
Abstract
Esophageal cancer is the eighth leading cause of cancer and the sixth most common cause of cancer-related death worldwide. Despite recent advances in the development of surgical techniques in combination with the use of radiotherapy and chemotherapy, the prognosis for esophageal cancer remains poor. The cellular and molecular mechanisms that drive the pathogenesis of esophageal cancer are still poorly understood. Hence, understanding these mechanisms is crucial to improving outcomes for patients with esophageal cancer. Mouse models constitute valuable tools for modeling human cancers and for the preclinical testing of therapeutic strategies in a manner not possible in human subjects. Mice are excellent models for studying human cancers because they are similar to humans at the physiological and molecular levels and because they have a shorter gestation time and life cycle. Moreover, a wide range of well-developed technologies for introducing genetic modifications into mice are currently available. In this review, we describe how different mouse models are used to study esophageal cancer.
Introduction
Esophageal cancer (EC), regardless of its histological subtype, is one of the most common causes of cancer-related deaths worldwide.1,2 Therefore, a better understanding of the pathogenesis of this deadly disease is urgently needed. EC can be divided into two major histological subtypes: esophageal squamous cell cancer (ESCC), which is the most common histological subtype in the world, and esophageal adenocarcinoma (EAC), which represents the most rapidly increasing cancer in Western countries.1,3
ESCC occurs in the middle and lower esophagus. Major risk factors include alcohol consumption, tobacco use, and dietary factors (eg, vitamin deficiency, high nitrite levels, and/or nitrosamine consumption). Conversely, EAC occurs primarily in the distal esophagus. The etiology includes gastroesophageal reflux disease, which can lead to complications such as Barrett's Esophagus (BE), a condition defined by the replacement of the normal stratified squamous epithelium of the esophagus by an intestinal-like columnar epithelium. 4 Obesity is another risk factor associated with EAC. Although the etiologies of both types of ECs differ, the pathogenesis of both types has been linked to inflammation.5–8
EC is usually aggressive and invasive in nature, and thus, many patients have a poor prognosis. The overall 5-year survival rate for patients with EC ranges from 15% to 25%.9,10 Better outcomes are correlated with early diagnosis, and poor outcomes are associated with the presence of metastases.9,10 EC is considered a “silent” cancer. Most patients are diagnosed at a late stage due to lack of symptoms in the early stages of the disease.11,12 Hence, the molecular mechanisms that promote esophageal carcinogenesis are not yet fully understood, especially for the early stages of pathogenesis of this disease. Locally advanced disease is generally treated with surgical approaches, whereas advanced (metastatic and disseminated) and/or recurrent disease is treated with chemotherapy and local therapies, such as radiotherapy or endoscopic palliation. The management of EC is well summarized in a few reviews.9,10 Mouse models have permitted the modeling of human diseases and the testing of therapeutic approaches in a manner that is not possible in human subjects. 13 Establishment of adequate mouse models of EC is necessary for exploring the mechanisms of esophageal tumorigenesis. Mouse models of EC have provided important insights into the biology of the histological subtypes of EC. In this article, we review various mouse models of ECs, including xenograft models, orthotopic models, genetically engineered mouse models (GEMMs), and chemical-induced or diet-induced mouse models (Table 1). For each model, we discuss the contributions and limitations, as well as highlighting the opportunities for further research in the field (Table 1).
