
Abstract
Tumor targeting is a booming business: The global therapeutic monoclonal antibody market accounted for more than $78 billion in 2012 and is expanding exponentially. Tumors can be targeted with an extensive arsenal of monoclonal antibodies, ligand proteins, peptides, RNAs, and small molecules. In addition to therapeutic targeting, some of these compounds can also be applied for tumor visualization before or during surgery, after conjugation with radionuclides and/or near-infrared fluorescent dyes. The majority of these tumor-targeting compounds are directed against cell membrane-bound proteins. Various categories of targetable membrane-bound proteins, such as anchoring proteins, receptors, enzymes, and transporter proteins, exist. The functions and biological characteristics of these proteins determine their location and distribution on the cell membrane, making them more, or less, accessible, and therefore, it is important to understand these features. In this review, we evaluate the characteristics of cancer-associated membrane proteins and discuss their overall usability for cancer targeting, especially focusing on imaging applications.
Introduction
Tumor targeting is a relatively novel but rapidly expanding technique applied for cancer treatment as well as visualization. Targeted anticancer therapies consist generally of antibodies or antibody-derived fragments, proteins, peptides, small molecule inhibitors, or DNA/RNA aptamers directing an attached drug to the tumor cell.1,2 Just the global therapeutic monoclonal antibody market alone accounted already for more than $78 billion in 2012, indicating the potential of targeting for research and development. 3 Of this total amount, 75% was spent for arthritis and cancer, with Remicade, Avastin, Rituxan, Humira, and Herceptin respectively being the top five mega sellers.
Tumor targets are in general membrane proteins or, in some cases, their ligands, with enhanced expression on tumor or tumor-associated cells, such as malignant cells, angiogenic endothelial cells, or inflammatory cells. 2 Besides therapy, the targeting component of these drugs is in principle also suited for the development of tumor-visualizing tracers, which could be used for the early diagnosis or localization of tumors before or during surgery. Generally, all membrane proteins that are overexpressed on tumor or tumor-associated cells are potentially suitable for tumor-targeted imaging. Out of the ~7,000 known transmembrane proteins, ~150 are overexpressed on tumor cells or tumor-associated vessels, which makes them potential candidates for therapeutic targeting or imaging. 4 However, there is surprisingly little knowledge about which targets should be used for optimal results per tumor type, or even better, per individual tumor or patient.5,6 To date, most of the imaging probes have been designed to target the vast majority of tumors. The development of personalized probes, customized for specific tumor types, will be inevitable for optimal clinical applications and will require more specific knowledge about tumor targets.
This overview of possible tumor targets is primarily based on the prioritization list of cancer antigens issued by the National Cancer Institute (NCI). 7 In this list consisting of 75 proteins, only 13 members are actually cell membrane-associated proteins. Seven of these proteins are receptors and three are linked to the cell via a glycosylphosphatidyl inositol (GPI) anchor. The other membrane proteins in the list comprise two enzymes, an adhesion molecule, and a glycoprotein with a lubrication/barrier/signaling function (Table 1). To put the various targeting candidates into perspective, the following section provides an overview of each subgroup of membrane proteins. The focus will be on the general characteristics with respect to their function, type of anchorage, and behavior within the cell membrane. For each subgroup, we highlight the proteins from the NCI prioritization list, complete with interesting/important targets from the recent literature. The review concludes with a general section on the optimal characteristics of tumor-imaging targets, followed by a paragraph on future perspectives.
