
Abstract
A variety of post-translational protein modifications (PTMs) are known to be altered as a result of cancer development. Thus, these PTMs are potentially useful biomarkers for breast cancer. Mass spectrometry, antibody microarrays and immunohistochemistry techniques have shown promise for identifying changes in PTMs. In this review, we summarize the current literature on PTMs identified in the plasma and tumor tissue of breast-cancer patients or in breast cell lines. We also discuss some of the analytical techniques currently being used to evaluate PTMs.
Introduction
Viable cells maintain membrane integrity, cytoskeleton morphology and proliferation status based on changes in protein structure and function. The complexity of regulation of so many different biomolecules goes beyond the “central dogma” of biochemistry, which implies that one gene encodes for one protein. This complexity of regulation not only results from variable mRNA splicing and DNA transcription, such that one gene can produce many mRNA and protein sequences, but also because one protein sequence can have multiple functions as a result of covalent modifications after synthesis. These post-translational modifications (PTMs) include phosphorylation, methylation, glycosylation, acylation, oxidation and ubiquitinylation. During cancer progression, many PTMs contribute to abnormal cellular proliferation, adhesion characteristics and morphology.1–4 In breast cancer, recent studies suggest that PTM profiles can be used as “biochemical footprints” for tracking and verifying the function and activity of key cellular signaling pathways.5–7 This conclusion suggests that, for early detection, PTMs may be useful biomarkers.
Breast cancer is the second most common type of cancer (after lung cancer), and the fifth most common cause of cancer death. According to the American Cancer Society, in 2008, an estimated 182,460 new cases of invasive breast cancer were diagnosed among US women. Approximately 40,480 of these women are expected to die from this disease (http://www.cancer.org/downloads/STT/2008CAFFfinalsecured.pdf). Like many other cancers, breast cancer is the result of multiple environmental and hereditary factors. Although risk factors such as lesions to DNA, failure of immune surveillance, abnormal growth factor signaling, and inherited or somatic genetic defects (e.g. in p53, BRCA1, BRCA2 genes) are associated with breast cancer development, the cause of any individual breast cancer case is typically unknown. As many studies have suggested, changes in gene expression levels for breast cancer may not fully reflect the true state of cancer progression or development.5,8,9 This conclusion suggests that many of the differences between normal and cancer tissue may be caused by PTMs.1,3,5,6,8–10
This review mainly focuses on the most recent publications on PTMs (especially oxidation and glycosylation) discovered in blood or tissue from breast cancer patients or from breast cancer cell lines. For more general reviews of PTMs, see prior reviews.1–4,7,11
Enzymatic PTMs
Covalent modification of one or more amino acids of a given protein can dramatically alter the biological function of that protein. The likelihood that a particular reactive protein residue will undergo a modification reaction is influenced by the spatial orientation of that amino acid residue(s) in the protein, and is influenced by the adjacent amino acids, which can alter reactivity of the susceptible amino acid by influencing its electrophilic nature. Specific enzymes commonly catalyze these reactions. For example phosphorylation (phosphokinase), methylation (methylase), acetylation (acetyltransferase), and glycosylation (glycosyltransferases) are PTMs that are mediated by the indicated enzyme.1–4,7,11 Many PTMs also result from spontaneous reaction of susceptible residues with certain reactive chemical agents. For example, glycation (commonly called advanced glycation end product, or AGE) is the result of an activated sugar molecule, such as fructose or glucose, bonding to a protein without direct enzymatic involvement. 12 For other PTMs, although enzymes may play an important role in producing the reactive molecule that results in the protein modification, the covalent modification occurs spontaneously without enzymatic activity. For the purposes of this review, we define these PTMs as non-enzymatic if an enzyme is not required for the actual protein modification. For example, peroxynitrite directly reacts with proteins to form nitrotyrosine.13–15 Although peroxynitrite formation appears to require enzymatic production of reactive precursors, its binding to proteins is non-enzymatic, and therefore we consider nitrotyrosine to be a non-enzymatic PTM.
