
Abstract
Embryonic stem cells (ESCs) are pluripotent cells with great therapeutic potentials. The in vitro differentiation of ESC was designed by recapitulating embryogenesis. Significant progress has been made to improve the in vitro differentiation protocols by toning soluble maintenance factors. However, more robust methods for lineage-specific differentiation and maturation are still under development. Considering the complexity of in vivo embryogenesis environment, extracellular matrix (ECM) cues should be considered besides growth factor cues. ECM proteins bind to cells and act as ligands of integrin receptors on cell surfaces. Here, we summarize the role of the ECM and integrins in the formation of three germ layer progenies. Various ECM–integrin interactions were found, facilitating differentiation toward definitive endoderm, hepatocyte-like cells, pancreatic beta cells, early mesodermal progenitors, cardiomyocytes, neuroectoderm lineages, and epidermal cells, such as keratinocytes and melanocytes. In the future, ECM combinations for the optimal ESC differentiation environment will require substantial study.
Introduction
Embryonic stem cells (ESCs) are derived from the inner cell mass of blastocysts.1,2 Their self-renewal and pluripo-tency to differentiate toward almost all cell types make ESC invaluable in development research, regenerative medicine, and drug screening. The differentiation of specific cell types starts from the formation of three germ layers: endoderm, mesoderm, and ectoderm. Endoderm gives rise to progenies such as liver, pancreas, and lung; mesoderm gives rise to progenies such as blood, heart, and skeletal muscle; and ectoderm gives rise to progenies such as central nervous system, hair, and skin. 3 Great efforts have been made to differentiate ESC toward derivatives of all three germ layers, such as hepatocyte-like cells, 4 pancreatic beta cells, 5 cardiomyocytes, 6 skeletal myogenic cells, 7 and motor neurons. 8 These in vitro ESC derivation strategies are a recapitulation of biochemical cues present during embryogenesis. For ESCs to reach their full potential toward specific cell types, we must improve our understanding of factors controlling cell fate. Despite a developing understanding of the role extracellular matrix (ECM) plays in controlling cell fate, 9 most current lineage-specific protocols utilize the same differentiation substrates, typically feeder cell layers or complex components containing Matrigel. The complexity of these substrates may introduce a quality control issue when differentiated products are used in clinical applications. Meanwhile, these substrates may not induce the optimal differentiation outcome, further limiting their usefulness for therapeutic applications. In response, some groups have started exploring the use of simplified ECM protein formulations or synthesized materials with known binding integrin receptors to replace feeder layers and Matrigel. The scattered information on how ECM proteins and integrin receptors are involved in ESC differentiation toward different lineages needs to be collected. In this review, we summarize the use of ECM proteins or synthesized materials in various ESC differentiation directions and evaluate the important integrin subunits involved in different lineage derivations.
Extracellular Matrix
In vivo stem cells live in a complex microenvironment called the stem cell niche. The stem cell niche influences stem cell behavior and regulates stem cell fate decisions by providing a variety of signals. These signals may be structural, physical, electrical, or biochemical. ECM is an essential component of the stem cell niche, and it is involved in almost all of these signals. 10
ECM is the physical scaffold synthesized by cells. Cells elaborate their ECM by secreting proteins, and in turn, the ECM regulates cell behavior and influences the remodeling of their matrix.11,12 This relationship between cells and ECM was proposed as dynamic reciprocity by Bornstein et al in 1982. 13 The understanding of precisely how this dynamic reciprocity regulates cell behavior is still evolving. Only recently has it become widely accepted that ECM properties may play a critical role in controlling stem cell fate. Such ECM properties include the protein composition of matrices, the availability of binding sites for specific integrin heterodimers, and physical properties like rigidity. 14 Owing to the complexity of in vivo niche, it is difficult to study all the parameters simultaneously. Efforts have been made to examine how individual matrix properties can impact stem cell fate. Engler et al15,16 demonstrated that manipulating matrix rigidity could direct lineage specification in human mesenchymal stem cells. Trappmann et al 17 went further to show that ECM cross-linking and integrin-binding site orientation and availability impact the ability of mesenchymal stem cells to sense stiffness. ESCs are more elastic and much more sensitive to local stresses induced by cell spreading.18,19 Subsequently, Engler et al's and Trappman et al's findings were also demonstrated to be true in ESC.20,21 For example, high rigid substrates were found promoting both early mesendoderm and cardiomyocyte differentiation from mouse ESCs.22,23 Besides these efforts toward ECM physical property modifications, studies on how ECM protein composition and integrin-binding site availability impact the fate of ESC are emerging. Next, we review these studies in the endoderm, mesoderm, and ectoderm sections.
