
Abstract
Caveolae are specialized cell membrane invaginations known to regulate several cancer cell functions and oncogenic signaling pathways. Among other caveolar proteins, they are characterized by the presence of proteins of the cavin family. In this study, we assessed the impact of cavin-1, cavin-2, and cavin-3 on cell migration in a human HT-1080 fibrosarcoma model. We found that all cavin-1, -2 and -3 transcripts were expressed and that treatment with phorbol 12-myristate 13-acetate (PMA), which is known to prime cell migration and proliferation, specifically upregulated cavin-3 gene and protein expression. PMA also triggered matrix metalloproteinase (MMP)-9 secretion, but reduced the global cell migration index. Overexpression of recombinant forms of the three cavins demonstrated that only cavin-3 was able to reduce basal cell migration, and this anti-migratory effect was potentiated by PMA. Interestingly, cavin-3 overexpression inhibited PMA-induced MMP-9, while cavin-3 gene silencing led to an increase in MMP-9 gene expression and secretion. Furthermore, recombinant cavin-3 significantly prevented PMA-mediated dephosphorylation of AKT, a crucial regulator in MMP-9 transcription. In conclusion, our results demonstrate that cellular cavin-3 expression may repress MMP-9 transcriptional regulation in part through AKT. We suggest that the balance in cavin-3-to-MMP-9 expression regulates the extent of extracellular matrix degradation, confirming the tumor-suppressive role of cavin-3 in controlling the invasive potential of human fibrosarcoma cells.
Introduction
Caveolae are specialized cholesterol- and sphingolipid-enriched plasma membrane invaginations, which, in cancer cells, regulate oncogenic functions through multiple signaling cascades. 1 Among these functions, roles of caveolae in mechanosensing, vesicular trafficking, and shuttling of molecules through the cell have been described.2–5 Caveolae have also been ascribed functions in many physiological events, such as wound healing, embryonic development and tissue regeneration,6,7 and in pathological settings such as cancer progression and metastasis. 8 Cancer cell invasion, which involves several cellular processes including disruption of cell adhesion, degradation of the extracellular matrix (ECM) and regulation of cell invasion of blood and lymph vessels, 9 has also been shown, in part, to require caveolae.
Caveolae control cancer cell motility by coordinating several oncogenic signaling intermediates, including eNOS, VEGF receptor 2, EGF receptor, PDGF receptor, several regulators of Ca2+ signaling such as a Ca2+ ATPase, and members of the Rho family of small GTPases.10,11 Expression of caveolar proteins is consequently well recognized as involved in cancer cell invasion, and these proteins are believed to act as either positive or negative regulators of ECM degradation and cancer aggressiveness.8,12 Similarly, uncontrolled proteolysis of ECM components involves a family of zinc-dependant endopeptidase or matrix metalloproteinases (MMPs), amongst which MMP-9 plays a major role in the disruption of the blood-brain barrier, an initial event leading to multiple sclerosis lesion formation, 13 leukemic cell infiltration into the central nervous system,14,15 and sepsis development. 16 Disruption of caveolin-1 has been shown to downregulate MMP-9 expression, 17 while enhanced formation of caveolae has been correlated with increased MMP-9 activity. 18
More recently, it has been suggested that coat proteins called cavins not only work together with caveolins to regulate the formation of caveolae, but also have the potential to transmit signals dynamically from caveolae to various cellular destinations. 19 Of the four cavin proteins, PTRF/cavin-1 has been shown to mediate the reduction of MMP-9 production, independent of caveolar formation, and has been reported to decrease metastatic PC3 prostate cancer cell migration.20,21 Altogether, these observations support the notion that control of MMP-9 expression/secretion occurs through caveolar components or through caveolae-mediated signaling. MMP-9 transcriptional regulation is already, in fact, considered to be triggered by cytokines, such as tumor necrosis factor (TNF)-α, or inducers such as phorbol-12-myristate-13-acetate (PMA), through transcriptional factors including SP1, AP-1 and NF-κB. 22
Several studies have pointed out dual roles for the other cavin proteins. 23 Cavin-2 also known as SDPR, for serum deprivation protein response, and cavin-3 SRBC, as it is an Sdr-related gene product that binds to c-kinase. Cavin-4 MURC, for muscle restricted coiled-coiled protein. Cavin-2 shares 20% homology with cavin-1, and a decrease in cavin-2 expression leads to a decrease in caveolar formation and numbers. It is also localized at the plasma membrane, along with caveolin-1 and cavin-1. 24 Cavin-3 was first identified as a substrate of PKC delta and is located in a tumor suppressor region at chromosome 11p15.5, which is lost in breast, prostate and pancreatic cancer cells. 25 Cavin-4 expression is restricted to smooth muscle, such as skeletal and cardiac myocytes. 26
In the current study, the roles of cavin proteins in the aggressiveness of sarcomas, particularly in human fibrosarcoma cells, have been examined. We first performed a comparative study between the expression of cavins-1, -2 and -3 and assessed their effects on the migration of PMA-activated fibrosarcoma cells. We then focused on the role of the cavin-3-to-MMP-9 signaling axis as a regulator of fibrosarcoma cell migration.
