
Abstract
Nonmelanoma skin cancer (NMSC) is a major health concern worldwide. With increasing numbers in high-risk groups such as organ transplant recipients and patients taking photosensitizing medications, the incidence of NMSC continues to rise. Mouse models of NMSC allow us to better understand the molecular signaling cascades involved in skin tumor development in order to identify novel therapeutic strategies. Here we review the models designed to determine the role of the polyamines in NMSC development and maintenance. Elevated polyamines are absolutely required for tumor growth, and dysregulation of their biosynthetic and catabolic enzymes has been observed in NMSC. Studies using mice with genetic alterations in epidermal polyamines suggest that they play key roles in tumor promotion and epithelial cell survival pathways, and recent clinical trials indicate that pharmacological inhibitors of polyamine metabolism show promise in individuals at high risk for NMSC.
Introduction
Nonmelanoma skin cancer (NMSC) is a major health concern worldwide and accounts for 40% of all diagnosed cancers in the United States.1,2 Recent studies estimate a more than 300% increase in NMSC in the United States since 1994, with an annual cost of $650 million. 3 There are two major forms of NMSC, basal cell carcinoma (BCC) and squamous cell carcinoma (SCC), which account for 80% and 16% of all diagnosed skin cancers, respectively. 1 BCCs are slow-growing and rarely metastasize, whereas SCCs are more invasive and metastasize at a higher frequency.1,4 As with most cancers, NMSC prevalence increases with age. It has been estimated that 80% of NMSC cases occur in individuals aged 60 or older.4,5 However, another estimate has shown NMSC to be on the rise in young adults. 5 Overall, a 3%–8% increase in NMSC incidence has been reported worldwide since 1960. 4 These alarming statistics emphasize the importance of studying the mechanisms underlying this disease by implementing physiologically relevant animal models in order to identify novel preventative and therapeutic strategies.
Risks of NMSC
Exposure to ultraviolet radiation (UVR) is the primary risk factor for NMSC, as well as melanoma.1,6 The risk of SCC is increased in fair-skinned individuals, those with increased cumulative sun exposure, those living in geographic locations closer to the equator, and older populations (reviewed by Kim and Armstrong). 7 The 3-year risk of SCC in patients previously diagnosed with SCC is more than 10 times the rate in the general population, 8 and patients previously diagnosed with actinic keratosis (AK), intraepidermal malignancies that exist on a continuum with SCC, are also at increased risk for SCC. 9
In addition to the increasing overall rate of NMSC, there are also specific groups who are at particularly high risk for developing SCC. Xeroderma pigmentosum is an autosomal recessive genetic disease resulting from mutations in the DNA damage repair machinery. 10 These patients have hypersensitivity to UV light and significantly increased (2,000-fold higher) risk of SCC, BCC, and melanoma.10–12 Organ transplant recipients taking immunosuppressive drugs to prevent rejection are at a higher general cancer risk, with SCC being the most common neoplasm. 13 The incidence of SCC is 60- to 100-fold greater in organ transplant recipients than the general population, and frequently patients are diagnosed with multiple SCCs.14,15 Finally, a recent large case–control study found an increased risk for developing SCC and BCC associated with photosensitizing medications, particularly antimicrobials used for skin conditions and thiazide diuretics. 16 These high-risk populations not only provide valuable information about the pathways involved in NMSC development but also highlight the critical need to develop strategies for prevention and therapy.
Mouse Models of NMSC
Multistage chemical carcinogenesis model
The mouse multistage skin chemical carcinogenesis model is one of the most extensively studied in vivo models of epithelial tumorigenesis.
17
This highly reproducible system is seen as a prototypical model for the initiation, promotion, and progression phases of epithelial tumorigenesis and provides the opportunity to study the effects of chemopreventive and chemotherapeutic agents, as well as genetic mutations and dietary manipulations, at various stages of tumor development.
18
Chemical carcinogenesis in mouse skin has been confirmed as a valid tool to study human epithelial cancers, as humans are naturally exposed to low doses of carcinogens and tumor-promoting agents.