Integrins
Cells interact with ECM molecules via a family of surface receptors called integrins. The type and quantity of integrins on each cell are specific to the cell and tissue type. 24 Unlike the majority of cellular receptors, integrins function primarily to facilitate the interactions between cells and ECM and transduce chemical and physical signals from the matrix. They are involved in many cellular functions, including cell cycle progression, cell adhesion, migration, and survival. 25 They are also responsible for the organization of the cytoskeleton and structural components of ECM. 25
Integrins are heterodimeric transmembrane receptors. 26 An integrin molecule is composed of two glycoprotein subunits, α and β. In vertebrates, 18 α subunits and 8 β subunits have been found, and they can form 24 different heterodimeric structures. 24 The extracellular domain of integrins binds to ECM ligands, such as laminin, fibronectin, collagen, and vitronectin, while the intracellular domain connects with cytoskeletal proteins, such as α-actinin and talin, as well as regulatory proteins, such as calreticulin and cytohesin. 27 Integrins typically interact with ligands through weak interactions, but the ligand-binding affinity may be modulated by intracellular signals. In general, the ligand-binding domain of integrins is at the ends of integrins, and it has portions on both subunits. As the ligand binds between the two subunits, the induced conformational change physically pushes the two subunits apart and initiates downstream signaling. 28 The binding between integrin receptors and ECM proteins is not specific. One ligand may interact with different integrin receptors, while one integrin may recognize different ligands. 27 For example, fibronectin interacts with at least eight integrins, and integrin αVβ5 can recognize vitronectin, osteo-pontin, and fibronectin.26,27 This transmembrane linker role makes integrins important in cell–cell/cell–ECM adhesion, signal transduction, and growth factor receptor responses.27,29
Integrins have been recognized as a key regulator in embryogenesis. 27 However, our ability to utilize integrin signaling to direct cell fate is relatively crude. What we currently know about the involvement of specific integrins in in vitro ESC differentiation is presented in Table 1.
Integrins and their ECM ligands/materials in ESC differentiation.
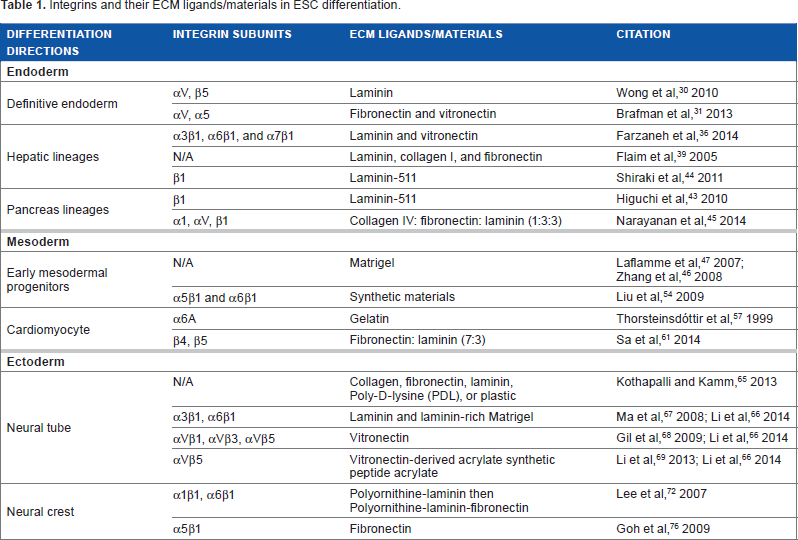
Endoderm
Endoderm is one of the three primary germ layers. Endoderm can develop into internal organs, such as pancreas, liver, and intestines. It is of great interest to differentiate ESC from endoderm and further lineages in regenerative medicine. ECM constitutions and integrin receptors mediate ESC differentiation toward both early stage definitive endoderm (DE) and advanced endoderm lineages, such as hepatocytes and pancreatic beta cells.