Materials and Methods
Cell Culture, cDNA and siRNA Transfection
The human HT-1080 fibrosarcoma cell line was purchased from American Type Culture Collection and maintained in Eagle's Minimum Essential Medium (EMEM) containing 10% (v/v) fetal calf serum (FCS) (HyClone Laboratories), 2 mM glutamine, 100 units/mL penicillin and 100 μg/mL streptomycin, and were cultured at 37°C under a humidified atmosphere containing 5% CO2. C-terminal turboGFP-tagged full length cDNAs encoding human cavin-1, cavin-2, and cavin-3 were purchased from OriGene Technologies. HT-1080 cells were transiently transfected with expression vectors using the Xtreme Gene transfection reagent (Roche Diagnostics) according to the manufacturer's instructions. Transfection efficiency was confirmed either by confocal fluorescence microscopy or by immunoblotting. The pEGFP cDNA plasmid was used as a control. Gene silencing of cavin-3 was performed through cell transfection of 20 nM cavin-3 specific siRNA (Qiagen, SI02622473), or with a scrambled siRNA sequence (Qiagen, AllStar Negative Control siRNA, SI03650318) as a control using lipofectamine 2000 reagent (Invitrogen) following the manufacturer's instructions. Twenty-four hours post-transfection, cells were treated with 100 nM PMA or vehicle in a serum-free medium for 18 hours.
Cell Migration Assay Using the xCELLigence Biosensor System
Cell migration assay experiments were carried out using the Real-Time Cell Analyzer (RTCA) Dual-Plate (DP) Instrument of the xCELLigence system (Roche Diagnostics). Twenty- four hours post-transfection, mock-, cDNA-, or siRNA-transfected HT-1080 cells were trypsinized and 20,000 cells/well were seeded onto CIM-Plates 16 (Roche Diagnostics). These plates are similar to conventional Transwells (8 μm pore size), with gold electrode arrays on the bottom side of the membrane to provide a real-time measurement of cell migration. Prior to cell seeding, the underside of the wells from the upper chamber was coated with 25 μL of 0.15% gelatin in PBS and incubated for 1 hour at 37°C. PMA was dissolved in serum-free culture medium and added to the upper wells, while the lower chamber was filled with 10% FCS medium to act as a chemoattractant. Cell migration was monitored for 8 hours. The impedance values were measured by the RTCA DP Instrument software and were expressed in arbitrary units as the Relative Cell Migration Index. Each experiment was performed in quadruplicate.
Immunoblotting Procedures
Following treatments or transfection, cells were washed with PBS and lysed with lysis buffer (50 mM Tris-HCl, pH 7.4, 120 mM NaCl, 5 mM EDTA, 0.5% Nonidet P-40, 0.1% Triton) in the presence of phosphatase and protease inhibitors on ice for 30 minutes. Cell debris was pelleted by centrifugation for 10 min at high speed. Protein concentration was quantified using a micro bicinchoninic acid protein assay kit (Thermo Fisher Scientific Inc). Proteins from control and treated cells were separated by Sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis (PAGE). After electrophoresis, proteins were electrotransferred to polyvinylidene difluoride membranes, which were then blocked overnight at 4°C with 5% non-fat dry milk in Tris-buffered saline (150 mM NaCl, 20 mM Tris–HCl, pH 7.5) containing 0.3% Tween-20 (TBST). Membranes were further washed in TBST and incubated with primary antibodies directed against either turboGFP (1/10,000), AKT, phosphorylated AKT (1/1,000), ERK, phosphorylated ERK (1/2,000), or anti-cavin-3 (1/1,500). Washing was then performed in TBST, followed by a 1 hour incubation with horseradish peroxidase-conjugated anti-rabbit IgG (1/10,000) or anti-mouse IgG (1/5,000) in TBST containing 5% non-fat dry milk. Immunore-active material was visualized by Western Lightning Enhanced Chemiluminescence Pro (Perkin Elmer).
