amino terminal of a peptide sequence with 11 amino acid residues (GPICFFRLGKC), which was specific for cathepsin D. The peptide was then linked to a synthetic graft copolymer. A striking 350-fold signal amplification was observed upon protease cleavage during in vitro testing with a rodent tumor cell line stably transfected with human cathepsin D. However, in vivo imaging was not achieved, likely due to the low expression level of cathepsin D in physiological conditions. A fluorescent probe for cathepsin S imaging has also been designed which incorporated its substrate, the Leu-Arg dipeptide. 39 Again, no in vivo studies have been reported.
Imaging of MMPs
Besides cathepsins, MMPs are also attractive targets for cancer imaging. Tumor-associated MMP expression and activity originated from not only tumor cells, but also the surrounding stromal cells. In fact, studies have shown that in certain epithelial cancers, most of the upregulated MMPs are expressed by the host stromal cells. 40 Multiple MMPs are induced in connective tissue cells, such as fibroblasts and inflammatory cells in response to tumor formation, which can provoke invasion of the malignant epithelial cells. 41 MMPs are also expressed in tumor cells. For example, MMP-7 is commonly expressed in the epithelial component of adenocarcinomas and several MMPs are expressed in the malignant epithelium of tumors that have undergone an epithelial-to-mesenchymal transformation. 41
MMP-2 has been recognized as one of the key MMPs involved in tumor angiogenesis, invasion, and metastasis. It is capable of degrading type IV collagen, the major component of basement membranes. 42 The first molecular probe capable of imaging MMP activity was a slight modification from the original protease sensing probes, which incorporated a peptide linker that can be cleaved by MMP-2 and certain other proteases. 43 The peptide substrate (GPLGVRGKC) was labeled with a Cy5 dye, which can self-quench when in close proximity. This probe allowed for the assessment of tumor-associated MMP activity in response to treatment with a MMP inhibitor (MMPI), Prinomastat. After demonstrating its capability to sense and image MMP response directly in vivo, this probe was also tested in other tumor models and high signal-to-background ratio was achieved in MMP-2 positive tumors. 44
In one study, a protease sensitive probe was developed which incorporated cell penetrating peptides. 45 The fluorescein- or Cy5-labeled probe can penetrate into the cytosol of mammalian cells, upon cleavage of the peptide sequence (PLGLAG) by MMP-2/9. This new strategy could selectively deliver molecules to tumor cells, allowing for accumulation and concentration of imaging or therapeutic agents within the tumor cells or within the vicinity of MMP-2/9. In another report, a self-assembling homotrimeric triple helical peptide, incorporating segments of type V collagen, with high specificity to MMP-2/9 has been reported. 46 This agent utilizes a pair of fluorophores conjugated to the ε-amino groups of lysine on both sides of the cleavage site, which can efficiently quench the signal and fluoresce after hydrolysis by MMPs.
Nanotechnology has also been applied for MMP-2/9 imaging in vivo. A gold nanoparticle- and Cy5.5-based NIRF probe for MMP imaging in vivo was designed (the substrate sequence: PLGVR). 47 This probe exhibited satisfactory imaging results in a SCC7 squamous cell carcinoma model and could also be useful in drug screening. Recently, a protein interaction-based fluorescence imaging system was designed to image MMP-2 activity and its reaction to MMPIs. 48 This study provided a direct method of molecular target assessment which may benefit MMP-targeted drug screening.
Aside from MMP-2/9, MMP-7 is another important target for tumor imaging. In one study, dendrimer-based fluorogenic substrates were developed to image MMP-7 activity. 49 The substrate used in this study, (Ahx)RPLALWRS(Ahx)C where Ahx represents aminohexanoic acid, was shown to be more selective for MMP-7 than the other MMPs within the tumor microenvironment. Recently, a similar construct using NIR FRET pairs was developed to reduce the absorption and scattering of the fluorescence signal, thereby improving the response of a proteolytic beacon. 50 This approach was shown to be highly sensitive and tumors as small as 0.11 cm in diameter could be detected.