Wong et al 30 revealed that laminin could support the DE differentiation of human ESC (hESC) as efficient as the basement membrane (BM) matrix Matrigel. In this study, an expression profile of integrin subunits on both undifferentiated hESC and induced DE was established. In both undifferentiated and DE cells, gene expression of 11 integrin α subunits and 8 integrin β subunits was detected on different levels. The expression of three integrin subunits α3, α6, and β4 decreased in DE compared to undifferentiated cells. Meanwhile, integrins αV and β5 are highly expressed in undifferentiated hESC, and this expression is significantly increased after DE formation. Further data on the role of integrin αV in DE differentiation were also shown through a short hairpin RNA-mediated conditional knockdown of integrins αV and α5 in hESC. 31 Integrin αVβ5 has been demonstrated regulating the transforming growth factor-beta (TGF-β) signaling pathway in a number of cell types, such as myofibroblast and airway smooth muscle cells.32,33 TGF-β signaling is important for the maintenance and DE differentiation of hESC. 34 Further investigation on the correlations between TGF-β signaling and upregulation of integrins αV and β5 in the process of DE differentiation may help to achieve a higher efficiency of DE differentiation.
In the United States, end-stage liver disease (ESLD) represents the 7th leading cause of death among people between the ages of 25 and 64 as well as the 12th leading overall cause of death. 35 Liver transplantation is a lifesaving treatment for patients with ESLD, but its application is limited by the shortage of donor organs. The hESC-derived hepatocytes may offer an unlimited source for transplantations. Farzaneh et al 36 reported that human fibroblast-generated GoGel could support efficient hepatocyte-like cell differentiation from hESC. GoGel bound to α3βl, α6βl, and α7β1 integrins. 36 Among these integrins, α3β1 integrin has a high affinity for laminin, and it is important for immature and transformed hepatocyte morphogenesis.37,38 Additionally, Flaim et al 39 developed an ECM microarray that contained 32 various combinations of collagen I, collagen III, collagen IV, laminin, and fibronectin. Through this microarray, the hepatic differentiation of mouse ESC reporter cell line Ankrd17–beta-galactosidase was found the most efficient on laminin + collagen I + fibronectin. This ECM microarray platform can be a valuable tool in probing the role of the ECM and integrins in other ESC differentiation directions. Moreover, hepatocyte growth factor and TGF-β-Smad signaling mediate integrin β1 in liver development. 40 A crosstalk between integrin and growth factor signaling is indicated in this hepatic differentiation process. 40
In the treatment of type I diabetes, islet cell transplantation can restore the beta-cell mass and achieve a normal long-term glucose homeostasis. 41 ESC-derived mature pancreatic beta cells are a potential transplantation source for diabetic patients. Laminin-511 (laminin α5βlγ1 or laminin-10) is an important component of human endocrine islet BMs, and it plays a critical role in the microenvironment of pancreatic islet cells. 42 Higuchi et al 43 differentiated mouse ESCs and induced pluripotent stem cells into the DE, and then, pancreatic lineages on a synthesized BM substratum with HEK293 cell expressed laminin-511. They identified that the pancreatic differentiation signal was mediated by laminin α5 in the BM and transduced through integrin β1. As discussed in the previous section, integrin receptor β1 is a key mediator of hepatocyte differentiation; therefore, it is not surprising that this synthesized BM was also found suitable for the hepatic differentiation. 44 In addition, the expression of ECM protein collagen IV/fibronectin/laminin was estimated at 1:3:3 in rat pancreatic beta-cell line RIN5F. 45 Therefore, as expected, hESC-derived pancreatic cells expressed the highest beta-cell-specific genes, such as insulin, Glut2, and PDX1, when the ratio of collagen IV/fibronectin/laminin was 1:3:3. In this insulin-producing cell differentiation process, integrins αl, αV, and β1 were expressed as the highest among all test subunits. 45
In summary, when ESCs are directed to differentiate toward endoderm lineages, laminin substrates may be useful for improving the efficiency of differentiation toward DE. This response to laminin is mediated by integrins αV and β5. When cells are differentiated further toward pancreas or hepatic lineages, integrin β1 becomes important. In these further differentiation stages, although laminin can continue to provide necessary support, an addition of collagen, fibronectin, and/or vitronectin may offer a more optimal environment. Interestingly, the ratio of these components impacts their efficacy in directing differentiation.
