
Abstract
Mitochondria are subcellular organelles that provide energy for a variety of basic cellular processes in eukaryotic cells. Mitochondria maintain their own genomes and many of their endosymbiont genes are encoded by nuclear genomes. The crosstalk between the mitochondrial and nuclear genomes ensures mitochondrial biogenesis, dynamics and maintenance. Mitochondrial proteins are partly encoded by nucleus and synthesized in the cytosol and partly in the mitochondria coded by mitochondrial genome. The efficiency of transport systems that transport nuclear encoded gene products such as proteins and mRNAs to the mitochondrial vicinity to allow for their translation and/or import are recently receiving wide attention. There is currently no concrete evidence that nuclear encoded mRNA is transported into the mitochondria, however, they can be transported onto the mitochondrial surface and translated at the surface of mitochondria utilizing cytosolic machinery. In this review we present an overview of the recent advances in the mRNA transport, with emphasis on the transport of nuclear-encoded mitochondrial protein mRNA into the mitochondria.
Introduction
Mitochondria, the major energy providers for eukaryotic cells, are also involved in a variety of basic cellular processes such as lipid metabolism, buffering for calcium ions and the regulation of apoptosis in higher organisms. Mitochondria are considered to have evolved from eubacterial endosymbiotic progenitor and their genomes closely resemble that of the prokaryotic form,1,2 such as compact arrangement of genes and absence of introns. 3 Although mitochondria still maintain independent gene expression systems, only about a few protein coding genes (i.e. coding for respiratory complex) retained and most of them are thought to have migrated to host genome. To reconstruct the functional complex, the organelles have to import products coded in the nuclear genome. Through relatively comprehensive proteomics studies in yeast mitochondria, it has been proven that about 99% of mitochondrial proteins are translated in the cytoplasm and imported into the mitochondria by different mechanisms. 4 Other studies have shown that in a typical somatic human cell, there are about 100-10,000 copies of mitochondrial DNA, which encode two ribosomal RNAs, 22 tRNAs and 13 genes encoding subunits of the respiratory chain enzymes located in the inner mitochondrial membrane. 5 These proteins include seven NADH dehydrogenase subunits (ND4L and ND1-ND6), three cytochrome c oxidase subunits (COX1-COX3), a subunit of complex IV cytochrome b (CYB) and two subunits of the mitochondrial ATPase 6 and ATPase 8 in human cells. The remaining subunits are encoded by the nuclear DNA.3,6 Besides, A majority of proteins involved in mitochondrial replication, transcription and translation are also coded by nuclear genes. 3
Recently, many studies have been focusing on the importing process of mitochondrial proteins.7–10 However, the mechanisms by which the nuclear coded mitochondrial products are transported, how the gene are selected and targeted for transport remain unclear. Most mRNAs of nuclear-coded mitochondrial proteins are translated in the cytoplasm and possibly transported in form of nascent polypeptide-associated complex (NAC).11,12 In studies of lower eukaryotes, some proteins are found to be co-translationally imported into the mitochondria via mitochondrial surface bound ribosomes,13,14 suggesting existence of another form of transport mechanism, that is the transport of mRNA. The mRNA transport process is a relatively new discovery and now receiving wide attention because of several reasons: (a) moving mRNA to mitochondria is an efficient way to get multiple products in the mitochondria by using a single mRNA template. By transporting mRNA into the mitochondrial vicinity, cells achieve savings of the cost to transport each copy of the translated protein. For example, cofactors such as ATP and the molecular motors are some of the costs for precise transportation of most cargos, including protein and RNAs. There are multiple copies of mitochondria in one cell, which require multiple copies of proteins to construct fully functional mitochondrial machinery. It is unlikely that all the mitochondrial components are delivered in protein form because this would probably be limited by the amount of cofactors. (b) Translation is a process that consumes most energy as cells spend more energy on protein synthesis than on any other activity. 15 Regulating mRNA location before translation is a faster and energy-conservative way to respond to environmental changes. (c) mRNA localization might facilitate assembly of multifactor complexes by bringing the different protein subunit templates to same location.