19
Furthermore, the model is thought to be relevant to human NMSC development because progression of benign lesions to SCCs occurs in a stepwise manner, and it induces intracellular signaling alterations similar to those produced by UVR exposure. Activating mutations in codon 61 of Ha-
Early predecessors of the multistage chemical carcinogenesis model date back to the 1910s when Yamagiwa and Ichikawa painted rabbit skin with coal tar to induce tumors (reviewed by Marks and Furstenberger).
30
In the 1920s, Deelman discovered that wounding after application of carcinogenic tar caused the development of skin tumors in mice.
31
Friedwald and Rous were the first to define the terms “initiation” and “promotion” when describing skin carcinogenesis. Using rabbits, they demonstrated that tumor cells were initiated by a single treatment of the carcinogen 3-methylcholanthrene. The initiated cells could be promoted into tumors after subsequent treatment of the skin with agents that induce proliferation but were not able to cause neoplastic transformation when applied alone.
32
In the 1940s, the initiation and promotion approach was established in mice. This occurred after the discovery that croton oil was a potent promoting agent. Tumor initiation was achieved by treating mice with a subcarcinogenic dose of carcinogen, most commonly 7,12-dimethylbenz[a]anthracene (DMBA). Promotion occurred via the chronic application of croton oil.33,34 The use of this model revealed that initiation was irreversible and that the sequence of initiation and promotion was not interchangeable.35,36 The discovery that phorbol diesters, particularly 12-
The multistage chemical carcinogenesis model is depicted in Figure 1. The polycyclic aromatic hydrocarbon DMBA is typically applied topically as the initiating agent during the resting phase of the mouse hair cycle (telogen). This application results in mutations to regulatory genes in epidermal keratinocytes by DMBA interacting with DNA and forming N
6
-dAdo DNA adducts.
19
One hallmark of this model is A to T transversions that frequently occur in the Ha
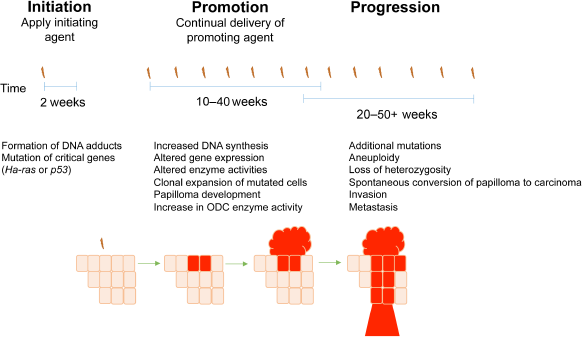
Multistep skin tumorigenesis. The evolution of benign papillomas and squamous cell carcinomas in response to treatment of normal skin with initiating and promoting agents is shown. Important genetic and biochemical changes associated with tumor initiation, promotion, and progression are listed.
During tumor promotion, initiated cells that have acquired a growth advantage clonally expand to form papillomas. Promotion occurs after repeated applications of a tumor-promoting agent, usually TPA. The application of tumor-promoting agents primarily induces biochemical rather than genetic alterations and often leads to skin hyperplasia and increased epidermal thickness. 40 Although the exact mechanism whereby TPA induces skin tumors is not known, the hydrophobicity of the acyl chain in all phorbol esters is critical for their tumor-promoting ability, and phorbol esters have been shown to increase mRNA and protein synthesis. 19 TPA is an analog of diacylglycerol and binds to protein kinase C (PKC), leading to activation of PKC downstream targets, 41 and further work by Verma and colleagues has established the importance of PKC in the development of cutaneous SCC.42–44 While the contribution of chronic inflammation to the development of NMSC is not well understood, it is also known that tumor promoters induce secretion of pro-inflammatory molecules by keratinocytes. This in turn results in the recruitment of inflammatory cells into the dermis, which produce cytokines and chemokines that suppress adaptive immunity and promote tumor growth (reviewed by Rundhaug and Fischer). 45
Progression is defined as the conversion of benign papillomas to carcinomas. During progression, additional genetic mutations occur and chromosomal abnormalities develop, such as aneuploidy. The resulting carcinomas may be both invasive and metastatic. The progression of papillomas to SCCs is strain dependent and occurs stochastically; there is an increased probability of additional genetic alterations as the cell population expands, and mutagen treatment or loss of p53 leads to enhanced progression.46–50 The subsequent conversion of SCCs to spindle carcinomas is a rare event.
