
Abstract
Spinal cord injury (SCI) causes functional impairment as a result of the initial injury followed by secondary injury mechanism. SCI provokes an inflammatory response that causes secondary tissue damage and neurodegeneration. While the use of neural stem cell (NSC) engraftment to mitigate secondary injury has been of interest to many researchers, it still faces several limitations. As such, we investigated if NSC-conditioned medium (NSC-M) possesses therapeutic potential for the treatment of SCI. It has been proposed that many of the beneficial effects attributed to stem cell therapies are due to secreted factors. Utilizing primary cell culture and murine models of SCI, we determined that systemic treatment with NSC-M was able to significantly improve motor function and lesion healing. In addition, NSC-M demonstrated significant anti-inflammatory potential in vitro and in vivo, reducing inflammatory cytokine expression in both activated macrophages and injured spinal cord tissues. NSC-M was also able to reduce the expression of inducible nitric oxide synthase (iNOS) within the spleen of injured animals, indicating an ability to reduce systemic inflammation. Thus, we believe that NSC-M offers a possible alternative to direct stem cell engraftment for the treatment of SCI.
Introduction
According to the National Institute of Neurological Disorders and Stroke, there are approximately 12,000 new cases of spinal cord injury (SCI) within the US annually, and more than a quarter of a million Americans are currently living with SCI 1 . The pathology of SCI can be divided into two distinct phases. First is the initial physical event, which usually involves compression and/or contusion of the spinal cord resulting in axonal damage. This is followed hours to days later by secondary injury mechanisms 2 . These secondary injuries involve persistent inflammation, glial scar formation, demyelination of surrounding neurons, and substantial cellular death3,4. Significant research has been directed toward mitigating the effects of secondary injury either by preventing further neuronal damage or by promoting neuroregeneration.
Next to surgical intervention, a common approach to treat SCI was the administration of large doses of steroids, usually methylprednisolone 5 . However, this treatment has been largely discontinued because of its time sensitivity, mild results, and high possibility of severe side effects6–9. Consequently, alternative treatment strategies are under evaluation, ranging from using antibiotics to affect microglial apoptosis to using anti-Nogo-A antibodies to promote axonal recovery10,11. However, cellular therapies, including the use of Schwann cells, embryonic stem cells (ESCs), and neural progenitors, have been of particular interest to researchers and extensively examined for their neuroregenerative potential12,13. While cellular therapies have shown promise within research settings, they still face certain limitations, including loss of differentiation potential, initiation of host immune responses, and difficulties in delivery14–16. The cytotoxic nature of the acute SCI lesion microenvironment also presents challenges to cellular engraftment 17 . Moreover, some therapies have more specific concerns, such as neural stem cell (NSC)-related induction of allodynia, pain response from a nonpainful stimuli, and ESC-specific teratoma formation and ethical concerns13,18,19.
NSCs have been shown to be highly anti-inflammatory, being able to modulate responses to cerebral inflammation and hemorrhagic stroke 20 . This ability is of interest to us considering that inflammation is a key regulator of SCI secondary injury. However, given the current limitations to clinical cell-based treatments, we instead investigated the ability of NSC-conditioned medium (NSC-M) to treat SCI. It has been postulated that stem cell-conditioned medium could be used for regenerative medicine 21 . Consequently, we wanted to determine if NSC-M could facilitate recovery within SCI. Our study found that NSC-M possesses substantial anti-inflammatory properties and can facilitate improvements to locomotion and lesion size. We believe that NSC-M offers an exciting avenue for the development of novel treatment options for those suffering from SCI.
Materials and Methods
Reagents and Antibodies
Chemical reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA). Cell culture medium was purchased from Invitrogen (Carlsbad, CA, USA) unless otherwise indicated. Basic fibroblast growth factor (bFGF; GF003-AF), epithelial growth factor (EGF), and anti-myelin basic protein (MBP; MAB386) were purchased from Millipore (Billerica, MA, USA). F4/80 and Mac-2 antibodies were produced with hybridoma cell lines (HB-198 and TIB-166) obtained from the American Tissue Culture Collection (ATCC; Manassas, VA, USA). Anti-ionized calcium-binding adapter molecule (IBA1; 019–19741) was purchased from Wako (Osaka, Japan). Anti-inducible nitric oxide synthase (iNOS; 610329) was purchased from BD Biosciences (San Jose, CA, USA). Alexa Fluor 488 anti-CD86 (105017) and Alexa Fluor 647 anti-CD206 (141711) were obtained from BioLegend (San Diego, CA, USA). Anti-glial fibrillary acidic protein (GFAP; ab53554) was purchased from Abcam (Cambridge, MA, USA). All secondary antibodies were purchased from Invitrogen.