An alternative approach to image MMP-7 activity has been developed using activatable quantum dots (QDs). Comparing to conventional fluorophores such as Cy5, QDs offer brighter signal, better photostability, and the enormous potential for multiplexed imaging.15,51 In one report, the QD fluorescence was quenched (based on FRET) by a quencher molecule (BHQ-1) with an absorption band that overlaps the QD emission. 52 The BHQ-1 was linked to the QD through a MMP-7-cleavable peptide (GPLGLARK). Adjusting the QD/BHQ-1 ratio and the peptide conformation was able to modify the sensitivity and signal-to-background ratio of the reagent, as well as controlling the MMP-7-mediated photodynamic cytotoxicity in cancer cells. In another study, MMP-2/7-mediated uptake (substrate sequence: PLGVRG) of QDs into tumor cells was reported. 53 Such protease-modulated cellular uptake of QDs may also be applied to other nanoparticles for biological imaging and selective drug delivery into tumor cells.
Imaging of uPA
A significant body of in vitro and in vivo data has established the uPA system as a promising target for cancer drug development.54,55 It has been demonstrated to participate in multiple processes during tumor development, tumor progression, and angiogenesis. In 2004, the development of a uPA activatable NIRF probe was reported. 56 This probe consists of a uPA-cleaving motif (GGSGRSANAKA), terminally capped with different NIR fluorochromes (Cy5.5 or Cy7), and a PEGylated poly-L-lysine graft copolymer. Upon addition of recombinant human uPA to the probe, significant fluorescence signal amplification (up to 680%) was observed while no activation was detected with the negative control compounds or in the presence of uPA inhibitors. Subsequently, this uPA-sensitive probe was tested in two distinct tumor models (human colon adenocarcinoma HT-29 and human fibrosarcoma HT-1080). 57 The observed changes in the fluorescence signal, about three-fold higher in animals with probe injection, correlated well with tumor-associated uPA activity.
Generally speaking, there are several advantages of using proteases as the targets for optical imaging probe development. These enzymes are secreted and activated in the extracellular environment, which partially avoid the need of delivering the probe to intracellular compartments. Proteases are generally active at the physiological pH, and perhaps most importantly, such activity is catalytic which provides an opportunity for signal amplification that can not be achieved for probes based on direct binding to their targets. However, despite all these advantages, optical imaging has very limited potential for clinical application, even with NIRF probes, because of the limited tissue penetration of the fluorescence signal (typical less than 1–2 cm).58,59 In the clinical setting, optical imaging is only suitable for imaging lesions close to the skin (e.g. skin and breast cancer), tissues accessible by endoscopy (e.g. colon cancer), or during surgery (e.g. visualizing the tumor margins). Overall, fluorescence imaging is more suitable for small animal studies which can serve as a less expensive, convenient platform/model for future clinical research.
BLI of Protease Activity
BLI is another optical imaging technique that has virtually non-existent background signal, a major limitation for fluorescence imaging even in the NIR range. 60 Most BLI studies use D-luciferin as the firefly luciferase substrate, which is tolerant to very limited chemical modification.61,62 Therefore, the biggest limitation of using BLI to detect MMP activity is the requirement for S’ residues in the recognition site, since the downstream residues (luciferin in this case) can dramatically interfere with MMP activity. The Promega Corporation recently presented a novel strategy to overcome this limitation. 60 Using a circularly-permuted firefly luciferase, they joined the original termini of the firefly luciferase with a protease cleavable peptide sequence, thereby locking the luciferase in an inactive state. Cleave of the protease substrate results in a conformational change that activates the luciferase, which neatly provides the opportunity for in vivo imaging of MMP (or other proteases if other peptide sequences are used) activity with BLI. In vivo imaging with this technology has not been reported yet.
Caspases are crucial mediators of programmed cell death (apoptosis). Among them, caspase-3 is a frequently activated death protease, catalyzing the specific cleavage of many key cellular proteins.63,64 BLI of caspase-3 has been reported for the evaluation of tumor apoptosis or the responses to drugs. 65 In this pioneering report, tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-mediated apoptosis and resulting changes in tumor burden was imaged in the same animal by dual-substrate BLI, i.e. with D-luciferin (the natural substrate for firefly luciferase) and DEVD-luciferin (active only after caspase cleavage) (Fig. 3). Surprisingly, no follow-up study has been reported since. Another group has also reported the use of the same probe in a cell-based study. 66
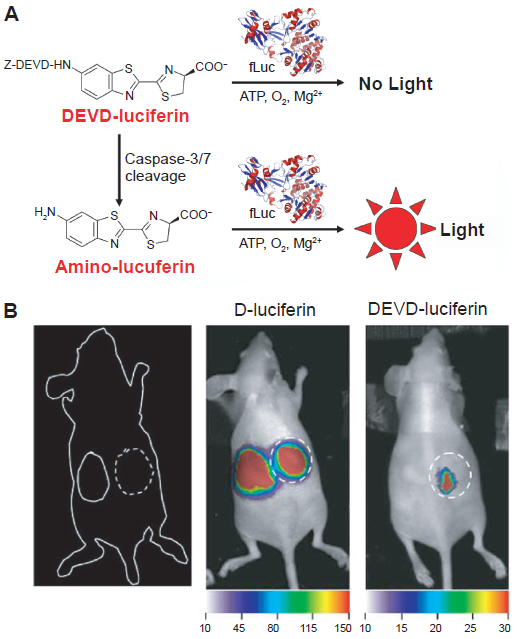
Bioluminescence imaging of protease activity in vivo.