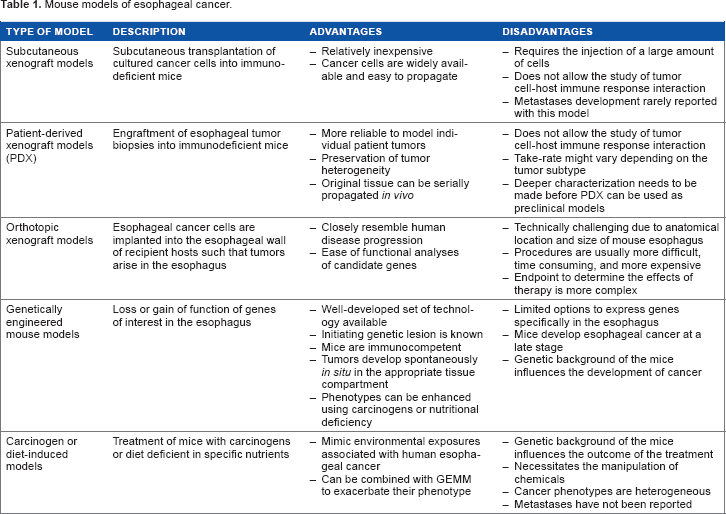
Mouse models of esophageal cancer.
Histology of the Esophagus
The esophagus consists of four primary layers: mucosa, submucosa, muscularis propria, and adventitia. Unlike other areas of the gastrointestinal tract, the esophagus does not contain a serous membrane (serosa). Therefore, esophageal tumors can spread more easily, which makes them more difficult to treat surgically. 14 The stratified epithelium of the esophageal mucosa is composed of three compartments: the basal layer (stratum basale), suprabasal layer (stratum intermedium or prickle), and superficial layer (stratum superficiale). 15 Cells proliferate in the basal layer of the esophagus and undergo differentiation as they migrate through the suprabasal layer to the superficial layer. Esophageal stem cells are located in the basal layer of the epithelium. 16 The submucosa contains connective tissue, immune cells, nerve cells, vascular network, and small mucous glands. The muscularis propria of the upper esophagus is predominantly composed of skeletal muscle, which gradually transitions to predominant smooth muscle in the distal esophagus. Between the upper and lower portions of the esophagus, the muscularis propria is composed of a mixture of both muscle cell types. The esophagus is covered by the adventitia, which is an external fibrous layer. One important histological difference between mouse and human esophagi is that the epithelium is keratinized in mice but not in humans. 15
Mouse Models of EC
Subcutaneous xenograft models
The subcutaneous xenograft model has been used extensively in EC research and is generated by the transplanting of cultured cells derived from human esophageal tumors into immunodeficient mice. In addition to being relatively inexpensive, it allows direct tumor growth assessment.17,18 Moreover, because cancer cell lines are easy to propagate, there is an unlimited supply of cells that are available for use in mouse models. 18 However, this approach also has limitations. 19 For example, xenograft models usually require an injection of a large number of cells into the host and hence fail to mirror the development of human tumors that arise from a limited number of cells. 18 Additionally, the use of immunodeficient mice does not allow for the characterization of study of tumor cell-host immune response interactions, which is a critical component of tumor development and metastasis formation. 20 Finally, few studies of subcutaneous xenograft models of EC reported the development of distant metastases, a common component of EC. 21 However, whether subcutaneous xenograft models of EC cannot lead to metastases is open for discussion. It is possible that some studies have focused solely on primary tumors and missed the formation of metastases, because of long-distance metastases, though it is also possible that distant metastases cannot develop in this model due to regulations stating that the tumor volume must not exceed 1 cm or because of long-distance metastases.3,21
Xenograft models of ESCC and EAC have been published extensively since the early 1980s. In 1983, Robinson and Bux 22 reported the successful establishment of subcutaneous xenograft tumors using human esophageal carcinoma cell lines. Because the number of studies using subcutaneous xenograft models is extremely high and because numerous cancer cell lines have been generated and used in these models, we do not discuss this model further in this review; instead, we discuss the mouse models of EC that have been emerging.