Gelatin Zymography
To assess the levels of MMP-2 and MMP-9 activity, gelatin zymography was used as described previously. 27 Briefly, an aliquot (20 μL) of the culture medium was subjected to SDS-PAGE in a gel containing 0.1 mg/mL gelatin. The gels were then incubated in 2.5% Triton X-100 and rinsed in nanopure distilled H2O. Gels were further incubated at 37°C for 20 hrs in 20 mM NaCl, 5 mM CaCl2, 0.02% Brij-35, 50 mM Tris-HCl buffer, pH 7.6, then stained with 0.1% Coomassie Brilliant blue R-250 and destained in 10% acetic acid, 30% methanol in H2O. Gelatinolytic activity was detected as unstained bands on a blue background. All experiments were carried out with cells that had been serum-deprived by overnight incubation.
Total RNA isolation, cDNA Synthesis and Real-Time Quantitative RT-PCR
Total RNA was extracted from cell monolayers using TriZol reagent (Life Technologies). For cDNA synthesis, 2 μg of total RNA were reverse-transcribed using a high capacity cDNA reverse transcription kit (Applied Biosystems). Gene expression was quantified by real-time quantitative PCR using SsoFast EvaGreen Supermix (Bio-Rad). DNA amplification was carried out using a CFX connect Real-Time System (Bio-Rad) and product detection was performed by measuring binding of the fluorescent dye EvaGreen to double-stranded DNA. The QuantiTect primer sets were provided by QIA-GEN: MMP-9 (Hs_MMP9_1_SG, QT00040040), cavin-1 (Hs_PTRF_3_SG, QT02453507), cavin-2 (Hs_SDPR_1_SG, QT00201453), cavin-3 (Hs_PRKCDBP_1_SG, QT00218316), GAPDH (Hs_GAPDH_2_SG QT01192646), β-actin (Hs_Actb_2_SG QT01680476) and PPIA (Hs_PPIA_4_SG QT01866137). The relative quantities of target gene mRNA compared with two internal controls, chosen from GAPDH, β-actin or PPIA RNA, were measured by following a δCT method employing an amplification plot (fluorescence signal vs. cycle number). The difference (δCT) between the mean values in the triplicate samples of target gene and those of GAPDH and β-actin mRNAs were calculated by CFX manager Software version 2.1 (Bio-Rad) and the relative quantified value (RQV) was expressed as 2–δCT.
Statistical Data Analysis
Data are representative of three or more independent experiments. Results are expressed as the mean ± SEM and statistically analyzed using the GraphPad Prism 5 software with two-way ANOVA or one-way ANOVA analysis for three groups or more. Bonferroni post-test with values of p ≤ 0.05 (*) have been considered significant.