UVR model
UVR exposure has been shown to induce oncogenic mutations in epidermal keratinocytes as well as metabolic changes in immune cells within the tumor microenvironment (reviewed by Kim and He).
51
The UV spectrum is divided into UVC (200–280 nm), UVB (280–320 nm) and UVA (320–400 nm). Of these, UVB wavelengths are the most energetic and account for the majority of the biologically damaging effects of sun exposure.
52
Studies in mice have shown that UVA and UVB are complete carcinogens, acting as both tumor-initiating and tumor-promoting agents.53,54 However, UVA is a weak complete carcinogen, functioning as a more potent tumor promoter than initiating agent. UVR is absorbed by macromolecules, inducing direct DNA damage by initiating the formation of cyclobutane dimers, 6-4 photoproducts, cytosine photohydrates, DNA cross-links, and DNA double-strand breaks.
1
If such adducts are not repaired, then classical CC to TT and C to T transitions can ensue during normal DNA replication. Such UVR mutation signatures have been detected at a high frequency in the
Methods described for the mouse UVR model of skin carcinogenesis are more variable than the well-characterized multistage chemical carcinogenesis model. Numerous doses of UVR (usually UVB), time points, and mouse strains have been used to study the progression of NMSC, with no standard procedure being defined.59,60 It can be argued that the UV-induced mouse skin cancer model provides a more physiologically relevant method to study the underlying mechanisms of skin tumorigenesis because the carcinogens used are analogous to those that cause human disease. Moreover, while mutations in both
Polyamine Metabolism
A large number of signal transduction pathways are affected during NMSC development, and several have been the subjects of recent reviews.63–65 This review concentrates on the role of the polyamines in NMSC development and maintenance.
Polyamines, small ubiquitous polycations, are essential for normal cell growth and development,66–68 yet elevated levels of the polyamines and polyamine biosynthetic enzymes are associated with epithelial carcinogenesis and neoplastic growth.69–74 Under normal physiological conditions, intracellular polyamine levels are tightly regulated by complex metabolic, catabolic, and poorly understood transport mechanisms (Fig. 2).69,73,74 Due to the cationic nature of the polyamines, they interact with anionic molecules such as proteins, DNA, and mRNA and can modify diverse processes such as chromatin and DNA structure, DNA damage, histone acetylation, transcription, mRNA processing, stability and translation, cell cycle progression, kinase activity, and ion channel function.70,74–79 Changes in polyamine content can also induce selective rather than global effects on DNA–protein interactions 80 and the translation, 81 levels, 82 or stability 83 of specific transcripts.
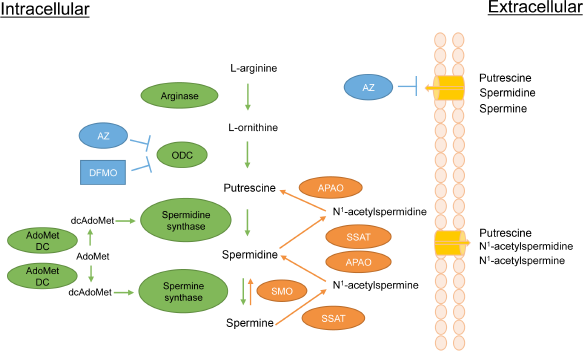
The polyamine pathway in mammalian cells. Details of the pathway are described in the text. Metabolic enzymes are in green while enzymes of polyamine catabolism are in orange. ornithine decarboxylase (ODC) inhibitors α-difluoromethylornithine (DFMO) and antizyme (AZ) are in blue. In addition to inhibiting ODC, AZ causes the ODC protein to be degraded and has the additional effect of inhibiting polyamine transport by an unknown mechanism. other abbreviations are as follows:
In mammals, polyamines are synthesized from the amino acids l-methionine and l-arginine. l-arginine is metabolized into l-ornithine by arginase, and ornithine is subsequently decarboxylated by the rate-limiting enzyme ornithine decarboxylase (ODC) to produce the diamine putrescine. Putrescine is converted to the higher polyamines spermidine and spermine by spermidine synthase (SpdS) and spermine synthase (SpmS), respectively. The rate-limiting enzyme for the synthesis of spermidine and spermine is
Both ODC and AdoMetDC activities are essential for cell growth and proliferation in a vast number of experimental models, and knockout mouse alleles for either gene are lethal at extremely early stages of embryonic development.67,87 ODC is strongly induced by proliferative stimuli and is often upregulated in cancer by various transcriptional, post-transcriptional, translational, and stability mechanisms.88–90 AdoMetDC is also a highly inducible enzyme that exhibits increased activity in association with growth-promoting stimuli, 91 and it has been considered as a therapeutic target in cancer.92–94
The ODC enzyme is active as a homodimer and has a short half-life ranging from 10 to 30 minutes. 90 The degradation of ODC is unique among short-lived proteins in that it is ubiquitin independent. 95 Instead, the monomeric form of ODC associates noncovalently with a protein known as antizyme (AZ), which directs the ODC protein to the 26S proteasome for degradation. 95 The AZ family consists of at least three differentially distributed proteins, all of which can regulate ODC. 90 AZ1, the best-characterized AZ family member, is synthesized in a polyamine-dependent manner. Increases in cellular polyamine levels stimulate a +1 frameshifting event in the translation of the AZ1 mRNA, thereby increasing the expression of functional AZ1 protein. 96 AZ1 not only enhances ODC degradation but also inhibits ODC activity and suppresses polyamine uptake.90,97,98 Thus, AZ1 acts as a multifunctional negative regulator of intracellular polyamine content.