Characteristics of tumor-associated membrane proteins.
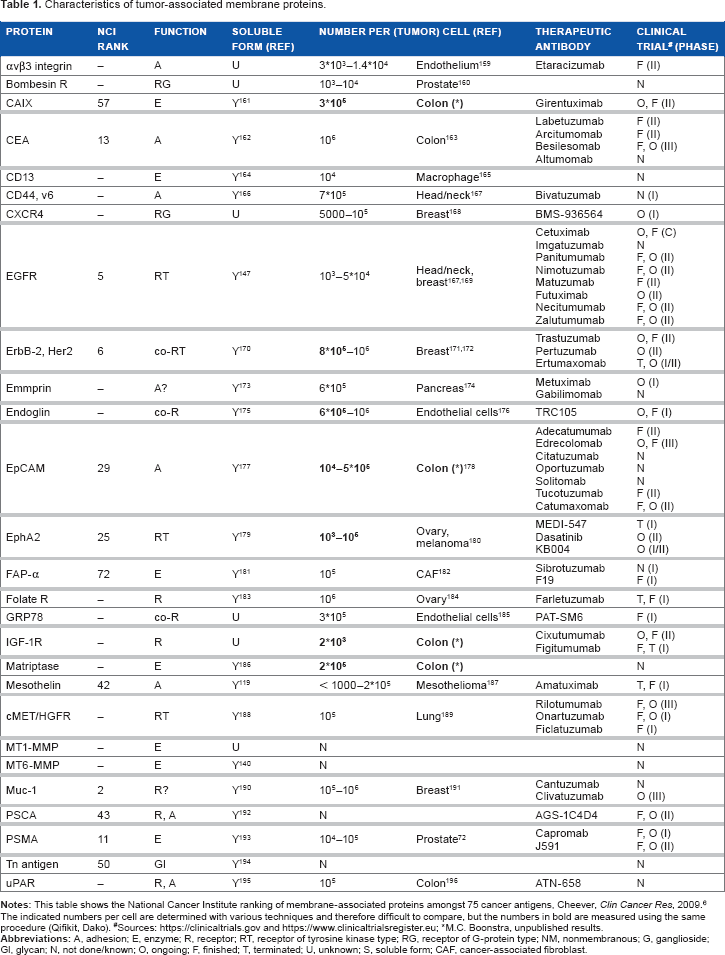
Types and Functions of Membrane Proteins
Based on their topology and structure, membrane proteins are historically categorized as subclass types I, II, III, IV, or V.8,9 Most eukaryotic membrane proteins belong to type I or type III, with respectively one or multiple transmembrane spanning domains. Type II membrane proteins span the membrane similar to type I, but in an opposite orientation, ie, with the amino terminus within the cytoplasm. Type IV and V membrane proteins are anchored to the cell via a chain of lipids or a GPI module (Fig. 1). For this overview, we use a slightly adapted classification, subdividing membrane proteins based on their biological functions into the following: A) receptors, B) cell adhesion or anchoring proteins, C) cell membrane-associated enzymes, D) transporter proteins, and E) GPI proteins 4 (schematic overview is provided in Fig. 2). Because not all of these groups are equally associated with enhanced expression levels in neoplastic growth, we primarily focus on tyrosine kinase receptors (TKRs), G-protein-coupled receptors (GPCRs), cell adhesion molecules (CAMs), membrane-bound enzymes, and GPI-anchored proteins. 7 Although transporter proteins are also reported to be upregulated in cancer cells, they have not been extensively studied for therapeutic tumor targeting yet and are not present in the NCI list. Because the glucose transporter proteins of the GLUT family are actually the targets for the majority of positron emission tomography (PET)-based tumor imaging research, based on the uptake of the glucose analog 18 F-fluorodeoxyglucose (FDG), GLUT1 is included in this overview. Moreover, transporter proteins ABC (ATP-binding cassette), SLC5a* (sodium/glucose cotransporter-5a), and more recently TRPM8 (transient receptor potential cation channel subfamily M, member 8; Fig. 3) seem to have potential for imaging purposes as well.10,11 A number of membrane-associated antigens from the NCI list are not proteins, but these are mentioned in a separate section.
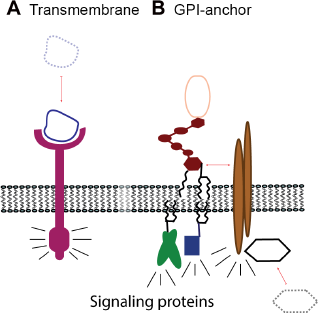
The essential difference in protein attachment to the cell membrane and the impact on signaling between A) transmembrane receptors and B) GPI-anchored receptors. The arrows indicate binding of ligands and/or additional adaptor protein, needed for signal transduction via signaling proteins.
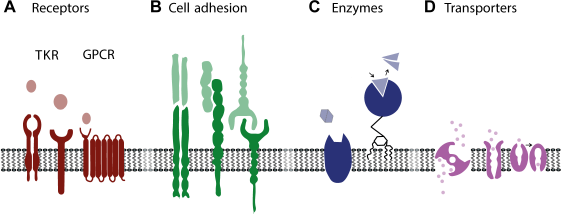
Schematic representation of diverse groups of membrane-associated proteins. (A) Receptors of tyrosine kinase receptor (TKR) and G-protein-coupled receptor (GPCR) subtypes, (B) cell anchoring proteins, C) enzymes, and D) transporter proteins. The respective extracellular ligands, binding proteins, substrates, and transported substances are indicated for each group.
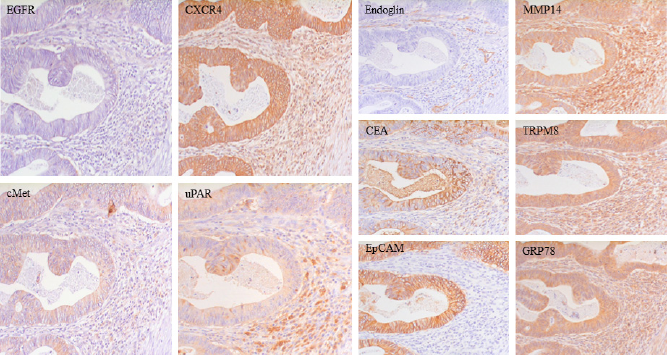
Sequential sections from a representative human colon cancer tissue immunohistochemically stained for various types of membrane-associated proteins. Examples of all the functional groups are presented: tyrosine kinase receptors, epidermal growth factor receptor (EGFR), and hepatocyte growth factor receptor (cMET), C-X-C chemokine receptor-4 (CXCR4) representing the G-protein-coupled receptors (GPCRs), glycosylphosphatidyl inositol (GPI)-anchored receptor urokinase-type plasminogen activator receptor (uPAR), coreceptor endoglin, anchoring proteins carcinoembryonic antigen (CEA) and epithelial cell adhesion molecule (EpCAM), membrane-associated enzyme membrane type-1 matrix metalloproteinase MT1-MMP/MMP14, transporter protein TRPM8 (Transient receptor potential cation channel subfamily M member 8), and the relocalized endoplasmatic reticulum protein glucose-regulated protein-78 (GRP78) (unpublished data, magnification 400x).