Results
PMA Triggers MMP-9 and Cavin-3 Expression, but Reduces the HT-1080 cell Migration Index
Phorbol-12-myristate-13-acetate (PMA) is routinely used to activate several signaling pathways involved in the regulation of cell migration. These include phosphorylation of extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and P38 mitogen-activated protein kinase. 28 However, while it is accepted that PMA-treated cells are consequently in an “activated state”, the overall effect on the cells' migration index has rarely been evaluated. We therefore first investigated the effect of PMA on human HT-1080 fibrosarcoma cell migration. Live cells were seeded and cell migration assessed in real-time as described in the Methods section. We observed that PMA dose-dependently reduced the relative cell migration index (Fig. 1A) while it concomitantly induced MMP-9 and cavin-3 gene expression levels, but not those of cavin-1 or cavin-2 (Fig. 1B). The MMP-9 increases were further reflected by increases at the functional level as measured by zymography, which shows increased hydrolytic activity of MMP-9 secreted in the cell conditioned media, while that of MMP-2 remained relatively unaltered (Fig. 1C). The protein expression level of cavin-3 was increased in cell lysates as determined by immunoblotting upon PMA treatment (Fig. 1D). Interestingly, we observed that 10 nM PMA maximally induced MMP-9 gene and protein expression. While such expression reached a plateau, that of cavin-3 was still able to be induced at higher PMA concentrations. This can, in part, be explained by the differential involvement of transcriptional factors in the regulation of each of MMP-9 and cavin-3 genes. Moreover, the fact that increasing cavin-3 expression correlated with a decreased-to-plateau effect on MMP-9 supports that cavin-3 exerts repressive regulation on MMP-9. Altogether, although increased MMP-9 secretion is classically associated with increased cell migration, our data suggeste that the relative migration index of PMA-activated cells is actually reduced, as has been also reported in anaplastic human thyroid cancer cells, 29 and is correlated with increased cavin-3 expression. The existence of alternate compensatory mechanisms, which possibly involve cavin-3, was next explored.
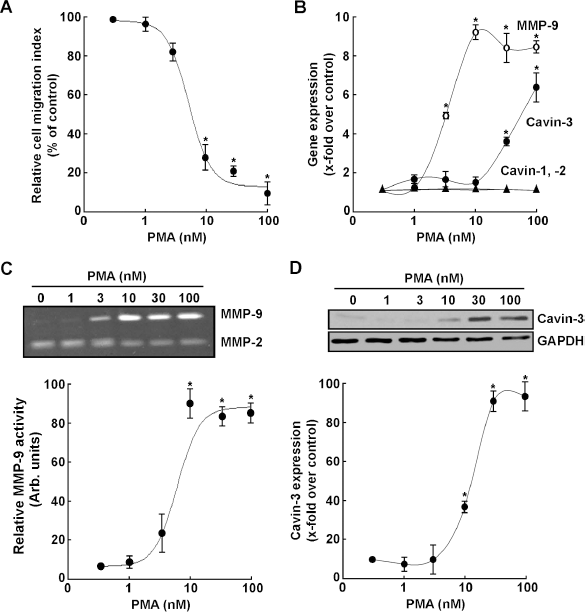
PMA treatment triggers MMP-9 secretion and cavin-3 expression, but reduces the HT-1080 cell migration index. (A) Human HT-1080 fibrosarcoma cells were treated with varying concentrations of PMA in serum-free medium for 18 hours. Cells were then collected and analyzed for their migration capacity as described in the Methods section. (B) Total RNA was extracted from the remaining of the cells and qRT-PCR was performed to assess expression of cavin-1, cavin-2, cavin-3 and MMP-9 transcripts. Values were normalized over the housekeeping genes GAPDH and PPly and are the mean ± S.E.M of triplicate values from one out of three representative experiments. (C) Conditioned media was harvested to assess the extent of MMP-2 and MMP-9 gelatinolytic activity by zymography. (D) PMA-induced changes in endogenous cavin-3 protein expression were analyzed by Western blotting.
Overexpression of Recombinant Cavin-3 Specifically Decreases Cell Migration and Abrogates PMA-Induced MMP-9 Expression
In order to directly assess the impact of cavin-3 on cell migration, cDNA plasmids encoding green fluorescent protein (GFP)-tagged recombinant cavin-1, cavin-2, or cavin-3 were transiently transfected in HT-1080 cells. While the fluorescent recombinant cavin proteins were all expressed, as confirmed by confocal fluorescent microscopy (Fig. 2A), only cavin-3 significantly reduced the HT-1080 cell migration index (Fig. 2B). The impact of recombinant cavins was also assessed on MMP-9 transcription, and we observed that cavin-3 was the only recombinant cavin protein tested that inhibited both endogenous (Fig. 2C, white bars) and PMA- induced MMP-9 gene expression (Fig. 2C, black bars). To test whether a balance exists between cavin-3 expression and MMP-9 expression, we transfected increasing amounts of cavin-3 cDNA in HT-1080 cells and then stimulated them with PMA. We found that expression of recombinant GFP-tagged cavin-3, discriminated from the endogenous form via detection with the anti-turbo-GFP antibody, increased in cell lysates in a dose-dependent manner in both untreated and PMA-treated cells (Fig. 3A), whereas MMP-9 secretion was only expressed in PMA-treated cells (Fig. 3B) and expression inhibited with increasing quantity of cavin-3 cDNA transfected. Overall, these observations confirm that a cavin-3/MMP-9 expression balance may exert a crucial regulation of cell migration, and that cavin-3 could in fact exert tumour suppressive activity in human fibrosarcoma cells, in part, by downregulating cell migration in PMA-activated cells through decreased MMP-9 expression.