Several questions pertaining to mRNA transport to mitochondria remain to be addressed in a satisfactory manner. For example, what are the known common properties of mRNAs translocated to mitochondria? What are the modes of export of mitochondrial mRNA from the nucleus? How do the exporter proteins interact with mRNAs cargos in the ribonucleoprotein (RNP) complex? How do the RNP complexes interact with mitochondria? In this review we present an overview of recent advances on the mRNA transport, with emphasis on transport to the mitochondrial surface.
Review
mRNA export from nucleus
In Eukaryotic cells, most mRNAs are transcribed within the nucleus. Generally, the nascent mRNA is co-transcriptionally packaged with transacting proteins into a messenger ribonucleoprotein particle (mRNP), and is subsequently exported from the nucleus through the nuclear pore complex (NPC), as depicted in a simple schematic diagram (Fig. 1). The majority of nucleocytoplasmic transport factors for proteins and RNAs belong to karyopherin β family (importin-β like proteins), which use Ran cycle to achieve the exchanging cargo with the help of some adaptors. 16 Ran is a small GTPase. Ran exists in two states that interact differently with other proteins depending on whether it is bound to GTP or GDP. Ran-GTP has higher affinity to karyopherin β family. Ran-GTP is converted to Ran-GDP by a GTP activating protein (Ran-GAP), while Ran-GDP cycles back to Ran-GTP by Regulator of Chromosome Condensation 1 (RCC1) protein. 17 Ran-GDP releases cargos in the cytoplasm and in the process GTP is hydrolyzed to GDP. Instead of Ran and importin-β like proteins, the mRNA utilizes a different way of cargo exportation as well. We will briefly discuss the general mechanism of mRNAs export from the nucleus in following sections.
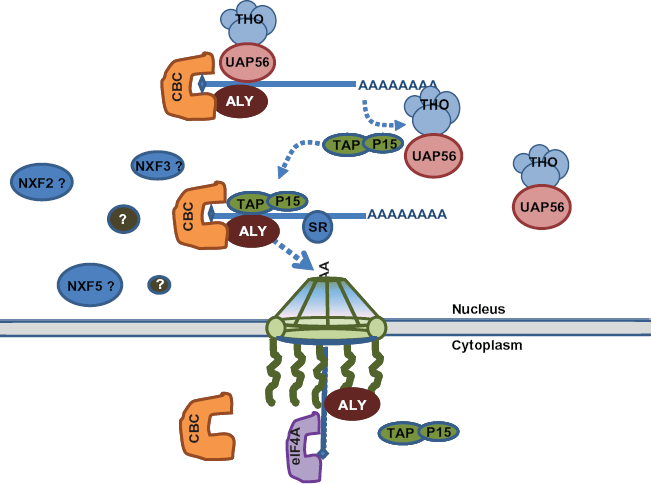
Simplified schematic diagram of some major known proteins and interactions in nuclear coded mRNA export process. mRNA are transcribed from the DNA (not shown) and assembled into RNP complex. TAP/P15 heterodimer is recruited to mRNA by adaptor Aly, and are replaced the THO/UAP5 complex. The newly formed RNP complex passes through the NPC, and CBC is replaced by eIF4A cap binding protein in the cytoplasm. Polyadenylated mRNA is represented by a line with a poly A tail. Question marks indicate cofactors that are known to exist in specific regulation of mitochondrial mRNA but yet to be identified. Factors that have been identified but for whom the specific interactions remains to be elucidated bear question marks at their end. Such factors are involved in processes including targeting, translocation and selectivity.
One of the key players in mRNA export has been identified as Nuclear RNA Export Factor (NXF1) in S. cerevisiae and Tip-associating Protein (TAP) in mammalians. TAP (NXF1) associates with P15, known as a stimulator of nuclear export signal containing proteins, 18 as well as RNAs. 19 Certain RNA sequence, usually in the untranslated region (UTR), functions as cis element, which associates with cofactors to achieve specificity for export. A constitutive transport element (CTE) is one such example, that is required for export of retroviral transcripts. 20 TAP (NXF1) has been shown to bind to CTE. N-terminal region of TAP (NXF1) interacts with other cofactors such as exonic junction complex components (U2AF, 9G8, etc).21,22 Aly/REF, which is a component of THO/ TREX complex, was shown to recruit TAP (NXF1) to mRNA. Subsequently, TAP (NXF1) docks the mRNA to the nucleoporins, one of nuclear pore components, by C-terminal region. 23–25 Additional NXF like proteins such as NXF2, NXF3 and NXF5 are also proved to be involved in mRNA transport. Some of them have shows activity in mRNA export assay, whereas some such as human NXF3 are inactive. 26 Similar studies in Caenorhabditis elegans showed that Ce-NXF2, the homologue of NXF2, participates in posttranscriptional regulation of tra-2 mRNA, which is required for female development,27,28 and in human, a NXF5 nullisomy is linked to mental retardation. 29 These studies demonstrate importance of NXF family proteins in normal functioning of the cell.