Polyamine Catabolism
In addition to the highly complex biosynthetic pathway described above, the polyamines are regulated by an equally complex catabolic pathway. The relative susceptibility of spermine and spermidine to degradation/excretion is controlled by the activity of Spermidine/spermine N1-acetyltransferase (SSAT). SSAT catalyzes the formation of N1-acetylspermine or N1-acetylspermidine by transferring the acetyl group from acetyl-coenzyme A to the N1 position of spermine or spermidine. 99 These acetylated polyamines can be either exported or can serve as substrates for the flavin-dependent N1-acetylpolyamine oxidase (APAO). APAO converts the acetylated polyamines to spermidine or putrescine, depending on the substrate, as well as 3-aceto-aminopropanal and hydrogen peroxide.100–102 Although APAO may also be inducible in some cases,73,102 SSAT, which is induced by a wide range of stimuli including toxins, heat shock, hormones, and polyamines (reviewed by Casero and Pegg), 85 represents the rate-limiting step in this retroconversion. Acetylation of polyamines also leads to their efflux from the cell. 73 SSAT induction therefore leads to the reduction of polyamine levels via their conversion to putrescine and the excretion of putrescine, N1-acetylspermidine, and N1-acetylspermine. The enzyme spermine oxidase (SMO) is able to oxidize nonacetylated spermine directly to spermidine, creating the byproducts aldehyde-3-aminopropanal and hydrogen peroxide. 103 SMO is induced by several antitumorigenic polyamine analogs in cell lines derived from lung, breast, colon, and prostate tumors, suggesting that SMO is a possible target in malignancies.104,105
Polyamine Transport
As previously mentioned, intracellular polyamine content is regulated by a poorly characterized transport mechanism. Although this system is not well understood, it is evident that the polyamine transport system (PTS) plays a critical role in maintaining a specific range of cellular polyamines. The PTS is upregulated in response to polyamine depletion,106,107 and AZ has been shown to repress both ODC enzyme activity and the PTS. 106 However, it has been hypothesized that this repression of the PTS is not through AZ, but via changes in the abundance of an unidentified polyamine permease. 107 It is clear that additional research is needed in order to better identify the link between ODC, AZ, and the PTS. Moreover, the identification of the elusive polyamine transporter and its components is crucial for designing drugs that can target the polyamine pathway in diseases where it is dysregulated.