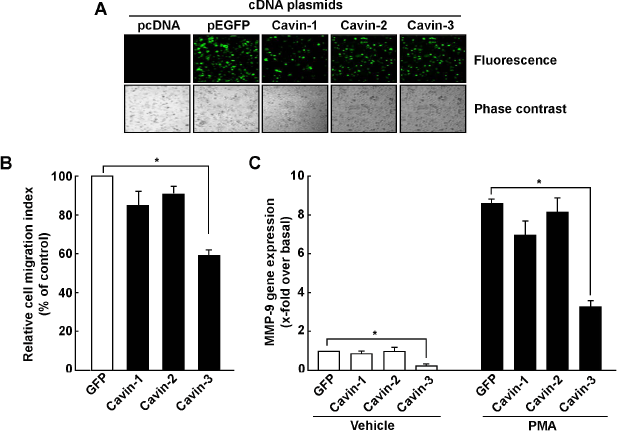
Overexpression of recombinant cavin-3 specifically decreases cell migration and abrogates PMA-induced MMP-9 expression. (A) Human HT-1080 fibrosarcoma cells were transiently transfected with 2 μg of cDNA plasmids and fluorescent confocal microscopy was used to visualize the encoded recombinant GFP-tagged cavin-1, cavin-2 or cavin-3 proteins 24–48 hours post transfection. pEGFP cDNA plasmid was used as control. (B) Cells were then harvested and cell migration assayed as described in the Methods section. (C) HT-1080 cells were transiently transfected with plasmids as described above. Twenty-four hours post transfection, cells were treated with vehicle (white bars) or 100 nM PMA (Black bars) in a serum-free medium for 18 hours. Total RNA was extracted and qRT-PCR was performed to measure the levels of MMP-9 transcript. Values of PCR are normalized over the expression of housekeeping genes GAPDH and PP/γ and are the mean ± S.E.M of triplicate values from one representative experiment.
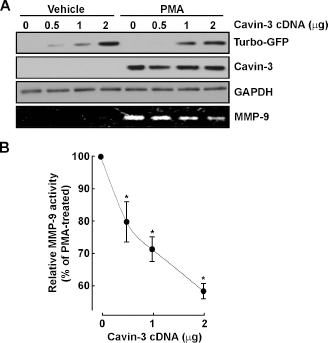
Recombinant cavin-3 expression abrogates PMA-induced-MMP-9 secretion. Human HT-1080 fibrosarcoma cells were transiently transfected with various amounts of a GFP-cavin-3 cDNA plasmid. twenty-four hours post transfection, cells were treated with vehicle or 100 nM PMA in a serum-free medium for 18 hours. (A) Cell lysates were used to analyze endogenous cavin-3, GFP-tagged recombinant cavin-3, and GAPDH expression by western blotting, and conditioned media was used to evaluate MMP-9 secretion by gelatin zymography (lower panel). (B) scanning densitometry was used to quantify the extent of gelatinolytic activity of MMP-9.