Generally, TAP (NXF1):p15 heterodimer are considered to be involved in most mRNA export from nucleus, 28 and the selectivity of mRNAs targeting at different locations in cytoplasm is probably achieved by the sequence or structure of the RNA, and proteinprotein interactions. However, the precise mechanistic details of mRNA export remains unclear, for example how nuclear coded mitochondrial mRNAs are exported is yet to be understood. Future comprehensive proteomic studies will identify other cofactors that specifically bind to mRNA. This will include the factors that are involved in the transport of the nuclear coded mitochondrial mRNA.
Similar to that in the nucleus, transport of mRNA in the cytoplasm is also determined by the primary structure of RNA and cofactors. In this respect it is important to understand the roles of mRNA binding cytoplasmic proteins and their properties. An example of the regulation of mRNA export comes from the studies in mouse model. It has been demonstrated that in mouse NXF2, NXF7, and TAP (NXF1) binds to the light chain of brain specific microtubule-associated protein (MAP1B), which interacts with cytoplasmic microtubules and actin filaments in cytoplasm and participates in the development and function of the nervous system. 28 Mouse NXF7 also co-localizes with the marker of neuronal RNA transport granules Staufen1 (stau1) protein and ribosomal RNA (rRNA). Another NXF-interacting factor is identified as a member of WD-repeat protein family Unrip/STRAP.28–30 Further studies showed human Unrip protein participates in the inhibition of transforming growth factor β signaling by direct binding to its unrip receptor. Unrip was proposed to act as a scaffold for the assembly of protein complexes or anchor such complexes to specific subcellular locations by its WD-repeat sequence.
Mitochondria contain their own translational apparatus where mitochondrial DNA encoded proteins are translated. In several species (yeast, higher plant and marsupials), it was found that some tRNAs coded by the nucleus is indeed transported into the mitochondria.31,32 These observations have added the tRNAs to the list of macromolecules transported into the mitochondria, in addition to, proteins. Besides, 5s rRNA has also been demonstrated to be imported into mammalian mitochondria. 33 The import of RNAs into the mitochondria (tRNAs and rRNAs) in different species is now well documented, however, no direct evidence exists as yet for import of mRNAs. 1 In yeast, a series of experiments demonstrated that some cytoplasmic ribosomes are bound to the surface of mitochondria,34,35 indicating that the several mRNAs might be transported to and translated in the vicinity of mitochondria.
We will discuss some reported examples of class of mRNAs that is translocated to mitochondria instead of their translated NACs.
Different types of microarray analyses reproducibly showed that about 50% of the mRNAs coding for products localized in mitochondria were primarily associated with polysomes that were bound to mitochondria. 36 In vivo green fluorescent protein (GFP)-labeling experiments have consistent shown that some mRNAs are actually localized in the vicinity of mitochondria.37,38 Mitochondrially-localized mRNA (MLR) is used to define the subclass of mRNA encoding mitochondrial proteins that are translated at the vicinity of mitochondria. MLRs were identified for many yeast genes by genome wide analysis of mRNAs and evaluating the correlation of MLR region by diverse bioinformatic searches. 39 Further analysis demonstrated that MLR proteins are mainly of prokaryotic origin, and are linked to the first step of complex core construction. They are also found to be imported principally through the translocase of outer membrane (TOM) and the translocase of inner membrane (TIM) pathway. 40 However, not much is yet known about the molecular basis of the mRNA translocation to mitochondria.
ATM1 is one of the earliest mRNAs demonstrated to be located at mitochondrial vicinity in yeast. ATM1 encodes an iron-sulfur clusters transporter protein, which belongs to ATP binding cassette complex of the mitochondrial inner membrane. The ATM1 mRNA was one of the five mRNAs (ATM1, COX10, TIM44, ATP2, and COT1) which are identified from mitochondria associated polysomes mRNAs, and this observation was further confirmed by fusing the ATM1-3′ untranslated regions (UTRs) fragment with CP (coat protein form the MS2 bacteriophage) binding sites in the cells expressing CP-GFP fusion protein. The 3′ UTR of ATM1 is demonstrated to be sufficient to target the mRNA to the mitochondria by tracking the GFP signal in vivo and by Western blotting analysis.