Phosphorylation
Phosphorylation is well recognized as a key regulator of enzyme activity. As the extensive research in protein phosphorylation has been carefully reviewed by others,16–18 we only briefly cover this topic here. Abnormal phosphorylation of defined signal transduction pathways can alter the growth properties of breast tumors. With the use of sequence-specific antibodies against phosphorylation sites, analysis of protein phosphorylation profiles allows one to determine the activation status of signaling pathways, which can provide valuable prognostic insights.19–21 Atsriku et al undertook a systematic mapping of PTMs in the human estrogen receptor alpha (ER-a) in the MCF7 breast cancer cell line. They applied HPLC-ESI and MALDI-MS techniques to identify the phosphorylation sites on the estrogen receptors in these cells. 22 Several novel phosphorylated serine residues were identified. The use of both HPLC-ESI and MALDI gave higher sequence coverage than either approach alone. Nine phosphorylated serine residues were identified, three of which were previously unreported. 22
Acetylation
Histone acetyltransferases and histone deacetylases modify histones by adding or removing an acetyl group from the e-amino group of lysines within a conserved lysine motif. Histone acetylation results in changes in chromatin structure in response to specific endocrine signaling in several cancers, including breast cancer. Recent studies found that acetylation of the ER is mediated by histone acetylases.23–25 The acetylation of ER-α alters its function in estrogen-dependent signaling.23,24 The regulation of ER by deacetylation provides a direct link between intracellular metabolism and hormone signaling.25,26 Wang et al
27
showed that the acetylation of ER-α alters its function
Glycosylation
Summary of recent glycosylation PTMs in breast cancer studies.
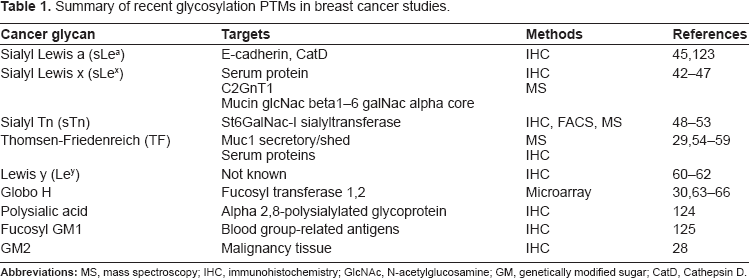
Changes in glycosylation for cancer cells include both reductions and increases in naturally occurring glycans, as well as increases in glycans primarily restricted to embryonic tissues.34,35 One of the most common changes is an increase in the side branching of N-linked glycans. 36 This increased branching is often attributed to increased activity of N-acetylglucosaminyltransferase V (GlcNAc-TV, also known as MGAT5; the enzyme that leads to β1,6GlcNAc branching).37–40 The increased branching creates additional sites for terminal sialic acid residues, which, in combination with up-regulation of sialyltransferases, leads to an increase in global sialylation. 41
In addition to changes in glycan core structures, altered terminal structures are commonly associated with malignant breast cancer.42–47 Glycosyltransferses (e.g. sialyltransferases and fucosyltransferases) involved in adding terminating residues to glycans tend to be over-expressed in breast cancer tissue.29,30,48–66 The increase in activity of these glycosyltransferases, in turn, leads to an increase of certain terminal glycans. Glycan residues commonly found on transformed cells include sialyl Lewis x, sialyl Tn, Globo H, Lewis y and polysialic acid. Many of these glycans are observed in malignant breast tissues (summarized in Table 1).