Animals
C57BL/6J mice were purchased from Jackson Laboratory (Bar Harbor, ME, USA). Mice approximately 8–10 weeks of age (body weight of 20–28 g) were used for the study. Mice were maintained in a pathogen-free animal facility at Florida State University's College of Medicine. All animal procedures and experiments were approved by Florida State University's Institutional Animal Care and Use Committee (IACUC) and performed in accordance with the Guide for the Care and Use of Laboratory Animals.
A total of 41 mice were used for locomotive recovery studies [sham (n = 3), unconditioned control medium (CON-M; n = 19), and NSC-M (n = 19), including 3 mice per experimental group (CON-M and NSC-M) sacrificed at 15 weeks]. Seven days after injury four mice from the CON-M group and four mice from the NSC-M group were sacrificed, and spinal cord tissues were analyzed via quantitative real-time polymerase chain reaction (qPCR; n = 4). Three mice from each group (CON-M and NSC-M), at the indicated time point, were used for immunofluorescence staining. Figure 1 summarizes the experimental timeline.
Timeline for presented in vivo studies. Locomotive recovery analysis was conducted between weeks 1 and 8. Other experiments were conducted when indicated. CON-M, unconditioned control medium; NSC-M, neural stem cell-conditioned medium; BMS, Basso mouse scale; iNOS, inducible nitric oxide synthase; SCI, spinal cord injury; qPCR, quantitative real-time PCR; GFAP, glial fibrillary acidic protein.
Spinal Cord Injury Model
Female mice approximately 8–10 weeks old were used throughout these experiments. Mice were anesthetized with an intraperitoneal (IP) injection of 80 mg/kg ketamine and 10 mg/kg xylazine. Following a T9–10 laminectomy, contusive SCI was induced using an NYU impactor by dropping a 5-g rod 6.25 mm onto the spinal cord to produce a mild injury, as previously described 22 . Following skin closure, mice were placed on a warming pad and observed for cyanosis. Sterile 0.9% saline (0.5 ml) and enrofloxacin (2.5 mg/kg) were injected subcutaneously, and 0.05–0.1 mg/kg of buprenorphine was intraperitoneally injected. The mice were placed back into their cages upon awakening. One milliliter of NSC-M or CON-M was injected intraperitoneally per mouse starting 24 h after SCI every day until the mice were sacrificed. Motor function was determined using the Basso mouse scale (BMS) 23 . Sham animals were subjected to the same surgical procedure; however, injury was not induced (Fig. 1).
Tissue Collection
Following transcardial perfusion and fixation with 4% paraformaldehyde (PFA), tissue samples were collected and placed in PFA overnight at 4°C. Tissues were then transferred to a 30% sucrose solution at 4°C until tissues sank. Samples were then mounted in optimum cutting temperature (OCT) compound (Sakura Finetek USA Inc., Torrance, CA, USA), sliced with a Microm HM505E cryostat (Wayzata, MN, USA), and mounted onto chromium potassium sulfate-treated slides. Slides were stored at −80°C.
Macrophage Isolation and Culture
Mouse bone marrow-derived macrophages (BM-DMs) were prepared as previously described 24 . Briefly, bone marrow cells were harvested from 8- to 10-week-old mice. Cells were seeded at a density of 1.3 × 107 cells/ml and cultured in Dulbecco's modified Eagle's medium (DMEM; Corning, Winchester, VA, USA) supplemented with 5% newborn calf serum (NCS), 15% L929 cell-conditioned medium, and penicillin/streptomycin (Corning). After 7 days, BM-DMs were used for experimentation.
Neural Stem Cell Isolation and Culture
NSCs were derived from the cerebral cortex of embryonic mice collected on day 14 of pregnancy, as previously described 25 . Briefly, the cerebral cortex was removed and mechanically dissociated into a single-cell suspension. Cells were suspended in untreated 10-cm Petri dishes at a concentration of 1 × 105/ml in 50:50 DMEM/F12 medium supplemented with 20 ng/ml EGF, 10 ng/ml bFGF, 2% B27 supplement, and penicillin/streptomycin. Under these culture conditions, cells proliferated and aggregated into neurospheres. One week after plating primary neurospheres were collected, mechanically dissociated, and resuspended under the same culture conditions to eliminate short-term dividing precursors. NSC colonies were passaged every 7 days, at which point conditioned medium would be collected. NSCs from passages 3–10 were used for this study.
NSC-Conditioned Medium Collection
NSC-M was prepared by centrifuging collected medium at 188×g for 5 min to pellet cells, collecting the supernatant, centrifuging again at 2,300×g for 30 min to remove cell debris, and then again collecting the supernatant. Supernatants collected from NSC passages 3–10 were pooled and stored at −80°C. Unconditioned NSC medium was treated similarly and used for CON-M.