ATP2, which encodes an important subunit of ATP synthase F1 subunit, is another mRNA that was found to be translated at mitochondrial vicinity. Although the precursor of ATP2 could be imported posttranslationally, 41 its preferred manner of import was not clear. A similar experiment have shown the localization of ATP2 in the mitochondria vicinity. In contrast, when 3′UTR region was replaced with ADH1-3′UTR (an mRNA encoding Alcohol dehydrogenase, a cytoplasmic protein which is required for the last step in the glycolytic pathway), the amount of mitochondrial ATP2 protein decreased considerably. In a different study, using RNA labeling technique, a conserved region of 3′ UTR of ATP2 was identified, which is sufficient to import mRNA to the mitochondria.42,43 A comparision of the 5 different yeast species have led to identification of a secondary structure loop in 3′-UTR with a conserved zip code. 43 This is one of the early studies of 3′ UTR function, subsequent to a previous study demonstrating ATM1 mRNA translocation to mitochondria. However, this phenomenon of mRNA transport into the mitochondria is not convincingly proven by other methods such as in vivo visualization of the RNA translocation or by other biochemical characterization.
The mitochondrial localization element for ATP2 mRNA was further narrowed down by nonhomologous random recombination, a method that enables random recombination without required homologues sequence. 44 As a result, two 50 nucleotide consensus sequence 263-312 and 164-213 downstream of the ATP2 stop codon were identified as conserved motif (Min-2). However, experiments targeting GFP to mitochondria by fusing the Min-2 element with GFP open reading frame (ORF) was unsuccessful. Such experimental failure suggests that the context of the mRNA itself might also contribute to the localization. Besides, 5′ UTR may have the ability to associate with different cofactors or ribosomes that in turn may favor cytoplasmic or mitochondrial bonding. 45
The mammalian spermatozoa was found to translate nuclear-encoded proteins by mitochondrial ribosomes. 46 Comparison of mitochondrial protein levels before and after applying with prokaryote specific translation inhibitor D-chloramphenicol 47 revealed changes in levels of proteins. Levels of several proteins such as protein kinase C (PKC), 48 angiotensin II type I receptor (AT1-R) (Gur et al. 1998), epidermal growth factor receptor (EGFR) (Lax et al. 1994), and progesterone receptor (PR) 49 were decreased in spermatozoa treated with D-chloramphenicol.
The mRNA localization to the mitochondrial surface was also performed with ATP6 gene. 50 ATP6 is mitochondrial genome coded protein. Two new constructs for ATP6 were made and transformed into the yeast which were hybridized with the mitochondria targeting sequence (MTS) of SOD2 (superoxide dismutase −2) representing 30 amino acids of ATP6 protein in frame: one with 3′ UTR of SV40 while the other hybridized with 3′ UTR of SOD2. 51 SOD2 mRNA which was found to be anchored to the mitochondrial surface through A-kinase anchor protein 121, AKAP121 by its 3′ UTR and the K homology domain of the protein.52,53 Upon transfection to HeLa cells, both constructs were translated in the cytoplasm and translocated into the mitochondria. The amount of imported ATP6 was 1.8 fold higher for the one with 3′ UTR sequence of SOD2, indicating that both pathways contribute to the protein import into the mitochondria. However, the function of the hybridized ATP6 protein is still left to be determined. Besides, overproduction of the karyopherin protein Pse1p/Kap121p improved the import of the hydrophobic protein Atm1 protein, and ABC transporter of the mitochondrial inner membrane as well as increased its transcript in cytoplasmic ribosomes that were associated with mitochondria. 39
A recent study applied the improved microarray technology and a better procedure for MLR calculation. 51 In this study, translational regulatory protein Puf3p belonging to Pumilio family (PUF) of proteins was used as a model protein to get genome wide mRNA analysis. 51 Similar to Puf6 protein, which was found to repress the translation of their target mRNAs by binding to elements located in the 3′ UTR, 54 Puf3p also binds to a consensus motif in the 3′ UTR of many mRNAs encoding mitochondrial proteins. Several Puf3p mutants, including deletion and Puf3p binding motif mutation, were generated for analysis. Comparision of the MLR expression between the wild type and Puf3p deleted yeast using microarray analysis allowed classification of the 480 MLR into 2 classes (termed as Class I and II). 244 out of 256 of Class I genes have 3′ UTR Puf3 motif and code mainly for the assembly factors of respiratory chain complexes and the mitochondrial translation machinery. Class II genes does not have not Puf3 motif and encode proteins of the respiratory chain or proteins involved in metabolic pathway. This is so far the most recent and comprehensive analysis of mRNA that get translated at mitochondrial vicinity. 55 A summary of some typical mRNA undergoing mitochondrial translocation is presented in Table 1.