BLI is superior to fluorescence imaging for many applications since it does not require any external light excitation and there is minimal background signal. A few proof-of-principle studies have been reported for BLI of proteases as discussed above. Although it can serve as an invaluable platform for in vitro and cell-based assays, there is still a long way to go before these agents can be widely used in animal studies. Much further effort will be needed in the future yet the high demand for protein engineering expertise in BLI of MMPs makes such techniques not readily generalizable. Lastly, one has to bear in mind that BLI is not applicable for clinical studies therefore it can only be used in cell and animal studies. To date, a variety of proteases have been studied with optical imaging techniques. However, studies on MRI, SPECT, and PET imaging of proteases have so far exclusively focused on the most intensively studied proteases, the MMPs.60,67
MRI of MMPs
MRI has become widely available in hospitals and clinical centers throughout the world and it is a commonly utilized modality for cancer imaging. 68 It detects the interaction of protons (or certain other nuclei) with each other and with the surrounding molecules in a tissue of interest. 69 Different tissues have different relaxation times that can result in endogenous MR contrast. Exogenous contrast agents (CAs) can further enhance this by selectively shortening either the T1 (longitudinal) or T2 (transverse) relaxation time.70,71 The MR image can be weighted to detect the differences in either T1 or T2 by adjusting certain parameters during data acquisition. Traditionally, Gd3+-chelates have been used to enhance the T1 contrast 72 and iron oxide nanoparticles have been used to increase the T2 contrast. 73 The advent of activatable fluorescent probes also accelerated the development of probes for MRI of enzyme activity. 74
Protease-modulated CAs were developed to detect the activity of MMPs in vivo with MRI. 75 The CAs were based on the concept of a solubility switch, from hydrophilic to hydrophobic, which significantly modifies the pharmacokinetic properties of the agents as evidenced by the slow efflux kinetics from the activity site. In live tumor-bearing mice, these CAs were demonstrated to be effective in non-invasive imaging of MMP-7 activity. Recently, acleavagemotifofMMP-9(GGPRQITAG) was incorporated into a nanosensor for in vitro detection of protease activity by MRI (Fig. 4). 76 The experimental results suggested that upon protease cleavage, the nanoparticles rapidly switch from a stable low-relaxivity stealth state to become adhesive, aggregating high-relaxivity particles. However, this agent has not been investigated in animals yet.
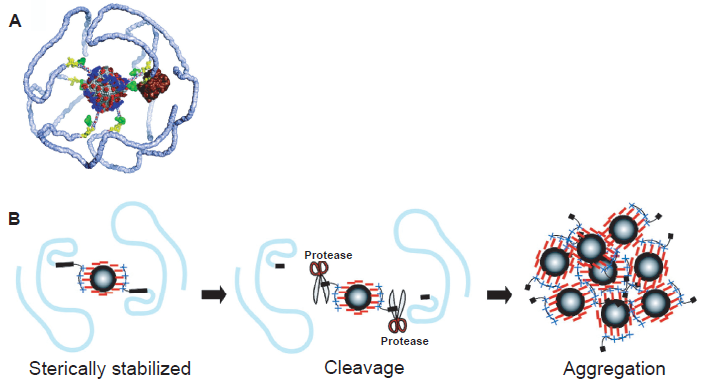
Protease-sensitive agents for MRI.