Receptors
Tyrosine kinase receptors
TKRs are type I transmembrane receptors that become activated through binding of the extracellular domain with its specific ligand (Fig. 2A). The consequent activation/phosphorylation of the intracellular domain leads to the onset of various pathways, ultimately culminating in a specific response. The 58 known TKRs consist of receptors for growth factors, cytokines, and hormones. A substantial number of TKRs are upregulated in neoplastic tissues, and they play pivotal roles in cancer biology during tumor development, proliferation, angiogenesis, and metastasis. Many studies have explored the potential application of interference of TKRs or their ligands for therapeutic anticancer purposes. Prominent cancer-associated TKRs are the members of the epidermal growth factor receptor family (EGFRs or ErbBs), Eph receptors, fibroblast growth factor receptors, hepatocyte growth factor receptor (HGFR or cMET), insulin-like growth factor (IGF) receptors, and vascular endothelial growth factor receptors (VEGFRs), which are discussed herein.
EGFR (ErbB-1) is upregulated in most cancer types and, therefore, is considered an attractive tumor target (NCI: position
In comparison to the relatively omnipotent EGFR/ ErbB-1, another member from the EGFR family, HER2 (ErbB-2) (NCI
EphA2 (NCI:
VEGFR2 (NCI:
Two other promising TKRs for targeting are the HGFR (cMET; Table 1) and IGF-1 receptor (IGF-1R). The WNT pathway target gene cMET is overexpressed in various tumor types, including colorectal cancer, as illustrated in Figure 3. 18 Several cMet-targeting therapeutic modalities, such as antibodies, hepatocyte-specific inhibitors, small inhibitory molecules, peptides, and even HGF (fragments), the natural ligand of cMet, are already in clinical trials.19–21 This wide range of available interfering compounds is also under evaluation for tumor imaging, with promising results.22,23
Similar to cMet, IGF-1R also is upregulated on the surface of various types of cancer cells. 24 Several monoclonal antibody-based studies have evaluated the therapeutic possibilities. The close resemblance with the insulin receptor and concomitant cross-reactivity might hamper the development of IGF-1R-based therapy. The practical use for imaging purposes is under investigation.25,26
G-protein-coupled receptors
GPCRs, also known as seven-transmembrane domain receptors, are type III transmembrane receptors located completely intramembranously with the exception of the binding loops (Fig. 2A). The GPCRs comprise approximately 3% of the encoded products of human genes, suggesting the existence of approximately 800 human GPCRs. 27 Individual cells can express up to more than 100 different GPCRs that are involved in the regulation of various cellular processes. 28 The ligands that activate receptor response are diverse, varying from hormones to neurotransmitters. Several GPCRs are clearly associated with cancer and, because interfering with GPCRs is relatively easy, one-third of all current therapeutics are actually directed against this type of receptors.5,29 Various clinical trials for GPCR-targeting drugs against different types of cancer are under evaluation, eg, atrasentan, zibotentan, and vismodegib.30,31 Yet, none of these GPCRs are in the NCI prioritization list of cancer targets. C-X-C chemokine receptor-4 (CXCR4), protease-activated receptor (PAR)-1, follicle-stimulating hormone receptor (FSH-R), glucose-regulated protein (GRP), and procaspase activating compound (PAC)-1 seem to be the most prominent upregulated components in breast, head and neck, lung, ovarian, prostate, and colon cancer and are therefore evaluated as candidates for imaging applications. 32 CXCR4 (Table 1) is the receptor for SDF1/CXCL12 and is implicated in the cross talk between (tumor) cells and their microenvironment. Overexpression has been found in all major tumor types, including colon cancer (Fig. 3). 33 Antibodies and peptides against CXCR4 are extensively being evaluated for molecular imaging purposes, with promising results. 34 PAR-1, a member of the protease-activated receptor subfamily, and FSH-R are mainly overexpressed on endothelial cells within the tumor environment and would therefore also be present in a broad range of tumors. PAR-1 has already been evaluated for in vivo imaging of angiogenic endothelial cells and malignant cells. 35 The abundant expression of FSH-R in angiogenic blood vessels, as recently shown in large numbers of tumors, would make this receptor a broadly applicable tumor-detecting target for the imaging of especially larger tumors.36,37
A number of GPCRs, including the receptors for somatostatin, gastrin, bombesin, and cholecystokinin (CCK), are already being clinically exploited for the diagnosis of neuroendocrine tumors (Table 1). Neuroendocrine tumors consist of a heterogeneous group of neoplasms, including carcinoids, pancreatic islet tumors, paragangliomas, and neuroblastomas, for which surgery is the only curative treatment. Accurate localization with radiolabeled peptide analogs of somatostatin, gastrin, bombesin, and CCK is used to target their respective GPCRs.38,39 The adaptation of these tracers with near-infrared (NIR) labels might lead to a wider application in cancer imaging, possibly also for more common tumors. 40
Cell adhesion molecules
CAMs form a large and diverse group of membrane-bound proteins that are defined as morphoregulatory molecules that affect cellular processes. The definition indicates that these proteins are not implicated only in cell-cell or cell-matrix adhesion as the name suggests (Fig. 1B).