Because the use of subcutaneous xenograft models derived from cancer cell lines has limitations, researchers have redirected their efforts toward developing patient-derived xenografts (PDX), which are developed by implanting a patient's tumor biopsy into an immunodeficient mouse immediately following surgery. 23 PDX models offer more reliability for the study of human EC compared with other xenograft models because they retain the architecture and stromal components of the original tumor and because they preserve tumor heterogeneity. Moreover, the original tissue that is used to generate PDXs can be serially propagated in vivo. Despite the advantages of PDXs, they also have their limitations, particularly, that an immunodeficient host is still required. 23
Very few studies have reported the successful generation of xenograft models derived from patients with ESCC and EAC. Unfortunately, most of these studies have focused only on the feasibility of the method. The first successful transplantation of an esophageal tumor into nude mice was reported in 1981. 24 Tumors were obtained from 36 patients with EC (34 ESCC, 1 EAC, and 1 undifferentiated carcinoma), 24 of which were primary tumors and 14 of which were metastatic tumors. Tumor growth was observed in 44% of the specimens engrafted to nude mice. 24 No correlation was observed between the differentiation stage of the tumors and the rate of engraftment. Another group used xenografts generated from human EAC to investigate DNA copy number changes. 25 Five tumors that were classified as intestinal-type adenocarcinomas were resected and transplanted into nude mice, and four of the specimens showed the presence of BE. Similarly, primary EACs and corresponding lymph node metastases were successfully transplanted into nude mice, with an engraftment success rate of 32%. 25 Recently, PDXs of EC were used to investigate whether engraftment correlates with clinicopathological features and to test whether they could be used for the pharmacologic evaluation of new therapies for EC.26,27 Although engraftment was feasible for all disease subtypes and histological differentiation stages, improved engraftment rates were associated with poor differentiation and lack of neoadjuvant chemoradiation. 27 The majority of the engrafted PDXs resembled their parental tumor (P0) and remained stable after multiple passages at both the histological and the molecular levels.26,27 Moreover, intratumoral heterogeneity was observed in both PDXs and their corresponding parental tumor. Interestingly, mRNA abundance profiling showed global differences during xenograft establishment (P1 versus P0), which can be explained by the replacement of human stroma with mouse fibroblasts. 26 This last observation is a common concern for PDXs in general. 23 Nonetheless, gene expression remained relatively stable between primary tumors and early-passage PDXs. The evaluation of different therapies on cell lines derived from PDX tumors showed a wide range of responses to standard chemotherapy, which, in some cases, led to resistance upon recurrent treatment. This observation recapitulates a challenge that is often observed in the clinic. Taken together, these studies demonstrate the feasibility of PDX models for EC, but a deeper characterization of these models needs to be made before they can be used as preclinical EC models.
Orthotopic xenograft models
The use of orthotopic models has increased over the past decade. Because orthotopic models resemble human ESCC disease progression more closely and frequently form distant metastases, they are considered to be a better option for studying EC than subcutaneous models.28,29 However, the establishment of this type of cancer model is extremely challenging technically due to the anatomical location and the size of the mouse's esophagus. Moreover, the accurate analysis of tumor progression in orthotopic studies is significantly more challenging than for subcutaneous xenograft models because it requires imaging. 28 Most studies commonly use survival rate and body weight to evaluate tumor progression in orthotopic models, which can be very inaccurate because many other factors can influence therapeutic efficacy. To circumvent this limitation, many experts in this field are now using tumor cells marked with a luciferase reporter gene combined with bioluminescent imaging, which allows researchers to monitor tumor growth non-invasively in vivo. 30
Esophageal squamous cell cancer
Few studies have described the generation of orthotopic xenograft models of ESCC in mice. The advanced surgical techniques that are required to generate such models due to the size and the anatomical location of the mouse esophagus might explain why, to date, only a few groups have been successful. The first orthotopic xenograft model of EC was reported in 1990 and was used to test the effect of erbstatin, an inhibitor of epidermal growth factor receptor (EGFR), on the growth of four esophageal tumors.
31
In another orthotopic xenograft model of ESCC, EC cells were injected into the submucosal space of the abdominal esophagus directly after opening the anterior wall of the stomach and performing a laparotomy.