Protease-modulated CA designed to detect MMP-2 activity was also reported and the accumulation of protease-cleaved CAs in a MMP-2-positive tumor was achieved in a mouse model. 77 This agent used a peptide substrate (SGESPAYYTA) that can be cleaved by MMP-2. When a scrambled peptide sequence was used, the cleavage efficiency of MMP-2 was markedly reduced. Recently, the same group designed new CAs for imaging tumor MMP-2 activity with a different strategy, 78 which contains a paramagnetic gadolinium chelate (i.e. Gd-DOTA) attached to the N-terminus of the same MMP-2 cleavable peptide sequence via a hydrophobic chain. Since the aqueous solubility of the CA depends on the presence of a PEG chain on the C-terminus of the peptide, MMP-2 cleavage of the peptide detaches the PEG chain from the CA and makes it less water-soluble. Although this CA and control compounds were tested in an animal model bearing two tumors with different levels of MMP-2 activity, strangely, no imaging data was reported. 78
Molecular MRI is in its infancy. The major disadvantage of MRI is the inherent low sensitivity, which can only be partially compensated by working at higher magnetic fields (4.7–14 T in animal studies), acquiring data for much longer time periods, and using exogenous contrast agents. Although proof-of-principle studies have been reported for molecular MRI of a few targets,9,79 whether molecular MRI can significantly improve cancer patient management remains to be elucidated. To overcome the intrinsic low sensitivity of MRI and poor tissue penetration capability of optical imaging, radioisotope-based imaging techniques (i.e. SPECT and PET) should be used if protease imaging is needed in the clinical setting.
SPECT Imaging of MMPs
SPECT imaging detects gamma rays. Internal radiation is administered through inhaling, ingesting, or injecting a low mass amount of radiolabeled pharmaceuticals. A collimator is used to only allow the emitted gamma photon to travel along certain directions to reach the detector, which ensures that the position on the detector accurately represents the source of the gamma ray. The gamma camera can be used in planar imaging to obtain two-dimensional images, or in SPECT imaging to obtain three-dimensional images.80,81 Because of the use of lead collimators to define the angle of incidence, SPECT imaging has a very low detection efficiency (<10–4 times the emitted number of gamma rays).8,82 Common radioisotopes used for SPECT imaging are 99mTc (t1/2: 6.0 h), 111 In (t1/2: 2.8 d), 123I (t1/2: 13.2 h), and 131I (t1/2: 8.0 d).
The proteolytic activities of MMPs are regulated by endogenous tissue inhibitors of MMPs (TIMPs). 83 TIMPs specifically inhibit active forms of MMPs, and in some cases latent MMPs as well, and disturbance in this balance may lead to pathological situations in tissues. Therefore, TIMPs can serve as potential targeting molecules for imaging MMP expression and such a radiopharmaceutical ( 111 In-DTPA-N-TIMP-2) has been reported (Fig. 5). 84 DTPA-N-TIMP-2 (DTPA represents diethylenetriamene pentaacetate) was labeled with 111 In in >95% radiochemical yield, which was quite stable in serum. This tracer has been tested in patients with Kaposi's Sarcoma for SPECT imaging applications. 85 Although the probe was found to be safe, the authors concluded that its tumor targeting/imaging capability was very poor and unlikely to be of use for imaging applications.
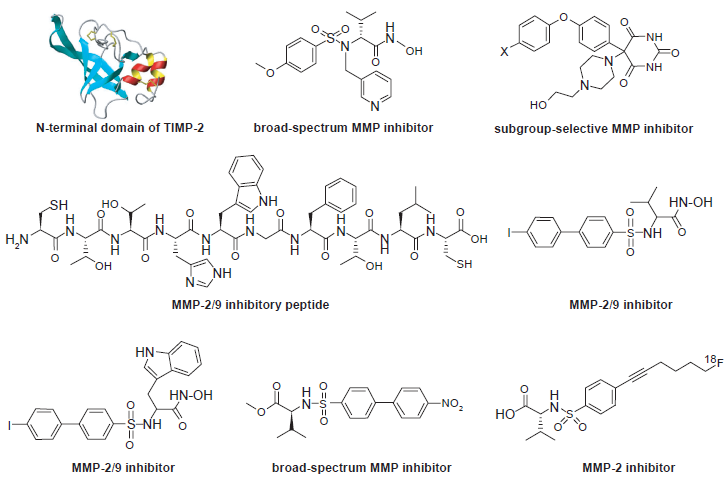
Representative MMPIs that have been radiolabeled for SPECT/PET applications.