Carcinoembryonic antigen (CEA; NCI:
Two CAMs that are highly associated with tumor proliferation and overexpression are EpCAM (epithelial cell adhesion molecule) and E-cadherin. EpCAM (NCI:
Another important family of CAMs are the integrins. They consist of heterodimer receptors involved in the regulation of the cell cycle, cellular shape, and motility, due to their interaction with other cells and with the extracellular matrix. 61 Specifically, integrins containing the alpha subunit are abundantly overexpressed in tumors, both on angiogenic endothelial cells, eg, αvβ3 (Table 1), as well as on malignant epithelial cells, eg, αvβ3, αvβ5, and αvβ6.62–64 Targeting of a number of integrins simultaneously via an arginyl-glycyl-aspartic acid (RGD) peptide is well established and is being explored for both tumor imaging and therapeutic purposes.65–67
Cell membrane-associated enzymes
An interesting group of possible tumor targets are the membrane-associated enzymes (Fig. 1C). This is a rather heterogeneous group of proteins primarily involved in the maintenance of cellular functions, uptake/secretion/proteolysis of proteins, and extracellular matrix remodeling. They are important for the homeostasis of the cell and regulate cell-cell and cell-matrix contacts. Upregulation under neoplastic conditions has been reported for a number of these enzymes, especially the proteases that are involved in extracellular matrix remodeling during migration and invasion.
Glutamate carboxypeptidase 2, also known as folate hydrolase 1 (FOLH1) or prostate-specific membrane antigen (PSMA; NCI:
Another type II membrane-bound peptidase that is investigated as a potential tumor target is aminopeptidase N, also known as CD13 (Table 1). This enzyme is abundantly expressed on fast-growing angiogenic endothelial cells but is also present on tumor cells. 74 Aminopeptidase N serves as a receptor for Asn-Gly-Arg (NGR) peptide(s). NGR peptides are intensively evaluated as a tumor target for both therapy and imaging.75,76 Clinical trials with NGR peptides conjugated to toxins or antitumor cytokines such as tumor necrosis factor (TNF) are under investigation. 77 As for PMSA, imaging of aminopeptidase N could be established by binding of a determinant to the protein, such as NGR peptide, but could also be based on the local proteolytic activity of the enzyme.78,79
Furthermore, overexpression of seprase and matriptase, two members of the transmembrane serine protease family, has been associated with several tumor types, including breast, colon, ovary, and prostate cancer.80,81 Seprase or fibroblast activation protein (FAP-α; NCI:
Matriptase (membrane-type serine protease 1, MT-SP1, Table 1) is enhanced in several tumor types, where it is suggested to play an active role via the activation of HGF and urokinase plasminogen activator. 84 NIR fluorescence and radiolabel imaging of antibodies against the active form of matriptase showed a tumor-specific signal in animal models, indicating that these membrane-bound enzymes, as well as their activities, could be used for tumor imaging. 81
The matrix metalloproteinases (MMPs) and the ADAMS (A disintegrin and metalloprotease domain) are the most prominent families of invasion-associated proteases. Two transmembrane members, membrane type-1 matrix metalloproteinase (MT1-MMP)/MMP14 (Fig. 3) and ADAM12, have been found to be upregulated in various types of cancer. 85 Targeting of MMP14 with a radiolabeled antibody confirmed the potential of this membrane protein as a tumor target. 86 As already indicated for PSMA and aminopeptidase N, an advantage of choosing proteolytic enzymes as a tumor target is the possibility of making use of their main feature, ie, activation of substrates. Several targeting drugs and imaging probes have been developed using upregulated membrane-bound or membrane-associated proteolytic enzymes, such as MMP-2, MMP-7, and MMP-9, for localized activation.87,88 Recently, first-in-human data have been presented for a cathepsin-activated probe, underscoring the potential of this approach. 89
Proteolytic enzymes are not the only molecules studied as tumor-specific targets. Carbonic anhydrase nine (CAIX; NCI:
Transporter proteins, mucins, and other membrane-associated proteins
Transporter proteins
Due to their changed physiology, most malignant cells show enhanced metabolic activity. Cell membrane transporter proteins such as GLUT, ABC, and SLC5a* are upregulated on cancer cells, compared to adjacent normal cells. High levels of glucose transporters GLUT1/ GLUT3/GLUT12 are found in a wide range of solid tumors correlating with poor survival. 94 Overexpression of GLUT proteins is often associated with hypoxic conditions and is generally present in perinecrotic areas within the tumors. Being the main glucose transporters, the GLUT proteins are the targets of one of the most commonly used oncologic PET tracers, FDG (2-deoxy-2-[18F]fluoro-D-glucose). Although efficient for PET imaging of many tumor types, due to the commonly encountered high uptake in normal tissues and the expression pattern within the tumor, GLUT proteins seem not particularly suited for NIRF imaging.