Non-Enzymatic PTMs
Oxidation
It has been hypothesized that cancer development is a process that is similar to “wounds that never heal”.67–70 Various studies have suggested that inflammation, which increases oxidative stress, is associated with cancer development or metastasis.67,71–74 Both mouse models and human-pathology studies suggest that there is a strong immune response in the early stages of breast cancer that disappears in more advanced disease.72,75 Consistent with this observation, tumor levels of nitrotyrosine (nTyr), which are believed to be indicative of NO and superoxide levels, have been reported to be increased in the early breast cancer, but not in more advanced disease.72,75 The NO and superoxide may be produced by activated macrophages. Therefore, localized oxidative stress associated with the immune response to breast cancer might result in modifications of proteins secreted by the breast cancer cells that could be used to detect early disease. Reactive oxygen species (ROS) also regulate the synthesis and secretion of many receptor ligands (e.g. growth factors and chemokines).76–78 These factors regulate important processes in epithelial cancers, including the ligand-dependent activation of the proliferation (MAPK/Erk) and anti-apoptosis (PI3K/Akt) pathways.79–81 Therefore, proteins modified by ROS may be useful biomarkers that can provide insight into molecular processes occurring in tumors. The oxidative stress associated with the immune response results in protein modifications that may be useful in detecting early breast cancer.
Summary of recent oxidation PTMs in breast cancer studies.
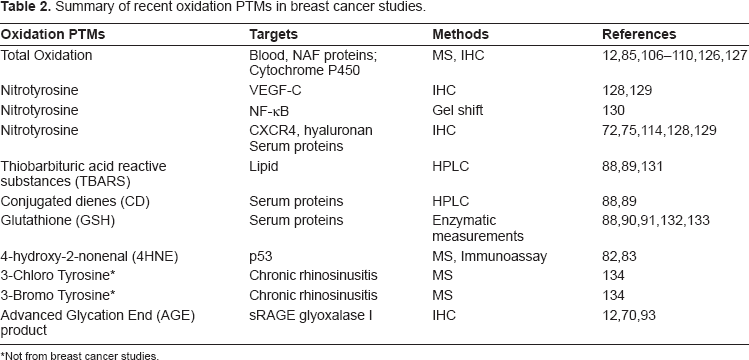
Not from breast cancer studies.
Advanced Glycation End
Oxidative and carbonyl stress may contribute to the progression of cancer; on the other hand, these modifications may have some antiproliferative effects. Tesarova et al 12 reported that serum levels of AGEs, carboxymethyllysine and advanced oxidation protein products (AOPP) in 86 patients with breast cancer and in 14 healthy age-matched control women could be subdivided based on the clinical stage, histological grading, and expression of hormone and Her2 receptors. Breast cancer patients had higher serum concentrations of AGEs even in the early stages of this disease; patients with advanced breast cancer (stages III and IV) had significantly higher AGE levels, not only compared to controls, but also compared to stages I and II breast cancer cases.12,70,93 Serum levels of AOPP were higher in patients having only weakly positive expression of Her2 compared to controls and in patients having the highest Her2 expression. 12 These authors concluded that breast cancer patients had an early increase of AGEs (a marker of the carbonyl stress) followed by further increase of AGEs and elevation of AOPP (a marker of oxidative stress) in more advanced disease. As the clinical significance of these observations is currently uncertain, further studies are needed to validate these results in terms of the usefulness of AGE in the early detection of breast cancer.
Methods for PTM Discovery and Analysis
Mass-Spectrometry-Based Proteomics
Summary of enzymatic PTMs biomarkers in breast cancer research.