32
Although primary growth was substantial, no metastases were observed. Another group used a different method and injected EC cells (TE-4 and TE-8 cells) into the esophageal lumen through the mouth. In this model, tumor growth occurred both inside and outside the esophageal lumen, with invasion to the dominant nerve, and the mice showed signs of dysphagia.
33
Another study used a noninvasive semiquantitative bioluminescent imaging method to monitor the development and progression of ESCC.
34
In this model, TE-8 EC cells suspended in Matrigel were implanted orthotopically into the subserosal space of the abdominal esophagus of nude mice without opening the stomach. Mice developed tumors at the implantation site, and local lymph node metastases and peritoneal disseminations were observed within 6 weeks after inoculation. However, there was no evidence of dysphagia or significant weight loss in these mice.
34
The use of immunodeficient mice is a limitation for this model as well. A similar injection technique was used to generate the first syngeneic orthotopic model of EC, which was used to test an adenoviral-based immunotherapy.
35
In this study, an EC cell line established from
Esophageal adenocarcinoma
A highly metastatic orthotopic model of EAC using the highly aggressive EC cell lines OE-19 and PT1590 was generated in NMR/nu mice. In this model, metastatic spread was detected in the liver and lungs and in the lymph nodes. This model was also optimized to use high-resolution imaging with green fluorescent protein and magnetic resonance imaging, which can be very useful for the noninvasive evaluation of therapeutic responses for the treatment of EC. 38 The potential of this orthotopic model of EAC as a preclinical model was demonstrated recently in a study evaluating the chemotherapeutic effects of a therapy targeted against HER-2, a member of the EGFR family, and therapy using the C-X-C motif chemokine receptor type 4 (CXCR4) antagonist CTCE-9908.39,40 A detailed protocol describing the generation of this mouse model using OE-19 and PT1590 cells is available in the literature. 41
Genetically engineered mouse models
GEMMs have been used extensively during the past decade to study cancer biology. They include transgenic mouse models, gene knockin and knockout models, and conditional/inducible models. The well-developed set of technologies available for genetic engineering allows researchers to control gene expression in almost any desired tissue and at any time point. The methods and methodologies most commonly used in generation of GEMMs have been described in detail in previous publications.13,42–45
Both ESCC and EAC have been extremely difficult to genetically model in vivo (Table 2). This might be explained in part by the histological differences between mouse and human esophagi. In the human esophagus, the gastroesophageal junction forms the border between the distal esophagus and the proximal stomach and correlates with the transition from the squamous epithelium of the esophagus transitions into the columnar epithelium of the gastric cardia. In the mouse esophagus, the transition from squamous epithelium to columnar epithelium occurs in the stomach. 46 Moreover, the esophageal epithelium is nonkeratinized in humans but is keratinized in mice. The presence of keratinization in mouse esophagus is thought to make it more resistant to injuries and to the development of EC. Another factor that could explain the difficulties in genetically modeling EC in vivo is the time course of ESCC development in humans, which peaks around the age of 70 years. 47
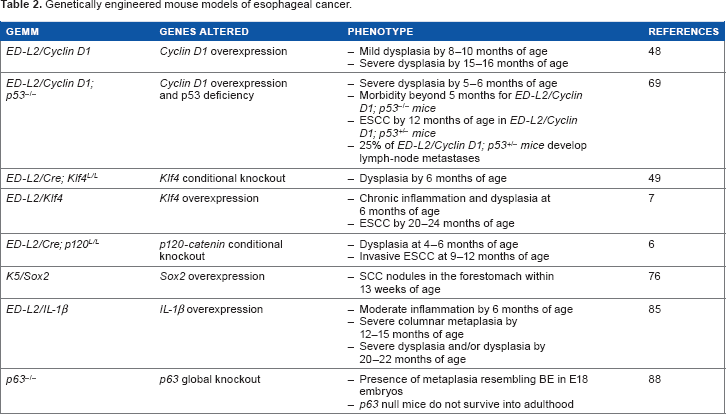
Genetically engineered mouse models of esophageal cancer.