Although aberrant expression of channels for ions such as Ca, Na, and K is a common feature of tumor cells, there are relatively few studies dedicated to the use of these membrane proteins for tumor targeting yet.10,11 An example of a potential candidate for tumor targeting is TRPM8.95,96 TRPM8 is aberrantly present on various types of cancer cells, including breast, lung, colorectal and pancreatic cancer, where it seems essential for the expanding growth of several types of tumors (Fig. 3).
Mucins and other difficult-to-categorize membrane-associated proteins
The highest ranked membranous protein in the NCI prioritization list is epithelial membrane antigen (EMA), also known as mucin-1 (MUC-1; NCI:
Some proteins that are generally located within the cell form membranous counterparts under tumor conditions. An intriguing example of the latter is GRP78, a member of the Hsp70 heat shock protein family, also known as BiP. Under normal conditions, GRP78 is present in the endoplasmatic reticulum (ER). Hypoxia or glucose-deprived conditions, as often found in poorly perfused tumor tissues, lead to upregulation of GRP78 and relocalization from the ER into the cytoplasm and the cell membrane, where it functions as a (co)receptor by binding to other membrane proteins.108,109 Enhanced membranous GRP78 has recently been recorded in most cancer types (Fig. 3), often associated with bad prognosis.110–112 Because of the tumor-associated membranous expression, GRP78 is regarded as a recognition element for cancer drug targeting, and antibodies are recently being evaluated for use as PET tracer for pancreatic cancer.111,113
Endoglin (CD105, Table 1) is a type I transmembrane glycoprotein (co)receptor for various ligands, including bone morphogenetic protein (BMP)-9 and transforming growth factor (TGF)-β, but it does not induce intrinsic kinase activity. Therefore, endoglin does not belong to either the TKR or the GPCR families of receptors. Apart from soluble ligands, endoglin also interacts with high affinity to membrane proteins such as the TGF-β receptors and various integrins. 114 Enhanced endoglin levels seem essential for angiogenesis during tumor development. Because of the low expression in normal tissues, endoglin on tumor-associated neoangiogenic cells is being explored as the target for a wide range of solid tumors (Fig. 3). Monoclonal antibodies against endoglin are being evaluated for anticancer therapy and imaging purposes. Multiple Phase II clinical trials are ongoing with TRC105, an endoglin-neutralizing antibody binding to the extracellular endoglin orphan domain, as antiangiogenic therapy. 115 In addition, TRC105 has also been preclinically applied for the imaging of tumor vasculature in mice cancer models. 116
In addition to proteins in the cell membrane, a number of nonproteins, such as glycans and phospholipids, are being explored or evaluated as candidate tumor targets. Examples that are associated with tumor targeting are CA19–9, CD77, fucosyl GM1, gangliosides, polysialic acid, GD3, sLea/sLex, sialyl-Tn antigen, and phosphatidylserine.
GPI- and lipid-anchored proteins
GPI- and lipid-anchored proteins are a relatively small and heterogeneous group of proteins, consisting of receptors and adhesion molecules, which cannot easily be integrated within the conventional classification systems (Fig. 2). They share solely the mode of attachment to the cell membrane. 117 We discuss this group separately in this overview because a relatively large number of GPI-anchored proteins are associated with cancer, among which the already-discussed CEA is the most prominent. 118 Here, we highlight other cancer-associated GPI proteins, namely, mesothelin, prostate stem cell antigen (PSCA), and the receptors for urokinase and folate.
Mesothelin (NCI:
PSCA (NCI:
uPAR (Table 1) localizes the proteolytic activity of urokinase, important for matrix degradation, but binding of urokinase to its receptor also results in cell signaling. Being a GPI-anchored protein and therefore lacking an intracellular domain, the signaling functions of uPAR are mediated by interactions with other membrane proteins, such as integrins (eg, α5β1), TKRs (eg, EGFR), GPCRs (eg, CXCR4), and matrix components such as vitronectin.123,124 Upregulation of uPAR levels has been found in the majority of tumor types 124 and was associated not only with malignant cells but also with macrophages, neutrophils, and endothelial cells within the tumor microenvironment. 125 Therefore, uPAR is being extensively studied as a target for cancer therapy and imaging using antibodies, peptides, as well as the amino terminal fragment derived from the natural ligand urokinase.126–129 First-in-human results have been presented recently.130,131
The folate receptors (FRs) are a set of two GPI-linked membrane proteins (isoforms α and β) absent in most normal tissues but frequently observed in various types of human cancers. FR-α has been considered a target for cancer therapy for more than a decade.132–135 Recently, various studies have used the natural ligand folate/folic acid, conjugated with NIRF and radioactive labels, for the imaging of various types of human tumors in animal models,136,137 culminating in the first-in-human imaging studies in ovarian cancer patients.138,139
Two members of the previously mentioned metalloproteinase family, MT4-MMP/MMP17 and MT6-MMP/ MMP25 are also GPI-anchored moieties and they are upregulated in various cancer types, wherein they are associated with tumor progression.140,141 The localized proteolytic activity of both MT-MMPs, especially at the interface between tumor and stromal cells, contributes to remodeling of the extracellular matrix, enabling metastatic dissemination.142,143 Although MMP-activated prodrugs are being investigated for tumor therapy and tumor imaging, they are not specifically designed for GPI-anchored MT-MMPs, lacking (tumor cell) specificity.