Summary of experiment related to mitochondria translocation.
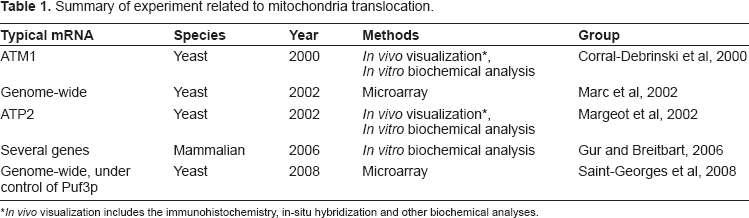
Summary of experiment related to mitochondria translocation.
In vivo visualization includes the immunohistochemistry, in-situ hybridization and other biochemical analyses.
The non-coding sequences contained within UTRs of many mRNAs serve as information for the specific localization of these mRNAs. We briefly described below a series of studies that explored general mechanisms for mRNA transportation. Although the specific mechanism for mRNAs targeting to mitochondria is not clear, it is likely that nuclear coded mitochondrial mRNAs share some common mechanism.
Passive diffusion is the simplest mean of transporting mRNA. This mode of transport also saves energy to achieve the asymmetric mRNA distribution. It does not occur only in the nucleus, but also occurs during mRNA transport in the cytoplasm. Passive diffusion results in cell polarity that are important in biological phenomenon such as oogenesis and neuronal cell development.56,57 Localized anchors in this process help to achieve translocation and translation of mRNA at a specific location. Enrichment of nanos, gcl (germ cell-less) and cyclin B mRNAs in Drosophila embryo pole plasma are examples of such mechanisms. During late oogenesis, when the cytoskeleton is no longer polarized along the anterior posterior axis, some of these mRNAs are still found enriched at the oocyte polar region. Localization of these mRNAs also depends on the earlier translation of oskar mRNA at the posterior pole and the subsequent assembly of polar granules,57–59 suggesting the existence of local anchors. Incorrectly localized mRNAs usually are degraded. 59
In the cell, active transport is a widely used mechanism to traffic organelles and other cargos.60,61 Active transport involves more elements and delivery of cargos is more precise. The mechanism consists of formation of anchoring of the particles to the cytoskeleton, translocation of the particle to its destination, and subsequent local translation of delivered mRNA molecules. 60
Molecular motors propel the transport of cargo including mRNA and this has been found to be involved in several cellular processes. Microtubules-depolymerizing drugs were shown to decrease both RNAs or mRNA binding proteins in neuronal cells.62,63 Overexpression of one of the kinesin component, conventional kinesin heavy chain (KIF5b), increased mRNA localization of distal dendrite in the cell.63,64 A well studied example of translocation along the cytoskeleton is ASH1 from budding yeast. ASH1 is a cell fate determinant. Several groups have demonstrated that ASH1 mRNA, along with at least 23 other transcripts, is localized to the budding tips of Saccharomyces cervisiae cells. 65 The expression of the ASH1 protein at budding tips is necessary for the suppression of mating type switching in the daughter cell by repression of the transcription of the HO endonuclease gene. Sequence analysis, revealed a zip-code secondary structure at 3′ UTR of the ASH1 mRNA.66,67 ASH1 mRNA is actively localized to the bud of the daughter cell. ASH1 associates with motors (myosin and actin) that transport the cargos. Two other proteins, she1 and she2 are also involved with the protein complex that forms for transport of the cargo. She2 is found to increase the affinity for the she3 C-terminus by binding to mRNA 3′ UTR zip-code. 67
Another example that highlights how nuclear acquired factors impact cytoplasmic mRNA metabolism is the localization of β-actin to the lamellar region by the zip-code binding protein (ZBP1) in several asymmetric cell types including dendrites of neurons. ZBP1 contains four KH domains and one RBD (RNA binding protein). ZBP1 binds to β-actin zip-code region, and is regulated during formation of synapses. 68 This interaction is essential for proper β-actin mRNA localization in the cytoplasm. 69 Further experiment showed that ZBP1 can block the joining of ribosomal subunits, and can be released from β-actin mRNA by Src phosporylation, which is involved in local β-actin synthesis. Mitochondrial Puf3p protein located at mitochondrial surface and associated with machinery for mitochondrial motility and inheritance, 55 was proposed to have the new function of specific localization, such as providing the conditions for a spatial-temporal regulation of mitochondrial protein import and mitochondrial protein complex assembly.54,55 Further investigations along these lines in future is expected to add more details of specific molecular mechanisms about mRNA transport near viscinity of mitochondria or into the mitochondria.