Different tissue- and cell-specific promoters have been used to study esophageal functions. The
Esophageal squamous cell cancer
ESCC is a multistep process that is caused by the accumulation of multiple genetic alterations, such as overexpression of EGFR and cyclin D1, inactivation of p16 and p53, and alterations in the Wnt and Notch pathways.58–60 GEMMs have helped improve understanding of the pathogenesis of ESCC (Table 2). Gene amplification and the overexpression of cyclin D1 occurs in EC patients.61–63 Moreover, constitutively active nuclear cyclin D1 mutants have been identified in patients with ESCC.64,65 The overexpression of cyclin D1 in esophageal epithelia of mice using the
Esophageal adenocarcinoma
Chronic inflammation has been linked strongly to the pathogenesis of BE and EAC.77–79 Until recently, only surgical animal models of esophagojejunostomy existed to induce BE and EAC.80–82 However, this model can be challenging to reproduce in mice and is therefore primarily done in rats. Given that genetic engineering technologies are still more advanced and widely used in mice, this major limitation has restricted our understanding of the origins and molecular pathogenesis of BE and EAC.
Fortunately, a new genetic mouse model of BE and EAC was generated several years ago (Table 2). The proinflammatory cytokine interleukin (IL)-1β is increased in patients with esophagitis, BE, and EAC.83,84 Therefore, the
Chemical- or diet-induced models
Carcinogen treatments or diets have been used to develop mouse models that exhibit phenotypes similar to those of human ESCC and EAC (Table 3). Numerous factors can influence the susceptibility to chemical carcinogen-induced cancers and the resultant tumor incidence and multiplicity, including the dose and schedule of the carcinogens, as well as the age and mouse strain used. 17 In some cases, a combination of carcinogens and genetic engineering has been applied to facilitate the development of EC.
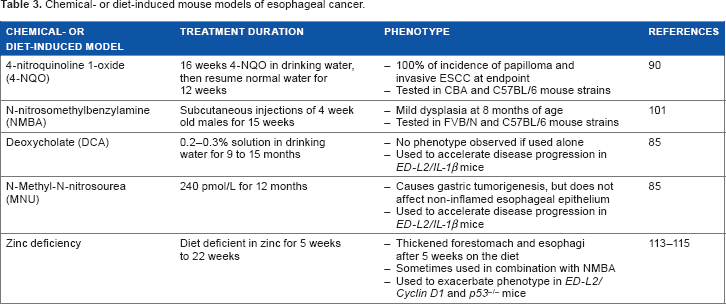
Chemical- or diet-induced mouse models of esophageal cancer.
Esophageal squamous cell cancer
Following ingestion of nitrates and nitrites, anaerobic bacteria generate N-nitroso compounds (NOCs) in the stomach. NOCs are carcinogenic to animals and may play a role as cancer inducers.
89
4-Nitroquinoline 1-oxide (4-NQO) is a quinoline derivative that has been used extensively to study oral and esophageal carcinogenesis.
90
4-NQO causes DNA damage, which includes DNA adducts, single-strand breaks, abasic sites, pyrimidine dimers, and oxidized bases.
91
Thus far, most studies have used 4-NQO painting of tongues in mice to study oral cavity tumorigenesis.90,92–94 When delivered in the drinking water of mice, 4-NQO treatment mimics many aspects of human ESCC.90,95 Notably, the increased expression of keratin-14, keratin-1, and EGFR was reported in esophageal tumors from mice treated with the carcinogen.
90
Reduced expression of the cell cycle inhibitor p16 was also observed in tumors from mice treated with 4-NQO.
90
In this study by Tang et al, 100% of c57BL/6 or CBA mice developed esophageal tumors by 28 weeks after the start of the experiment (16 weeks of 4-NQO at a concentration of 100 μg/mL and then resume normal water for 12 weeks).