Characteristics Defining Good Tumor Targets
In the previous section, a large number of tumor-associated cell membrane proteins have been discussed. Although they all have their specific pros and cons, not all of these proteins are equally good targets for therapy or imaging. Some generic characteristics for target tumor proteins have been defined recently.7,144 The most obvious criteria, besides the localization on the cell membrane, are the exclusive upregulation on cells within the tumor compared to adjacent normal tissue cells, the actual number of targeted proteins available per cell, and the percentage and distribution of positive cells within the tumor (Fig. 4). Another convenient but not essential criterion would be the presence on a wide variety of tumor types. The presence of high levels of soluble forms of the protein in the circulation could be regarded as disadvantageous. On the other hand, the presence of these proteins in the circulation could be used as indicators for the expression on the tumor, as indicated in Figure 4. In the next part of this overview, some key characteristics of good tumor target proteins will be discussed.
Number of target proteins per tumor cell
Upregulation of the number of target protein molecules is important for distinguishing tumors from normal tissue counterparts (Fig. 4). Two- to 100-fold upregulation levels have been reported for various cell membrane tumor markers. Rough estimations of the total copy numbers of membrane markers per (tumor) cell indicate that there are large differences between proteins and within the various groups of membrane proteins (Table 1). For targeting purposes, upregulation on tumor cells is only relevant if this culminates in significantly high(er) protein numbers per cell compared with cells in the adjacent normal tissue. A recent in vitro study has established a threshold for effective HER2 therapeutic targeting, starting from 2 x 10 5 receptors per cell. 145 Because HER2 overexpression is due to a genetic amplification of up to 50 gene copies, the number of HER2 molecules on positive tumor cells is 40- to 100-fold upregulated, culminating in levels of over 10 6 copies per cell. With this number, HER2 ranks among the highest expressed membrane proteins, which–-together with the low expression levels in nonmalignant cells–-renders it an ideal target, but unfortunately only in a relatively small percentage of tumors. 13 Especially for tumor-imaging purposes, the actual number of copies per tumor cell is probably less important than the ratio of copies between tumor cells and normal cells. Because the number of EGFRs on normal cells is between zero and 40,000 depending on the tissue type, the upregulation on cancer cells to a maximal 10 5 molecules per cell (Table 1) would result in, for some normal tissues, only marginally enhanced levels, whereas for other tissues, this ratio will suffice. 146 Table 1 gives an estimation of the number of copies per cell for many of the proteins discussed in this overview.
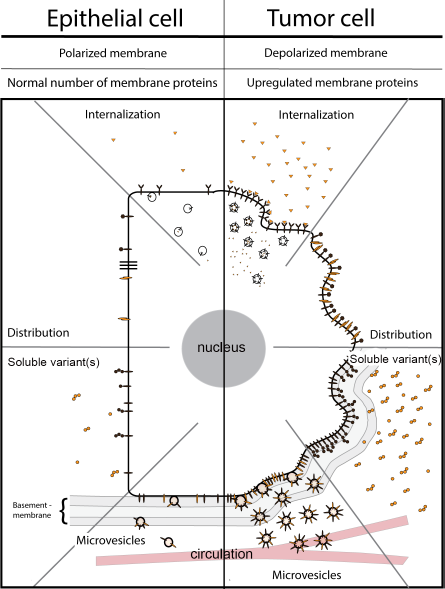
Schematic overview of membrane proteins on normal polarized epithelial cells (left) versus their counterparts on malignant tumor cells (right). The number, distribution, and conformation of cell membrane proteins on normal cells are determined by variables such as presence of ligands, internalization, shedding, and microvesicle formation. Although cancer cells often show enhanced expression of tumor-associated membrane proteins, the suitability as target for imaging of these proteins is often hampered by a changed distribution profile, increased internalization, shedding, and/or microvesicle formation.