Recently, several studies have explored possible mRNA translocation associated with endoplasmic reticulum (ER). Trafficking of mRNA via ER was observed in yeast, Drosophila as well as mammalian cells using techniques like cell fractionation, reverse transcript-PCR (RT-PCR), in-situ cDNA hybridization and microarray analyses.70,71 Signal recognition particle (SRP) pathway targets ribosome-mRNA-nascent chain complexes formed in the cytosol to the ER membrane, however, mRNAs encoding cytosolic proteins also often partition with ER membranes. 72 In different studies, some mRNAs encoding cytosolic proteins such as Hsp90 have been shown to be enriched within the membrane-bound polysomes fraction. Some other mRNAs such as β-catenin and protein kinase Cβ has been found to be distributed between free and polysomes-bound fractions. Recent studies in yeast using RT-PCR in combination with fluorescence microscopy studies have shown that daughter cell localized mRNAs encoding cytosolic factors, such as Ash1 and Sro7 (a SNARE regulator and polarity factor), are highly enriched in ER-containing fractions. 73 The mechanisms to traffic mRNA to mitochondria are most likely to bind with cofactors that guide them to the direction along the cytoskeleton or ER surface. It is possible that highly abundant mRNAs are translocated to mitochondria by passive diffusion.
It has been known that majority of mitochondrial proteins are encoded by the nuclear genome and are synthesized in the cytoplasm prior to their import to mitochondria. Generally, these proteins are synthesized with mitochondrial targeting sequence (MTS) at N-terminal. Receptors such as several molecular chaperones and other factors on the mitochondrial surface recognize the precursor proteins which are then translocated across the outer membrane through the preprotein translocase of TOM complex. MTS is usually cleaved off by the mitochondrial processing peptidase (MPP) when they enter into the mitochondrial matrix. TIM23 complex is the inner mitochondrial membrane translocase that allows precursor cross through in a membrane potential-dependent manner. 74
Studies from several different groups have shown evidence that some nuclear coded mitochondrial proteins’ translation and import are correlated. Some examples of evidence of such translocation are: accumulation of mitochondria-bound polysomes when exposed to antibiotics to arrest the translation in the cytoplasm in yeast cells, failure to detect the mitochondrial precursor protein in the cytoplasm, instantaneous inhibition of import by affecting protein synthesis, location of nuclear coded mitochondrial mRNA by using the GFP as marker.14,35,38,75 A subclass of proteins known to reside in the internal mitochondrial compartments were unexpectly found in a S. cerevisiae mitochondrial outer membrane proteomic analysis. 4 The enriched precursor proteins were subjected to systematic analysis. The analysis revealed the presence of high mRNA targeting value MLR, in these proteins, which further supports that a group of proteins are likely translated next to mitochondria. Instead of transporting in the protein form, the ribosome-mRNA-nascent chain complexes are likely delivered to mitochondria and start translation associated with import machinery associated with import into mitochondria. 75 Another study of proteins with higher MLR value suggested them to be imported into inner mitochondrial surface (IMS) through TOM-TIM23 complex dependent pathway.76,77 This inference was drawn from the established Oxa1 mediated co-translational insertion model of mitochondrial coded proteins.78,79 It has been proposed that the physical contact of ribosomes with mitochondria is maintained through certain factors on mitochondrial outer membrane. 77 However, several aspects of the mechanism of co-translational import into mitochondria remain poorly understood. A model summarizing different nuclear coded mitochondrial mRNA transport pathways in the cytoplasm have been presented in Figure 2.