The Polyamine Pathway in NMSC
Understanding the role of the polyamine pathway in skin carcinogenesis is of great importance to the design of agents for both chemoprevention and chemotherapy of NMSC as well as epithelial tumors in general, and much progress has been made in this field. Using the mouse skin carcinogenesis model, studies by Boutwell and colleagues were the first to link the activity of ODC to cancer of any kind. These early studies showed that TPA induces a massive transient increase in epidermal ODC activity along with a more prolonged moderate increase in AdoMetDC activity.91,108,109 ODC activity and polyamine content are constitutively elevated in DMBA/TPA-induced tumors.108,110,111 Studies utilizing either the multistage chemical carcinogenesis model or UV-irradiation have demonstrated that the upregulation of ODC is necessary for the onset of skin tumors in mice.91,112–114 Mice treated with DMBA/TPA or UVB, including
In addition to their essential role in cellular proliferation, the polyamines have also been linked to suppression of both innate and adaptive immunity in many systems, which could provide a mechanism by which tumors evade the immune response. Recent studies have shown that both spermidine and spermine reduced neutrophil infiltration in TPA-treated skin and suppressed IL-1β and tumor necrosis factor alpha (TNF-α) production in macrophages. 119 Moreover, simultaneous inhibition of ODC and polyamine transport suppressed skin tumor growth in immunocompetent mice but not in athymic nude mice lacking T-cells, suggesting that polyamine-targeted therapies may reverse tumor immunosuppression. 120 These data combined with the results above indicate that the polyamine pathway is a valid target in NMSC development, and some of the pivotal genetic mouse model studies designed to support this conclusion are highlighted in the text below and summarized in Table 1.
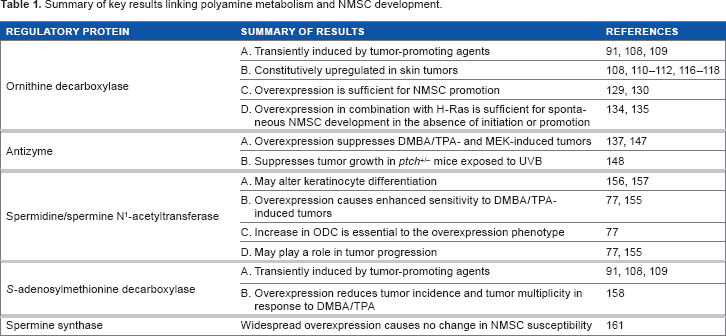
Summary of key results linking polyamine metabolism and NMSC development.
Mice with Genetically Altered Polyamines as Models to Probe NMSC Biology and Pathogenesis
In considering an in vivo model to test the biologic effects of genetically manipulating polyamine metabolism, mouse models in which transgenes are driven by keratin promoters offer the key advantage of tissue targeting based on the known expression pattern of keratin genes.121,122 The keratin 5 (K5), keratin 14 (K14), and keratin 6 (K6) promoters direct expression of transgenes to basal keratinocytes of the hair follicle outer root sheath (ORS) and the interfollicular epidermis.123,124 These regions include the critical follicular stem cell niche that supports hair follicle development and wound healing as well as the interfollicular stem cell niche that plays an important homeostatic role, and both niches are implicated in skin tumor development.49,125–127 The K5 promoter and its structural partner K14 are constitutively co-expressed in the ORS and interfollicular epidermis, while K6 expression is constitutive within the ORS and induced in the interfollicular epidermis by proliferative agents such as TPA. 128 Thus, these models allow one to modify important signal transduction pathways in a small population of cells and determine what effect these genetic alterations have on tumor development and maintenance of established tumors. Many of the models discussed in this section use these promoters to direct expression of transgenes designed to modify levels of epidermal polyamines.
Ornithine decarboxylase
Transgenic mice overexpressing ODC in hair follicle keratinocytes using keratin promoters (K6-ODC mice and K5-ODC mice) were shown by O'Brien and colleagues to be much more sensitive than littermate controls to DMBA-induced carcinogenesis and did not require treatment with a tumor promoter to develop tumors, suggesting that ODC overexpression is a sufficient promoting stimulus in this model.129,130 Interestingly, it was further shown that papillomas from DMBA-treated K6-ODC mice exhibit an increased frequency of activating mutations in K-
Results from the transgenic models described above have established ODC as an important factor in tumor promotion. However, the role of ODC in the later stages of tumorigenesis is less well defined. Polyamine depletion using DFMO has been shown to cause regression of both papillomas and SCCs in several transgenic models,42,135,136,138 as well as a reduction in carcinoma vascularization.