Availability/accessibility of the target on the cell membrane
All the discussed membrane proteins are in principle present in enhanced numbers on the membrane of the cell. However, many of these proteins do also have intracellular and/or extracellular variants (Fig. 4). The presence of both variants is not advantageous for tumor targeting. Intracellular forms are not directly accessible and reduce the number of membrane proteins per cell, except when internalization has occurred after targeting. Internalization is particularly associated with membrane receptors after binding of their specific ligand, but it can also occur with antibodies or other targeting probes. Extracellular forms of membrane proteins can originate from alternatively spliced variants but can also originate from the original membrane proteins after cleavage from the membrane, a process called shedding. These soluble receptors are still capable of binding ligands or antibodies, targeting the latter in the circulation, resulting in the need for higher doses, as indicated for CEA and EGFR.44,147 In addition to occurring as soluble proteins, extracellular membrane proteins are also present in the circulation on membrane particles called microvesicles (Fig. 4). Microvesicles are particles shed by (tumor) cells ranging in size from 100 nm to 1,000 nm in diameter. They consist of the membrane and cytosolic contents of their parental cells and generally arise from an unspecific spontaneous process. Involvement of microvesicles is indicated in cardiovascular disease, rheumatic arthritis, and cancer. 148 Many of the tumor cell-associated membrane proteins have been identified on microvesicles in the blood of cancer patients. Similar to their soluble counterparts, these membrane proteins scavenge a percentage of the targeting probe.
Even abundant presence on the cell membrane does not necessarily guarantee easy accessibility of a target. Determining factors are the solubility and the clustering of the protein within the membrane, the polarization state of the cell, the presence of various forms of the same protein, and the binding of these proteins to other proteins. All membrane proteins are to some extent able to float freely through the membrane bilayer of a cell, but some are more fluid than others. Because GPI anchors do not completely extend through the plasma membrane, GPI-anchored proteins belong to the most diffusive proteins on the cell surface, allowing a rapid response to external stimuli. 149 High membrane solubility and the highly associated cluster formation in microdomains are considered advantageous for tumor targeting. An example is the GPI-anchored FR-α. After the binding of folate, the receptor clusters in specific cellular membrane subdomains, followed by endocytosis. Intracellular folate dissociates from the receptor and is translocated into the cytoplasm, whereas the receptor recycles rapidly back to the cell membrane, available for the next ligand/probe. 150 Folate derivatives are therefore extensively studied/explored as probes for cancer therapy or imaging.
Normal epithelial cells are strongly polarized, with an apical side and a basolateral side. The distribution of most membrane proteins is strongly dependent on the function, eg, adhesion molecule EpCAM, which is primarily present at cell-cell and cell-matrix contact points. When epithelial cells differentiate to migrating cancerous cells, they generally lose their polarized structure, indicating that specifically arranged membrane proteins are not restricted to the different sides anymore but become distributed throughout the entire plasma membrane. 48 The conversion of a protein into various confirmation states might also hamper the traceability. For instance, the three-dimensional appearance of receptors changes considerably after the binding of a ligand, affecting the affinity of an antibody or peptide drastically. Adhesion molecules, on the other hand, have various activation states, which also influence the affinity for the targeting probes. 151
Considerations to Select the Optimal Target for Tumor Imaging in the Clinic
Although tumor imaging during an operation is considered a very powerful technique, the quest for optimal probes/targets for this application has only just begun. In the past 10 years, numerous tumor-targeting NIRF probes are, or have been, evaluated in preclinical animal models with generally good results. The translation of these results into the clinic however, is another story. A recent search for clinical trials using NIR probes revealed almost 80 hits (May 2016, clinicaltrials.gov, near infrared AND cancer/tumor). However, the vast majority of these studies are investigating the use of indocyanine green, a nontargeted dye, to detect sentinel lymph nodes in a wide range of tumor types. Only eight trials are actually using targeted probes, the majority being based on only two therapeutic antibodies targeting VEGF and endoglin, both predominantly present on (tumor) angiogenic cells. These studies are presumably just the tip of the iceberg still to come but indicate the relative immature status of the matter.
This overview of the different groups of cell membrane proteins and possible candidate members for tumor targeting shows that, in principle, many different proteins could be used and that most of them will work to a certain extent, at least in preclinical models. Selecting the best probes from among these candidates requires a systematic comparison of the performance of candidate targets in suitable model systems, preferably for various tumor types. Whether currently available mouse tumor models provide the proper tools to make adequate selections/comparisons of probe/target combinations is still questionable.
Unfortunately, the choice of a target protein is only the beginning. Various studies indicate that antibodies to the same protein show very different results on imaging, depending on the domain of the protein that is targeted or the affinity of the antibody for the protein. Apart from antibodies, there is a whole range of alternatives available, each with specific (dis)advantages. Recently published clinical studies with an antibody (bevacizumab), a natural receptor ligand (folate), and a peptide (cMET) suggest that tumor-targeting agents that were originally designed for cancer therapy might be useful for imaging purposes as well.23,152,153 Many of these targeting therapeutic agents are now being evaluated for imaging. These agents have been already evaluated in the clinic for side effects and doses, albeit without the NIRF dye or radiolabel being attached. Therefore, these therapeutic agents warrant a presumably faster translation into the clinic than newly designed imaging probes.
Overall Conclusion and Future Perspective
It should be clear from the previous paragraphs that the ultimate target for tumor targeting has not been found or does not exist. It is even impossible to indicate how it should look like or which subgroup of cell membrane proteins it should belong to. We do not even have proper tools/models to compare the targets/agents properly.