Mesoderm
Mesoderm derived from ESC has potential to regenerate multiple important tissues and organs because cardiac, vascular, skeletal, and hematopoietic lineages can be derived. 3
Early mesodermal progenitors and cardiomyocytes could be induced on Matrigel.46,47 Matrigel has complex components, including a variety of growth factors and ECM proteins.48,49 Which ECM component in Matrigel supports this differentiation is not illuminated in these studies. Fibronectin and laminin are present in early stage mesoderm development of the inner cell mass. 50 Integrins α5β1 and α6β1 are important cell-interacting receptors for fibronectin and laminin, respectively,51,52 and gene mutation of integrin α5 leads to mesodermal defects in mice embryogenesis. 53 These data imply that simplified ECM, which binds to integrins α5β1 and α6β1, may be useful in mesoderm induction. Liu et al 54 designed synthetic materials containing insoluble ligands for both α5β1 and α6β1 integrins. Combined ligands supported mesoderm differentiation, whereas either ligand alone did not. 54 The activation of α5β1 modulated BMP4 expression. 55 Short-term BMP4 treatment is essential for initiating mesoderm induction; BMP signaling, together with Wnt, fibroblast growth factor (FGF), and TGF-β/nodal/activin signaling, mediates this differentiation. 46 The direct connection between BMP signaling and integrins α5β1 and α6β1 in mesoderm formation requires elucidation.
Human pluripotent stem cell-derived cardiomyocytes are a promising source for repairing a damaged heart. Integrin α6A is a splice variant form of α6. Its ligand is laminin, and it is expressed in the myocardium. 56 When mouse ESC aggregates were stimulated to differentiate toward cardiac muscles, α6A expression was upregulated. 57 Many groups have shown the use of native cardiac ECM in cardiomyocyte studies. Native cardiac ECM secreted by cardiac fibroblasts contains collagen types I and III, laminin, fibronectin, and proteoglycans. Cardiomyocytes cultured in such an ECM exhibited superior growth characteristics. 58 When native heart ECM was used to support hESC differentiation, cardiac-specific marker was expressed on a higher level in 75% native heart ECM than in 25% ECM without supplemental growth factors. 59 When native ECM was used to support mouse ESC differentiation, cardiomyocytes exhibited superior spontaneous beating characteristics in response to drug treatments and better subcellular organelle development. 60 Sa et al 61 studied the ratio of fibronectin and laminin in the native ECM. A fibronectin-to-laminin ratio of 7:3 was considered to be more optimal when compared with gelatin, on which cardiomyocytes had been efficiently developed.61,62 Integrins β4 and β5 were observed upregulated in microarray data, and their antibodies reduced the differentiation level of cardiomyocytes. 61 These two integrins promoted ESC-derived cardiomyocytes through an integrin-mediated MEK/ERK signaling pathway. 61 MEK/ERK signaling pathway has been suggested as crucial in both cardiac morphogenesis and cardiac development at various stages. 63 MEK/ERK pathway can interact with growth factors, such as FGF 4, to mediate heart development. 63 The interaction between integrin, MEK/ERK, and other signaling pathways during in vitro cardiomyocyte differentiation needs to be further addressed.
In summary, in the early stage of ESC mesoderm differentiation, the binding of ECM to integrins α5β1 and α6β1 is crucial. Matrigel and synthetic materials can both lead to necessary binding; however, new ECM combinations, such as fibronectin and laminin, may be considered as an alternative substrate option. During cardiomyocyte differentiation, fibronectin and laminin at a 7:3 ratio were simplified from native cardiac ECM composition. These ligands interacted with integrins β4 and β5 to produce superior cardiomyocyte progenies relative to gelatin, which had been the gold standard for cardiomyocyte differentiation.