Loss of Cavin-3 Function Restores PMA-Decreased Cell Migration and Potentiates PMA-Induced MMP-9 Expression
In order to further examine the possible role of cavin-3 as a tumor suppressor via the balance between its expression and that of MMP-9 in PMA-treated cells, we used gene silencing strategies to repress cavin-3 gene expression by transiently transfecting HT-1080 cells with a siRNA against cavin-3. Gene silencing efficiency was demonstrated in both untreated and PMA-treated cells to range between 74–91% (Fig. 4A). When the relative cell migration indices were measured, the decrease in migration observed in PMA-treated cells was completely prevented when cavin-3 expression was silenced (Fig. 4B). Loss of Cavin-1 or Cavin-2 did neither alter basal cell migration nor did it reverse PMA-mediated decrease in cell migration (data not shown). Interestingly, loss of cavin-3 expression induced endogenous MMP-9 secretion in the conditioned media, whereas it significantly enhanced PMA's ability to trigger MMP-9 secretion (Fig. 4C). Such MMP-9 transcriptional regulation was also observed in the basal condition and in the PMA-treated cells, where silencing of the cavin-3 gene led to a further increase in MMP-9 transcription by PMA (Fig. 4D). Altogether, our data highlight an unreported contribution of cavin-3 in regulating cell migration and MMP-9 expression in PMA-activated cells. The identity of the specific cell signaling pathways involved in this process was next investigated.
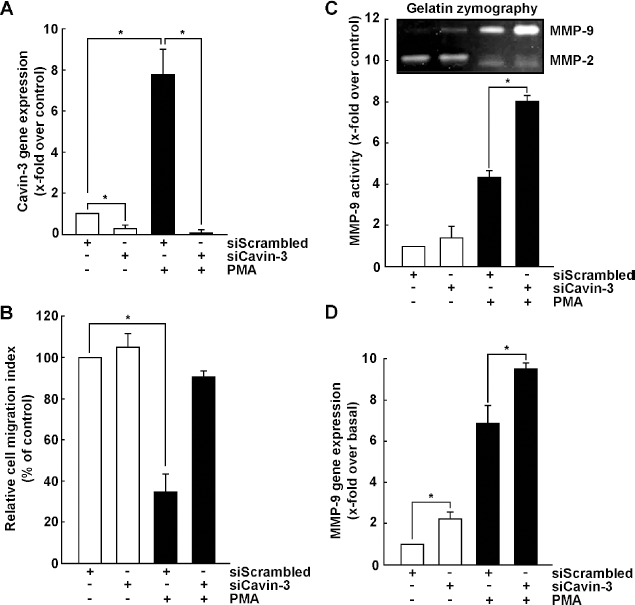
Loss of cavin-3 function restores PMA-decreased cell migration and potentiates PMA-induced MMP-9 expression. Human HT-1080 fibrosarcoma cells were transiently transfected with 20 nM siRNA against cavin-3 (siCavin-3) or a random sequence (siScrambled) as described in the Methods section. Cells were then treated with vehicle or with 100 nM PMA for 18 hours. Total RNA was extracted and qRT-PCR was performed to assess (A) cavin-3 and (D) MMP-9 transcript levels. Values were normalized over the expression of the housekeeping genes GAPDH and PP/γ and are the mean ± S.E.M of triplicate values from one representative experiment. (B) Cells were treated as described above, collected and analyzed for cell migration as described in the Methods section. (C) Conditioned media was also collected to assess the level of MMP-2 and MMP-9 gelatinolytic activity by zymography.
Cavin-3 Overexpression Does Not Alter PMA-Induced ERK Phosphorylation, but Potentiates PMA-Mediated AKT Dephosphorylation
It was recently suggested that cavin-3 dictates the balance between ERK and AKT signaling. 30 However, it is currently unknown whether either of these two signaling intermediates is also affected by cavin-3 in PMA-activated cells. Recombinant cavin-3 was therefore transiently overexpressed in HT-1080 cells, which were subsequently treated with PMA for up to 30 minutes in order to investigate the phosphorylation status of ERK1/2 and of AKT (Fig. 5A). Transfection efficiency and specific expression of the recombinant cavin-3 protein were confirmed as detected by the anti-turbo-GFP antibody, and did not vary upon PMA treatment (Fig. 5A). While the extent of ERK phosphorylation was unaltered by PMA (Fig. 5B), overexpression of recombinant cavin-3 significantly potentiated AKT dephosphorylation by PMA (Fig. 5C). Our results suggest that part of cavin-3's anti-migratory action may be attributed to its ability to alter the AKT signaling pathway, which, in turn, can also affect MMP-9 transcriptional regulation.