A MMP-2/9 inhibitory peptide, CTTHWGFTLC (CTT), was labeled with 125I and 99mTc. 86 It was shown that the radiolabeled CTT peptide inhibited MMP-2 activity in vitro and could home to a MMP-2-positive tumor in mice. SPECT imaging of the tumors in mice was achieved with liposomes coated with 99mTc-labeled CTT. The reason for using liposomes as a carrier can be explained by another report of the 125I-labeled CTT peptide. 87 In this study, it was shown that although the 125I-labeled CTT peptide exhibited interesting properties in vitro for targeting MMP-2/9, its poor solubility and metabolic instability made it unsuitable for in vivo applications.
One intrinsic disadvantage of peptide-based probes is their poor stability in vivo, in particular for linear peptides. Many non-peptidyl MMPIs have been reported in the literature (Fig. 5). Radioiodinated analogs of the non-peptidyl MMPI, CGS 27023A, were synthesized for MMP detection in vivo. 88 In vitro studies revealed that the non-radioactive analogs of the MMPI exhibited affinities against MMP-2 and MMP-9 in the nanomolar range. Biodistribution of the probes in mice showed rapid blood and plasma clearance and low retention in normal tissues. Subsequently, radioiodinated carboxylic and hydroxamic MMPIs were reported. 89 In vitro enzyme assays showed high inhibition capacities of these MMPIs on MMP-2/9. However, the in vivo tumor targeting capability was quite disappointing. Another study of 123I-labeled biphenylsulfonide (a small molecule MMPI) and its analogs in A549 lung carcinoma mice models also revealed that these probes were not suitable as tumor imaging agents. 90 Non-hydroxamate C-5-disubstituted pyrimidine-2,4,6-triones were reported to be subgroup-selective MMP inhibitors. 91 The compound has been labeled with 125I yet in vivo testing has not been reported yet.
To date, SPECT imaging of MMPs in cancer has not been successful. Interestingly, the development of an activatable, MMP-14 targeted SPECT imaging probe was reported recently. 92 Only a modest two-fold reduction of cellular radioactivity was observed in the presence of a broad-spectrum MMPI. In the past, gamma cameras and SPECT imaging systems were much more readily accessible than PET systems. 93 However, PET has significantly higher sensitivity than SPECT and with the continuous developmental effort, state-of-the-art small animal PET scanners can have spatial resolution (<1 mm) comparable to SPECT and they are also becoming increasingly widely available.94,95 Therefore, PET imaging will become more and more dominant over SPECT imaging in the future.
PET Imaging of MMPs
Several PET probes for imaging protease (almost exclusively MMPs) expression in tumors have been reported. Those probes are generally MMPIs that have been labeled with 11 C96–98 and 18 F.99–102 However, in most of these reports, selective binding of the labeled compounds to specific MMPs was not shown, and high non-specific binding was observed possibly due to low in vivo stability of the tracers. Except for one report, 103 none of the abovementioned studies showed any convincing in vivo tumor targeting/imaging results.
The previously mentioned MMP-2/9 inhibitory peptide, CTT, was conjugated with DOTA and labeled with 64 Cu for PET imaging of MMP expression in xenograft models. 103 Zymography of tumor extracts supported the in vivo PET imaging results. MMP-2 and MMP-9 bands were clearly detectable in the mouse imaged at 7 weeks, which had prominent tumor uptake, whereas the MMP-2 and MMP-9 expression was very weak afterwards, which gave low tumor uptake. However, MMP expression in the MDA-MB-435 tumor model used in this study had quite large individual variance. The low in vivo stability of the probe also limits further investigation of this strategy.
Conclusion and Future Perspectives
Molecular imaging of proteases in cancer has been investigated with a wide variety of agents (e.g. small molecules, peptides, proteins, polymers, and nanoparticles) labeled with many different imaging tags (e.g. fluorescent dyes, quantum dots, luciferins, Gd-chelates, magnetic nanoparticles, and radioisotopes) (Table 1). Optical techniques, in particular fluorescence imaging, are the most intensively validated and many of the protease imaging probes are already commercially available. It is also generally agreed that imaging with the “activatable probes” is the most accurate and appropriate means for measuring protease activity. Imaging protease activity with other techniques (i.e. MRI, SPECT, and PET) has not been well-documented in the literature which certainly deserves much effort in the future.
A brief summary of protease imaging in cancer.