135
These studies point to putrescine levels as an important regulator of tumor growth, but the molecular mechanisms are not known. DFMO-regressed tumors in K6-ODC/Ras mice showed no change in
Antizyme 1
As discussed above, AZ1 is an important endogenous regulator of ODC and polyamine homeostasis (Fig. 2). A critically important feature of AZ1 is its ability to suppress polyamine uptake, since the upregulation of polyamine transport represents one of the major limitations to the effectiveness of DFMO as a chemotherapeutic agent.69,70 In addition, studies in animal models 144 and humans145,146 demonstrated a loss of AZ1 expression or activity in tumors relative to normal tissue.
To determine whether suppression of ODC activity and polyamine uptake blocks the promotion of initiated target cells, we have used K5 and K6 promoter elements to drive expression of AZ1. These transgenic lines target inhibition of ODC activity to a specific subpopulation of epidermal keratinocytes, rather than the more general systemic effect seen with an inhibitor such as DFMO. K5-AZ1 and K6-AZ1 mice exhibit a substantial delay in tumor onset and a significant reduction in tumor multiplicity in response to both DMBA/TPA-induced chemical carcinogenesis and when crossed with K14-MEK mice, indicating AZ1 can act as a tumor suppressor in skin carcinogenesis.137,147 AZ1 was also found to suppress tumor growth in a UV carcinogenesis model that utilized mice heterozygous for
Spermidine/spermine-N1-acetyltransferase
It was originally hypothesized that genetic manipulations to enhance SSAT activity would decrease tumor susceptibility by depleting the higher polyamines spermidine and spermine. The importance of SSAT in maintaining polyamine content was shown by studies using transgenic mice with constitutive and ubiquitous overexpression of SSAT under its own promoter (SSAT 165 mice).149,150 These mice show a marked alteration in polyamine content with large increases in putrescine and declines in spermidine and spermine in multiple tissues, as well as permanent hair loss by age 3 weeks and development of large dermal cysts. Interestingly, an identical phenotype was described in K6-ODC mice, 151 which accompanies an increased susceptibility to skin tumorigenesis, while SSAT 165 mice were resistant to DMBA/TPA. 149 However, SSAT 165 mice also suffered from severe metabolic defects.152–154
In contrast to original predictions, mice with epidermal overexpression of SSAT (K6-SSAT mice) developed significantly more and larger tumors compared to controls in response to DMBA/TPA, and these tumors were much more likely to convert to SCCs. 155 This phenotype was linked to both increased putrescine levels and oxidative damage resulting from SSAT-stimulated polyamine catabolism. 77 ODC activity increased in response to spermine and spermidine depletion, and DFMO treatment as well as crosses with K6-AZ1 mice demonstrated that this increase in ODC was essential to the K6-SSAT phenotype. 77 Limited clinical studies have also linked SSAT induction to human skin disease. A family affected with keratosis follicularis spinulosa decalvans, a rare X-linked syndrome causing follicular hyperkeratosis, demonstrated duplication of the X-chromosome in the region containing SSAT as well as a three-fold increase in SSAT activity, accompanied by increased putrescine and decreased spermidine pools. 156 This phenotype is consistent with alterations in keratinocyte differentiation, which were also observed in organotypic keratinocyte cultures from SSAT-overexpressing mice. 157
AdoMetDC and SpmS
Of the polyamine biosynthetic enzymes, ODC has received the vast majority of attention in cancer studies since the initial report 40 years ago that TPA induces both ODC and AdoMetDC activity in mouse skin. To address this knowledge gap, we recently utilized a tetracycline-inducible system to achieve regulated AdoMetDC activity in mouse skin (TetO-AdoMetDC transgene (TAMD) mice). 158 This was the first transgenic model to be produced that allowed manipulation of cellular AdoMetDC content in a tissue-specific and regulated manner. Given that AdoMetDC activity is rate limiting for the biosynthesis of spermidine and spermine, it was predicted that these animals would have increased tumor susceptibility if these higher polyamines drive carcinogenesis. Interestingly, upon DMBA/TPA chemical carcinogenesis, TAMD mice exhibited significantly reduced tumor incidence (percent of mice with tumors) and tumor multiplicity (number of tumors per mouse) than controls. Furthermore, latent initiated cells persist in the skin of these mice despite the lack of macroscopic tumor formation. This novel finding was demonstrated by showing that upon silencing of AdoMetDC expression, the tumor multiplicity of TAMD mice rapidly increased to levels that are nearly equivalent to those in control animals. 158
Taken together with the results from ODC, AZ1, and SSAT models discussed above, studies using TAMD mice strongly support the concept that high levels of intracellular putrescine are critical for tumor promotion of initiated keratinocytes. This is in contrast to the effects seen in normal keratinocytes, where high ODC has been shown to elicit apoptosis,
159
perhaps through generation of reactive oxygen species caused by induction of APAO/SMO and activation of ataxia telangiectasia mutated (ATM)-DNA damage signaling.