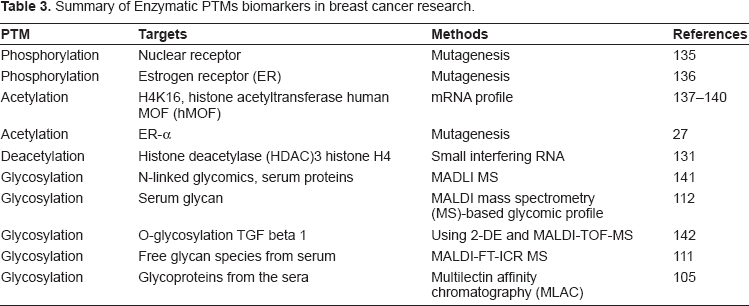
Most PTMs are present at low levels in cells and tissues, and are therefore difficult to detect by MS. For this reason, modification-specific analytical strategies that are designed to improve sensitivity and specificity have been employed to enrich and concentrate a specific class of PTM in complex biological samples. PTM peptide enrichment can employ either affinity 105 or chemical methods.12,106–110 During the MS analysis, multi-stage MS techniques that further fragment suspected PTM peptides 111 can improved confidence in peptide identification. Identification of PTMs commonly requires specialized bioinformatics tools, the validation of results by replicate analyses42–47 and follow-up biological experiments. 112 Such PTM-specific methods can be combined with semi-quantitative techniques, including stable-isotope labeling and peptide-intensity profiling. PTM-targeted methods have also been combined with subcellular fractionation to obtain biological insights about in the roles of specific organelles.113–117
ELISA Microarray
The microarray sandwich ELISA is an exceptionally sensitive analytical technique that can accurately measure individual protein concentrations down to the low or sub-pg/ml range.115,117–120 Adapted from the conventional sandwich ELISA, the ELISA microarray commonly uses complementary pairs of capture and detection antibodies (or, for glycan analysis, lectins) to measure trace antigens in complex biological fluids. The microarray technique is also suited for targeted discovery research because of its ability to simultaneously conduct multiple assays. At the same time, this multiplex analysis requires very little sample (20 or less, of diluted sample per multiplexed analysis, after at least a 5-fold dilution), thereby allowing the screening of many PTMs using very small sample volumes. Even so, there are several challenges for ELISA microarray analysis. One challenge is the need for highly specific antibodies. There is limited commercial availability of good antibodies for many PTMs. Classical strategies of antibody generation by animal immunization may not result in high-quality antibodies for the targeted PTM. The second challenge is the potential for cross reactivity with nonspecific antigens.
Immunohistochemistry
Immunohistochemistry (IHC) has been widely used for evaluating PTMs in breast cancer.113,116,121 To identify PTMs as potential tumor markers, IHC offers a rapid method for comparing PTM levels in cancer tissue and adjacent normal tissue. Altered expression and PTM of several proteins using immunoblot analysis and IHC have been reported by several research groups (Tables 1–3). For example, modification of the beta subunit of prolyl-4-hydroxylase and of annexin A2 in tumor tissues was confirmed by immunoblot and immunohistochemistry. 122 The determination of nitrotyrosine levels by IHC of breast cancer carcinoma tissue has been reported. 75 A drawback of IHC in PTM analysis is the difficulty in quantifying the results.
Conclusion
Plasma-, tissue- or cell-based studies for PTM biomarkers in breast cancer have provided promising data. Several PTMs can only be readily detected in breast cancer tissue but not in normal breast. In particular, glycosylation and oxidative modifications appear to have potential as biomarkers. These results suggest that levels of certain PTMs may be indicative of breast cancer progression or development, although the data on which proteins are actually modified is still very limited. Once this deficit is addressed, we conclude that the post-translational modifications on specific proteins may be useful as biomarkers for breast cancer.
Abbreviations
advanced glycation end products
breast-cancer susceptibility gene
conjugated dienes
ductal carcinoma in situ
enzyme-linked immunosorbent assay
estrogen receptor
4-hydroxynonenal
high-performance liquid chromatography-electrospray ionization-mass spectroscopy
immunohistochemistry
N-acetylglucosamine
N-acetylglucosaminyltransferase V
glutathione
glutathione S-transferase P1
mitogen-activated protein kinase/extracellular signal-regulated kinase
lobular carcinoma in situ
matrix-assisted laser desorption ionization-mass spectroscopy
nitric oxide
3-nitrotyrosine
RAS association family 1 gene
phosphoinositide-3 kinase/protein kinase B
post-translational modifications
Protein Microarray Analysis Tool
reactive oxygen species
sialyl lewis a
sialyl lewis x
sialyl Tn
Lewis y
surface-enhanced laser desorption/ionization
time-of-flight
mass spectroscopy
thiobarbituric acid reactive substances
Thomsen-Friedenreich
Disclosures
The authors report no conflicts of interest.
Footnotes
Acknowledgements
We thank Ms. Julie Wiley for the critical editing of this manuscript. This review was supported by the NIH grants U01 CA117378, R01 EB006177 and U54 ES016015.