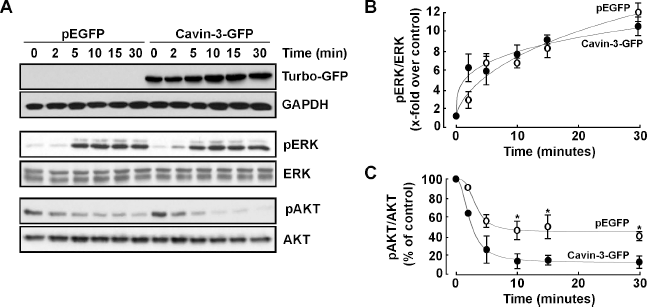
Cavin-3 overexpression does not alter PMA-induced ERK phosphorylation, but potentiates PMA-mediated AKT dephosphorylation. (A) Human HT-1080 fibrosarcoma cells were transfected with 2 μg of either pEGFP or cavin-3 cDNA plasmids. Twenty-four hours post transfection, cells were then treated with 100 nM PMA in serum-free medium for the indicated time. Cells were then lysed and the phosphorylation status of ERK1/2 and AKT assessed by Western blotting. Scanning densitometry was used to quantify the extent of phosphorylation of (B) ERK and (C) AKT in pEGFP- (open circles) and cavin-3-GFP- (closed circles) transfected cells.
Discussion
Assessment of in vitro cell migration alterations classically uses strategies to prime the cells, such as in response to serum-derived growth factors or to specific cytokines.31,32 Given the lack of in vitro microenvironment components and extracellular growth factor-mediated cues, treatments with phorbol esters such as PMA are therefore generally used to either trigger secretion of ECM hydrolytic enzymes such as MMPs or to initiate signaling pathways that affect cell migration.33,34 While several of these signaling pathways are well documented, the effective balance between positive and negative regulators of cell migration, within a defined cell model, in fact still remains poorly understood.
In this study, we used a human fibrosarcoma cell line that responded to PMA in part through increased secretion of MMP-9, 33 the secretion of which is classicaly correlated with pro-inflammatory,35,36 pro-angiogenic and metastatic processes. 37 Our study demonstrates that, although both MMP-9 expression and cell migration can effectively be triggered in vitro upon a defined PMA treatment, the overall effect that is actually measured is, in fact, a diminished cell migration index, in part consequent to the increased expression of the tumor suppressor cavin-3. In fact, our study suggests that PMA should rather be considered, in specific cell models, for its anti-migratory effects as it impaired HT-1080 fibrosarcoma cell growth and proliferation in agreement with previous reports.38–40
Our study further provides new insight into the tumor suppressive functions of cavin-3. We show that endogenous cavin-3 is undetectable at the protein level in the HT-1080 fibrosarcoma cell model we used, in agreement with the fact that many other cancer cell lines effectively also lack cavin-3,26,41,42 and this precluded any cavin-3 gene silencing strategies to be used in the non-activated cellular model. Cavin-3 expression was in fact shown to require the nuclear factor-kappa B (NF-κB) pathway following TNF-α-activation.42,43 Interestingly, PMA is also a well-established NF-κB activator15,27,35,44 and it is believed to relay its effects, to some extent, through similar signaling. More importantly, our study also demonstrates for the first time that overexpression of cavin-3 itself reduces basal as well as PMA-induced cell migration. Such regulation is believed to occur at the MMP-9 transcriptional level given that overexpression of recombinant cavin-3 reduced endogenous as well as PMA-induced MMP-9 expression. While cavin-3 overexpression did not affect PMA-induced ERK phosphorylation, we found that recombinant cavin-3 further and significantly reduced PMA-mediated AKT phosphorylation. Our results are in good agreement with a recent postulate that links ERK and AKT pathways to cell migration,45,46 and with evidence demonstrating that introduction of cavin-3 into a lung carcinoma cell line reduced AKT signalling.30,47 Blockage of the PI3K/AKT and MAPK signalling pathway was also found to decrease MMP-9 protein expression in an ovarian cancer cell model. 48 It is therefore tempting to speculate that cell migration is decreased as a consequence of cavin-3-mediated signaling, which prevents phosphorylated-AKT transcriptional regulation of MMP-9 gene expression. Prohibition of nuclear phosphorylated-AKT was recently reported also to inhibit epidermal growth factor (EGF)-induced MMP-9 transcription in glioblastoma cells. 49 Loss of Cavin-3 is therefore believed to alter the cell's ability to assemble optimal signaling modules with cell surface receptors. 30 Finally, it was previously demonstrated that cavin-1, another member of the cavin family, also decreased MMP-9, in prostate cancer. 20