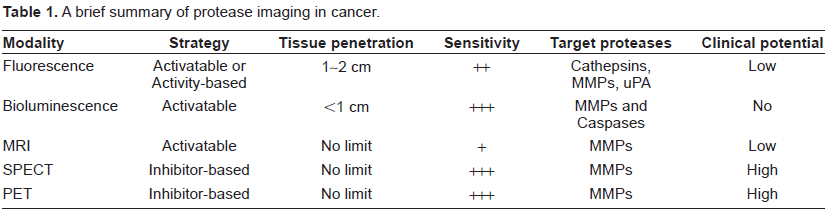
MRI of protease activity in cancer is so far limited to cell-based studies and whether molecular MRI of protease activity has a future in the clinic is questionable. PET/SPECT imaging certainly has the best potential for clinical application. However, suitable probes for imaging protease expression in cancer have yet to be developed. The fact that most of the radiolabeled MMPIs are not specific to a single MMP makes the scenario even more complicated as in many cases the experimental results were hard to interpret due to the presence of too many variables. First, the MMPs targeted in these studies are mostly cell secreted and their expression is known to vary extensively, depending on the stage of the tumor and the presence of naturally occurring TIMPs. Given the lack of data on the levels of MMP expression in the tumor tissue at the time of imaging in most studies reported to date, it cannot be excluded that the negative results reported are, in fact, false-negative imaging results.
Second, most of the agents used for imaging studies are intrinsically broad-spectrum agents. Therefore, their higher affinity for specific subsets of MMPs does not necessarily imply that a positive imaging result corresponds to over-expression of specific subsets of MMPs, as suggested in some reports. Well-characterized tumor models need to be developed for validating the currently available, as well as future, MMP imaging probes. To develop suitable probes for protease imaging in the clinic, inhibitors with better specificity and higher affinity to a certain MMP should be developed.
Although the proof-of-principle has been demonstrated for protease imaging, there are certain limitations/flaws in many of the reported in vivo imaging studies. For successful in vivo imaging applications, the biochemical and cell biological properties of the probes should be demonstrable ex vivo. However, very few of the probes described, in particular the fluorescent probes, have been shown to be optimal even for live cell imaging. In addition, some control studies in these reports were also not very convincing. In vivo imaging is much more challenging and complex than cell-based imaging. In many cases, the following concepts may be intermingled in the interpretation of signal that was observed in vivo: signal intensity vs. specific protease activity, permeability vs. cleavage, and affinity vs. specificity. In future studies, these issues should be evaluated in both ex vivo and in vivo environments to provide more robust results for in vivo protease imaging.
Until recently, clinical trials targeting MMPs have yielded disappointing results, highlighting the need for better insight into the mechanisms by which enzymes contribute to tumor growth.104,105 Molecular imaging to monitor protease expression non-invasively in vivo will be critical for early cancer detection, future protease-targeted drug development, and monitoring the therapeutic efficacy of these drugs. First, non-invasive imaging of proteases can help the clinicians to choose the right timing for protease inhibitor-based therapy. The reason why most MMPIs failed in clinical trials is that they are not administered at the right time. 106 Imaging can help determine the protease expression level at different stages, which can maximize the effect of protease inhibitors during treatment. Second, imaging of proteases can enable accurate monitoring of the therapeutic responses to certain treatment, since protease expression/activity is closely related to cancer progression and many proteases can serve as the target of drug treatment themselves. Lastly, protease imaging can also be used to develop in vitro, cell-based, or small animal assays for drug screening, which can benefit the cancer patients in the long run.
Protease imaging is not only applicable to cancer, it can also be applied in a wide variety of diseases such as arthritis, 107 renal diseases, 108 atherosclerosis,109,110 myocardial infarction, 111 viral infection, 112 among others. In vivo imaging of protease activity, using non-optical techniques (which has good clinical potential), is a field in its infancy but with substantial promise. The next decade is likely to witness dramatic advances in the technologies required to bring novel protease imaging agents to clinical trials.
Disclosure
The authors report no conflicts of interest.
Footnotes
Acknowledgments
The authors acknowledge financial support from the UW School of Medicine and Public Health's Medical Education and Research Committee through the Wisconsin Partnership Program, the University of Wisconsin Carbone Cancer Center, NCRR 1UL1RR025011 and a Susan G. Komen Postdoctoral Fellowship (to H. Hong).