160
These studies suggest that initiated keratinocytes respond differently to elevated putrescine. Further support for putrescine as a tumor-promoting agent was provided by findings in mice with widespread overexpression of SpmS driven by a composite cytomegalovirus-immediate early gene enhancer/chicken β-actin promoter (CAG-SpmS mice). These studies showed that elevated SpmS activity, and the resulting increase in the spermine:spermidine ratio, do not increase susceptibility to either skin chemical carcinogenesis or to spontaneous intestinal carcinogenesis in
Prospects for Polyamine-based Therapy in NMSC
DFMO is already approved by the US Food and Drug Administration for clinical use to prevent unwanted facial hair 162 and is undergoing intensive development as a chemopreventive agent,15,163–167 where it has shown promising efficacy in phase IIb and III clinical trials for BCC and SCC in patients with a previous history of skin cancer.168,169 The topical administration of DFMO to patients with existing AKs also significantly reduced the number of lesions.170,171 The strong chemopreventive effects of DFMO, along with minimal toxicity, reinforce the idea that polyamines are more essential for the growth and survival of tumor cells than their normal counterparts. These data support the use of DFMO as a chemopreventive agent for NMSC and underscore the relevance of the polyamine pathway in skin tumor development. Evidence from animal models using DFMO in combination with polyamine transport inhibitors172,173 or cyclooxygenase (COX) inhibitors174,175 demonstrated increased efficacy in both chemoprevention and therapy of NMSC. In related clinical trials, the combination of DFMO and the COX inhibitor sulindac was both safe and remarkably effective in preventing recurrence of colorectal adenomas in resected adenoma patients followed-up for 3 years, 176 suggesting this would also be a promising combination in individuals at high risk for NMSC such as organ transplant recipients or Xeroderma pigmentosum (XP) patients. Patients with XP in particular are essentially “initiated”; therefore, polyamine-targeted “antipromotion” therapy may be ideal in this population.
Conclusions and Outlook for Future Studies
Studies using the multiple mouse models designed to alter intracellular polyamines in keratinocytes have suggested that the polyamine pathway is a valid target in human NMSC. However, several hurdles remain that have so far resulted in limited therapeutic success. A greater understanding of polyamine transport is essential to design of polyamine-directed therapies and it has been a key challenge in the successful use of DFMO as a chemotherapeutic agent. As mentioned above, the combination of DFMO and a polyamine transport inhibitor was significantly more effective than DFMO alone in a mouse skin SCC model172,173 as well as a xenograft study with human breast cancer cells. 177 In addition, continued studies using TAMD mice will inform the use of AdoMetDC inhibitors either alone or in combination with DFMO in a number of cancers, and similar models with spatial and temporal control of ODC and AZ will broaden the scope of questions that can be addressed regarding metabolic regulation of the polyamines in a variety of diseases. Future development of similar mouse models manipulating epidermal SMO levels would aid in evaluating the preclinical use in NMSC of polyamine analogs that have shown antitumor properties in other epithelial cancer types. Since polyamine catabolism through either SMO or SSAT/APAO may potentially contribute to oxidative DNA damage as well as putrescine accumulation, these enzymes provide additional promising targets for both prevention and therapy of NMSC. Future studies will undoubtedly employ more advanced genetic engineering technologies that allow precise control of either gene overexpression or gene deletion. Utilization of these models will enable definitive analysis of stage-specific effects of the various proteins discussed in this review, thereby increasing our ability to determine critical factors in tumor initiation, promotion, progression, and maintenance.
Author Contributions
Wrote the first draft of the manuscript: SLN. Contributed to the writing of the manuscript: SLN, DJF, LMS. Agree with manuscript results and conclusions: SLN, DJF, LMS. Jointly developed the structure and arguments for the paper: SLN, LMS. Made critical revisions and approved final version: DJF. All authors reviewed and approved of the final manuscript.