As is the case for cavin-3, MMP-9 can also be activated by TNF-α, PMA, and EGF.36,44,50,51 Here, we show that PMA effectively induces MMP-9 transcription and secretion in HT-1080 fibrosarcoma cells, but that this induction in MMP-9 is actually the result of a change in the balance between PMA-induced MMP-9 and cavin-3 signaling (summarized in Fig. 6). This signaling balance was further strengthened by siRNA-mediated gene silencing of cavin-3, where silenced expression was demonstrated to relieve cavin-3's negative regulation of MMP-9, and to allow higher expression. Increases in MMP-9 expression by PMA can, in fact, be induced through activation of several signaling cascades involving mitogen-activated protein kinase (MAPK) and phosphoinositide-3 kinase (PI3K)/AKT. 52 Several transcription factors are involved in the transcriptional regulation of MMP-9 induced by PMA, including AP-1 and NF-κB through ERK, JNK and PI3K/AKT signaling pathways; the MMP-9 promoter bears several of the DNA consensus binding sequences corresponding to these transcriptional factors.53–56 Conceptual limitations of our current study must be forthrightly acknowldged as the results presented and conclusions raised may not necessarily be generalized to other cellular models. However, HT-1080 fibrosarcoma cells are considered as a relatively well-established cellular model and validated for studies related to in vivo mice models, 57 methodology development, 58 PMA impact assessment, 59 and in models assessing metastasis and cancer cell motility. 60 In conclusion, our study still demonstrates that cavin-3 expression potentiated a decrease in PMA-mediated AKT dephosphorylation and that cavin-3 loss of function restored increased MMP-9 expression upon PMA treatment. Our data therefore suggest that cavin-3 lessened phosphorylated-AKT transcriptional regulation of MMP-9 expression, and consequently decreased MMP-9-mediated ECM degradation, which in turn reduced cell migration of HT-1080 fibrosarcoma cells. Given that cavin-3 is ubiquitously expressed, it still remains however unknown whether differential cell-type functions are involved. 61
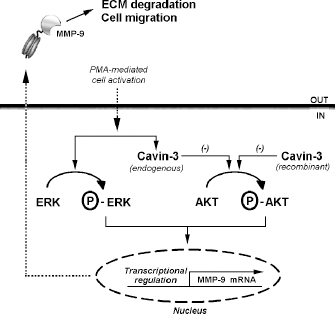
Scheme summarizing cavin-3-dependent regulation of PMA-activated HT-1080 fibrosarcoma cells. PMA-activated human HT-1080 fibrosarcoma cells show increased levels of ERK phosphorylation, endogenous cavin-3, and MMP-9 expression, all of which contribute to characterize the cells' migration index. We demonstrate that at certain doses PMA also inhibits AKT phosphorylation, an effect correlated with PMA-induced endogenous cavin-3 expression. This effect can be exacerbated by overexpression of constitutive recombinant cavin-3. Cavin-3 therefore potentiates PMA-mediated AKT dephosphorylation and prevents endogenous MMP-9 expression. These effects can be reversed upon cavin-3 gene silencing (Fig. 4). We conclude that a crucial role for the cavin-3/MMP-9 signaling axis may contribute to the regulation of PMA-activated human fibrosarcoma cell neoplastic phenotype.
Abbreviations
extracellular matrix;
extracellular signal-regulated kinase;
matrix metalloproteinase;
nuclear factor-kappa B;
phorbol-12-myristate-13-acetate;
tumour necrosis factor.
Author Contributions
Conceived and designed the experiments: CT, BoA. Analyzed the data: CT, CC, BaA. Wrote the first draft of the manuscript: CT, BoA. Contributed to the writing of the manuscript: CT, BaA. Agree with manuscript results and conclusions: CT, CC, BaA. All authors reviewed and approved the final manuscript.