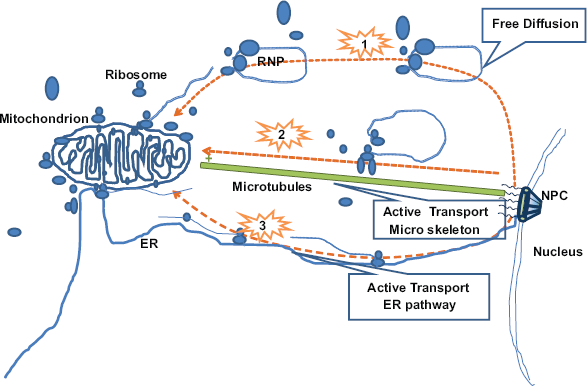
Model of different nuclear coded mitochondrial mRNA transport pathway in the cytoplasm. The possible pathways are shown as 1, 2, and 3. 1. RNP complexes are transported by free diffusion, and localized by the anchors on the mitochondrial surface. 2. RNP complexes are transported by molecular motors though the cytoskeleton such as Microtubules. 3. RNP complexes are transported by along the ER and localized at the mitochondrial surface. The mitochondrial mRNAs are probably translated at the ribosome bond to mitochondrial surface.
Generally, two transport machineries pertinent to protein/mRNA transport are thought to exist in the cytoplasm: 1) NAC transport associated with ribosomes in the cytosol, and 2) direct mRNA transport. As to mitochondrial mRNAs transport, studies in yeast of specific protein/mRNA such as ATM1 and ATP6,37,51 and genome wide analysis of mRNA targeting to mitochondria36,55 supports the direct transport of mRNA into the mitochondria in addition to protein import. However, most of these studies were more focused on the mRNP complex interactions at mitochondrial vicinity. In order to better understand the mechanism of mitochondrial mRNA transport, further investigations are necessary. Several questions remains to be answered: what group of mRNAs are delivered to the mitochondria? What are the common cofactors that interact with mRNAs? What roles they play in interacting with mRNA and other proteins of the RNP complexes? What are the post translational modifications of key proteins within the complexes? What are the interactions on the surface of mitochondria? The comprehensive genomic screen and proteomic studies are expected to provide us more details and further insights.
Mitochondria is known to use a translation machinery different than that of the cytosol, suggesting that it is unlikely that the expression of nuclear coded mRNA occurs in the mitochondria. However, it does not exclude the possibility of mRNA import into mitochondria, either as aberration or through well established mechanisms. A few studies have attempted to determine the existence of mRNA import into the mitochondria, however, availability of more direct evidence will be necessary to support or refute the idea.
As the central place for cell metabolism and their importance in cellular damage and subsequent dysfunction, mitochondria are getting more attention for a wide range of human diseases. Mitochondrial dysfunctions can lead to decreased ATP synthesis, increased endogenous ROS (Reactive Oxygen Species) production, drop in mitochondrial membrane potential (Δψm), the opening of the mitochondrial permeability transition pore (MPTP), and initiation of apoptosis. 80 Diseases associated with mitochondrial defects affect more than 1 in 8,000 individuals. 81 A substantial number of them are associated with diseases of nervous systems, for example Leber's hereditary optic neuropathy (LHON), myoclonic epilepsy, Alzheimer's disease, Parkinson's disease82–85 as well as with a variety of other systemic manifestations such as myopathy, diabetes, multiple endocrinopathy. Some of these diseases are caused by mutations in mitochondrial proteins. A large percent of these mutations are related to nuclear coded mitochondrial mRNA. The efficiency of nuclear coded mitochondrial mRNA transport to mitochondria could be an important factor for maintenance of the normal function of mitochondria. The better understanding of the mitochondrial mRNA transport mechanism is also expected to provide us insight into the diseases involving mitochondrial function and possibly new therapeutic intervention strategies.
Disclosures
The authors report no conflicts of interest.
Footnotes
Acknolwledgements
We thank Dr. Anthony Barrientos for his critical comments on the manuscript.