90
Few studies have used 4-NQO treatments on GEMMs to determine the influence of genetic factors in the development of ESCC. A number of genetically engineered mice show increased susceptibility to 4-NQO-induced esophageal carcinogenesis, such as transgenic mice with Cyclin D1 overexpression
96
and miR-31 overexpression,
97
as well as Nrf2 knockout mice.
98
Increased tumor formation is also observed in dominant-negative p53-mutated mice [p53(Val135/WT)] that are given 4-NQO in their drinking water.
99
By contrast, knockouts for DNA methyltransferase 1
100
and Keap1
98
are protective against esophageal tumor formation induced by 4-NQO. N-nitrosomethylbenzylamine (NMBA) is another NOC that induces dysplastic changes in the esophageal epithelia of wild-type mice.
101
Interestingly, the route of administration of NMBA influences its mode of carcinogenicity in mice. For example, oral gavage of NMBA was reported to induce esophageal tumors, but only forestomach tumors are observed when NMBA is given intraperitoneally.102–104 Conversely, mice that were given NMBA in their drinking water developed tumors in both the esophagus and the forestomach.
103
Treating
Esophageal adenocarcinoma
Components of gastroduodenal reflux, particularly unconjugated bile acids such as deoxycholate (DCA) induce DNA damage and may contribute to the development of BE.105,106 Given that
BE lesions have been observed in other chemically induced mouse models but never at the frequency reported with the
Other mouse models used to study metastasis
As other alternatives to study metastases, researchers have injected EC cells into the tail vein of nude mice.116,117 Thirty-five to 50 days postinjection, visible lung metastases were observed in control mice injected with the ESCC cell line KYSE-50.116,117 Another group developed a peritoneal metastatic mouse model and examined the effect of combinational therapies with anticancer drugs and triterpenes on tumor metastasis. The peritoneal metastatic mouse model consists of injecting ESCC cells into the peritoneum of immunodeficient mice, which causes tumor colonization of cancer cells in the peritoneal cavities and produces bloody ascites. 118
Conclusion
Over the past 10–15 years, different mouse models of EAC and ESCC have emerged. Although the perfect mouse model of ESCC or EAC does not exist and although the number of models is still very limited, these models have provided valuable insight into esophageal tumor biology. Despite their limitations and their challenges, the value of the mouse models must be recognized because a deeper understanding of this disease is needed to improve patient outcomes. Many preclinical animal models fail to accurately predict the clinical efficacy of anticancer agents. To maximize the potential of mouse models of EC, the combined use of multiple models (human samples, more than one type of mouse model) is highly recommended. This would ensure that the findings obtained with mouse models of EC more closely resemble the human disease. Moreover, further analyses of the currently available models are required to better understand the molecular mechanisms underlying the pathogenesis of EAC and ESCC.
Given that genomic analyses of ESCC and EAC have identified a mutational signature for both types of EC, the generation of future GEMMs should focus on evaluating the consequences of the expression of the mutated genes in EC. In addition, developing future GEMMs of EC should focus on having a faster progression to the development of invasive cancer phenotypes. Another issue for the EC field lies in obtaining a better understanding of the molecular and biological processes involved in the early stages of EC pathogenesis and on metastases, two areas that are currently underinvestigated. Therefore, future mouse models of EC should focus on addressing these matters. Furthermore, it would be beneficial for EC research to generate more transgenic mice expressing new esophageal cell type-specific promoters, including esophageal stem cells. It would also be beneficial for the field to generate mice with inducible expression of the
Author Contributions
Wrote the first draft of the manuscript: MPT. Contributed to the writing of the manuscript: MPT. Jointly developed the structure and arguments for the paper: MPT. Made critical revisions and approved final version: MPT. Author reviewed and approved of the final manuscript.
Footnotes
Acknowledgment
Nature Publishing Group Language Editing participated in English editing of the manuscript.