The fact that the majority of the priority proteins from the NCI list belong to the TKR subgroup indicates that these receptors are apparently suited for targeted therapy and/or imaging. The main reason is the large number of these receptors on tumor cells (Table 1). However, this by no means implies that all these receptors are always upregulated in the majority of tumors, as indicated for EGFR in Figure 3. Although one of the most robust targets, for colon tumor, EGFR is clearly outperformed by cMET, another member of the TKR subgroup, and also by CXCR4, MMP14, CEA, TRPM8, and GRP78, members of the GPCR, enzyme, adhesion molecule, transporter protein, and heat shock protein families, respectively.
Despite the prominent presence of soluble counterparts in the circulation, GPI-anchored proteins seem particularly good tumor targets, represented in Figure 3 by CEA, FR, and uPAR. In comparative studies, CEA is generally among the best markers for colorectal tumors and other cancer types. 46 CEA shares this multitumor type distribution with another non-GPI-anchored adhesion molecule EpCAM (Fig. 4A). FR and uPAR are also good examples of pluripotent tumor targets. Although uPAR expression is probably relatively low, it would have the extra advantage that upregulation is not limited to cancer cells only but also extends to stromal tumor cells such as angiogenic endothelial cells and macrophages (Fig. 3).
Targeting of tumor stromal cells rather than malignant cells has great potential, especially for tumor imaging. The initial studies on stromal tumors concentrated primarily on angiogenic endothelium, which is present in the periphery of many tumors. Especially, the αvβ3 integrin has been successfully targeted in many preclinical and clinical studies. In terms of numbers per cell, this adhesion molecule is a rather low-expression-level membrane protein (Table 1), indicating the power of targeting angiogenic cells for tumor imaging. With 10 times more proteins per cell membrane, the TGF-β coreceptor endoglin seems a promising alternative for αvβ3 for angiogenic endothelial targeting. 154
In conclusion, in the coming years, an enormous boost is expected from the rapidly advancing imaging techniques. The global optical imaging market is expected to reach US$1.9 billion by 2018. 155 These advanced camera systems will rely on high-quality targeting tracers. Better understanding of membrane proteins and their characteristics will be essential for the development of these compounds, regardless of whether the target is cancer or any other disease. The concept that one probe targeting one single protein will be able to detect all tumors seems too optimistic. Therefore, selecting the right patient/probe combination is of key importance. 156 Circulating soluble forms of targetable membrane proteins could offer a convenient noninvasive way to confirm the presence in the tumor. As previously mentioned, GPI-anchored membrane proteins, such as CEA, FSH-R, and uPAR, shed relatively easily from the cell membrane, which could be an advantage. Most research is focused on single target approaches, but the use of these types of probes has the intrinsic property that they are limited in their applicability for specific patient subgroups. 156 Recent developments suggest that approaches such as agents targeting two proteins, or alternatively two different epitopes on a biomarker (biparatopic), result in considerably higher affinity, specificity, and sensitivity and address the problem of intratumoral heterogeneity.157,158
Abbreviations
ATP-binding cassette
A disintegrin and metalloprotease domain
Amino terminal fragment
Binding immunoglobulin protein
Carbonic anhydrase-9
Cell adhesion molecule
Chole-cystokinin
Carcinoembryonic antigen
C-X-C chemokine ligand-12
C-X-C chemokine receptor-4
Hepatocyte growth factor receptor
Designed ankyrin repeat proteins
Ductal carcinoma in situ
Epidermal growth factor (receptor)
Extracellular matrix metalloproteinase inducer
Epithelial cell adhesion molecule
Ephrin receptor
Endoplasmatic reticulum
Fibroblastic activation protein-α
Fluorodeoxy glucose
Formyl-methionyl-leucyl-phenylalanine
Folate receptor-α
Follicle-stimulating hormone (receptor)
Glucose transporter
G-protein-coupled receptor
Glycosylphosphatidyl inositol
Glucose-regulated protein-78
Human epidermal growth factor receptor-2, ErbB-2
Hepatocyte growth factor receptor
Heat shock protein-70
Insulin-like growth factor-1 receptor
Matrix metalloproteinase
Mucin-1
National Cancer Institute
Asn-Gly-Arg peptide
Near-infrared fluorescence
Procaspase activating compound-1
Protease-activated receptor-1
Prostate stem cell antigen
Positron emission tomography
Prostate-specific membrane antigen
Radioactivity
Arginyl-glycyl-aspartic acid
Single-chain variable fragment
Stromal cell-derived factor-1
Sodium/glucose cotransporter-5a
Tumor-associated glycoprotein-72
Transforming growth factor-β
Tyrosine kinase receptor
Transmembrane
Tumor necrosis factor-α
Transient receptor potential cation channel subfamily M member 8
Urokinase-type plasminogen activator receptor
Vascular endothelial growth factor (receptor).
Author Contributions
Conceived and designed the content of the manuscript and the figures: MCB, SWLdeG, HAJMP, and CFMS. Provided immunohistochemical illustrations: HAJMP. Wrote the first draft of the manuscript: SWLdeG and MCB. Contributed to the writing of the manuscript: LJACH and PJKK. Made critical revisions and approved the final version of the manuscript: MCB, SWLdeG, LJACH, CJHvdeV, PJKK, ALV, and CFMS.