Ectoderm
Ectoderm is the most distal layer of the three primary germ layers in early embryos. As ectoderm develops, it differentiates into three primary parts: the neural tube, neural crest, and external ectoderm. The neural tube and neural crest are referred to as neuroectoderm. Neuroectoderm lineages are of great clinical importance. However, it is very difficult to obtain adult stem cells from neural tissues, and there remains controversy over reports that other sources of adult stem cells can be differentiated into neuroectoderm lineages. 64 Therefore, the generation of neuroectoderm lineages from ESC becomes the primary focus of present ectoderm differentiation work.
A thorough understanding of neural tube induction and its sequential differentiation into the central nervous system provides an exciting opportunity for studying and targeting diseases, such as Alzheimer's disease and Parkinson's disease. Varied integrin–ligand interactions steer different neural cell differentiation cell fates. Collagen-coated cultures favored neural differentiation, fibronectin and laminin promoted oligodendrocyte differentiation, poly-d-lysine (PDL) induced differentiations toward both the abovementioned directions, and sole plastic surface generated the highest level of astrocytes. 65 Li et al 66 have reviewed other findings in this field: in hESC, laminin and laminin-rich Matrigel interacted with α3β1 and α6β1 to enhance neuronal generation and neurite outgrowth, 67 vitronectin–-αvβ1/αvβ3/αvβ5–-fostered oligodendrocyte, 68 and vitronectin-derived synthetic peptide-acrylate surface bounded to αvβ5-stimulated oligodendrocyte progenitor cell differentiation. 69 The abovementioned findings are in agreement with the current knowledge of embryo development: deficiency of the α3 integrin subunit has been observed in mice with defective neuron migration 70 and the interaction of laminin and fibronectin with β1 integrins was reported to be facilitating the maintenance and migration of neural precursor cells. 71 .
The differentiation of neural crest cells from peripheral migrating delaminated neuroepithelium cells is vital to the formation of the autonomic nervous system. Efficient production of differentiated ESC is critical to investigate and target treatments for peripheral nervous system diseases, such as neuralgia and Guillain-Barrέ syndrome. Polyornithine–laminin coating was used in the generation of neural crest cells from hESC-derived neural rosettes. 72 Laminin may have interacted sequentially with integrins α1β1 and/or α6β1 in this process because neural crest cells express α1β1, but Schwann cell precursors do not,73,74 and undifferentiated neural crest cells do not express appreciable levels of integrin α6β1, while fully differentiated Schwann precursor cells do. 75 Next, fibronectin was added into polyornithine–laminin coating when hESC-derived neural crest cells were further differentiated into peripheral nerve or Schwann cells. 72 This result correlates with Goh et al's findings 76 that interaction between α5βl and fibronectin is important for the survival of specialized neural crest cells. Together, a sequential combination of different ECM components (polyornithine–laminin then polyornithine–laminin-fibronectin) was designed to progressively bind a serial chain of integrins, including α1β1, α6β1, and α5β1, which are crucial in the differentiation of specific neural crest lineages. By exploiting the potential interactions of multiple integrin subunits involved in various differentiation paths, this differentiation strategy of residing cells onto a more suitable ECM as differentiation progresses is particularly inspiring.
Aside from the neuroectoderm, another clinically significant cell type that differentiates from the ectoderm is epidermal cells. Skin substitutes derived from in vitro ESC differentiation may serve as a continual source for the treatment of wound healing and skin pathological conditions, such as hypopigmentation disorders.77,78 For keratinocytes, the primary interplay between integrins and ECM includes integrin α2βl-collagen IV, α3βl-laminin 331/511, and α6β4-laminin 331/511, 79 while for melanocytes, the major interplay includes integrin α6β1-laminin and integrins α2, α5, αvβ3–collagen IV. 80 In addition, β1 integrin is responsible for melanocyte proliferation,81,82 and α2, α3, and αvβ3 integrins are integral to the formation and function of the dendritic tips, 80 which function to supply melanin to keratinocytes. 83
Primarily utilizing BMP4 and ascorbic acid, recent advances in stem cell protocols have created homogeneous populations of keratinocytes 77 and melanocytes, 78 as well as a functional pluristratified epithelium 77 from hESC. Although these protocols were initially lauded as a success in producing differentiated cells with appropriate functionality and cell surface markers, the reproductions of the differentiation by other labs suggest that keratinocytes derived from hESCs could be an incomplete or divergent form of normal squamous epithelial development.84,85 Although the discrepancies could be due to epigenetic variations among disparate hESC lines, different culture methods, and limited knowledge of the respective cell types’ development, 77 one aspect of the differentiation that has been ignored is the effect of or absence of ECM in the protocol. The utilization of integrin-ligand interactions could prove instrumental in proper epidermal differentiation and truly embody a more thorough representation of the respective epithelial ECM niche environment. As mentioned earlier, laminin and collage IV mediate crucial interactions with the epidermal integrins of keratinocytes and melanocytes in the niche environment. The use of laminin and collage IV may be exploited to produce more complete forms of properly differentiated epithelium.
Conclusions
To date, most efforts to direct ESC differentiation for research and/or clinical applications have centered around the use of soluble growth factors and small molecules based on the knowledge we have of the role of these molecules during embryogenesis. These protocols have been designed without taking into consideration the substrate or ECM. This may be the reason that the derivation of mature cell types in vitro remains a significant hurdle in ESC research. Even in the studies described above where efforts were made to optimize substrate conditions for specific purposes, no ideal mature progenies were obtained. The reason for this may be that one single ECM is not ideal for different stages of differentiation; the substrate must be optimized for each stage of maturation. The differentiation strategy described above for neural crest cells is an example of how this strategy may be used. Another example is oligodendrocytes derived from hESC, relating to the first Food and Drug Administration-approved clinical trial of hESC-derived cells, GRNOPC1 Spinal Cord Injury Trial.86,87 The cells were initially grown on Matrigel and then transferred to poly-l-lysine-coated plates for terminal differentiation.86,87 This work demonstrated that cells may need to be transferred to different ECM substrates for optimal directional differentiation as key integrins change over developmental stages. 27
Recently, the use of a three-dimensional (3D) culture system in pluripotent stem cell differentiation is emerging88,89 as a higher cell maturation level may be achieved, but the mechanism of the complex tissue self-organization needs further investigation. One possible mechanism may be that in 3D culture, ECM is dependent on the secretion of surrounding cells. As the growth and development of 3D organoid is progressive, the resulting ECM condition becomes dynamic, thus facilitating the cell maturation.
Compared to ECM proteins, synthetic ECM materials are promising in the translation of laboratory-based pluripotent stem cell research to clinical application. Clearly defined biomaterials provide repeatable lineage-specific differentiation conditions and overcome barriers such as heterogeneous rejection.90,91 In this review, the use of synthetic materials in various differentiation directions is summarized: synthesized BM substratum promotes pancreatic and hepatic differentiation,43,44 synthetic materials containing insoluble ligands for both α5β1 and α6β1 integrins support mesoderm differentiation, 54 and vitronectin-derived synthetic peptide-acrylate surface stimulates oligodendrocyte progenitor cell differentiation. 69 Advances in synthetic ECM materials are happening rapidly, evidenced by the recent development of a polymer supporting the differentiation of hESC toward each of the three germ layers. 92 Therefore, the use of the aforementioned materials may offer a broader prospect for the application of ESC differentiation.
ECM is a major component of the stem cell niche, and we are developing an understanding of how ECM and its interactions with integrin receptors can influence ESC differentiation. A more thorough understanding of integrin-mediated cell–ECM interaction and the integrin signaling pathway crosstalk in ESC differentiation may help to increase the efficiency and specificity of ESC-directed differentiation for research and clinical applications.
In this review, we summarized the use of ECM proteins in various ESC differentiation directions, in the hope of providing an optimal microenvironment for a robust, mature, and lineage-specific cell formation.
Author Contributions
Contributed to the conception and the design of the work: HW. Drafted the work: HW, XL, and JL. Revised the paper: HW. All authors reviewed and approved the final manuscript.
Footnotes
Acknowledgments
We thank Dr. Pei Wang and the members of the Wang group for advice and assistance. We also thank Travis Block for his critical comments on this article.