Myelin Preparation and Phagocytosis Assay
Myelin debris was isolated from the brains of 8- to 10-week-old mice via sucrose density gradient centrifugation, as described previously 26 . Briefly, brains were homogenized in ice-cold 0.32 M sucrose with a homogenizer, placed on a 0.8 M sucrose cushion, and centrifuged at 46,700×g for 35 min at 4°C. After centrifugation, myelin debris was collected from between the interface of 0.32 M and 0.85 M sucrose, and then further purified by resuspension in water and centrifugation at 7,500×g for 10 min at 4°C twice. Finally, myelin debris was dissolved with phosphate-buffered saline (PBS) to a concentration of 100 mg myelin debris per milliliter.
For phagocytosis, BM-DMs were seeded in 24-well culture plates at a density of 5 × 104 cells per well and incubated with CON-M and NSC-M for 48 h. BM-DMs were then incubated with myelin debris for 1 h. Noningested myelin debris was washed away, and the cells were lysed and collected. Phagocytosis was investigated by determining the level of phagocytosed MBP within the lysate using MBP enzyme-linked immunosorbent assay (ELISA).
Enzyme-Linked Immunosorbent Assay
To detect MBP, cell samples were collected and lysed. Lysates were mixed 1:1 with 0.5 M sodium carbonate buffer pH 9.6, added to NUNC MaxiSorp™ ELISA plates (Thermo Fisher Scientific, Waltham, MA, USA), and incubated overnight at room temperature. Samples were then blocked for 2 h at 37°C, incubated with anti-MBP (1:1,000) for 2 h at 37°C, washed, incubated with secondary antibody (1:1,000) for 1 h at room temperature, washed again, incubated with 3,3′,5,5′-tetramethylbenzidine (TMB) solution (Sigma-Aldrich) for 30 min at room temperature, and then absorbance was read at 450 nm on a PowerWave X microplate reader (Biotek Instruments Inc., Winooski, VT, USA). MBP concentration was determined by comparing absorbance to standard curves. All experiments were run in triplicate, and results were obtained from three mice per group (n = 3).
Cell Length Assay
BM-DMs were incubated with either CON-M or NSC-M for 48 h. Cells were imaged with a Nikon Ti-E microscope (Nikon Instruments, Melville, NY, USA) and analyzed with ImageJ 1.48v software [National Institutes of Health (NIH), Bethesda, MD, USA] across three independent cultures (n = 3) per condition with two technical replicates for each culture. Fifty cells per replicate were manually measured for a total of 300 cells per condition. A cell's length was stated as being the longest end-to-end 2D length.
Flow Cytometry
BM-DMs were incubated with either CON-M or NSC-M for 24 h after activation with 10 ng/ml interferon-γ (IFN-γ; Millipore) for 24 h. Single-cell suspensions were prepared in PBS containing 2% NSCs. Cells were blocked with anti-mouse CD16/32 (1 μg per 1 × 106 cells) for 20 min at 4°C and then stained with anti-mouse CD86 antibody (1 μg per 1 × 106 cells) or anti-mouse CD206 antibody (1 μg per 1 × 106 cells) for 30 min at 4°C. After three PBS washes, samples were analyzed with a BD FACSCanto flow cytometry system.
RNA Purification, cDNA Synthesis, and Quantitative Real-Time PCR
For cellular studies, BM-DMs were incubated with either CON-M or NSC-M for 12 h and then incubated with or without 20 ng/ml lipopolysaccharide (LPS) for 6 h. For tissue studies, samples were collected from animals that received daily IP injections of either NSC-M or CON-M for 7 days after injury. The samples were collected fresh and stored in liquid nitrogen. Total RNA was isolated with TRIzol (Thermo Fisher Scientific) following the manufacturer's instructions and then reverse transcribed into cDNA using the SuperScript II system (Thermo Fisher Scientific). The primers used were interleukin-1β (IL-1β) primer pair (5′-AAGTGATATTCTCCATGAGCTTTGT-3′ and 5′-TTCTTCTTTGGGTATTGCTTGG-3′); iNOS primer pair (5′-TTGGAGCGAGTTGTGGATTGT-3′ and 5′-GTAGGTGAGGGCTTGGCTGA-3′); IL-6 primer pair (5′-TGGGAAATCGTGGAAATGAG-3′ and 5′-CTCTGAAGGACTCTGGCTTTG-3′); IL-10 primer pair (5′-CAACATACTGCTAACCGACTCCT-3′ and 5′-TGAGGGTCTTCAGCTTCTCAC-3′); IL-12 primer pair (5′-CCAAACCAGCACATTGAAGA-3′ and 5′-CTACCAAGGCACAGGGTCAT-3′); and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) primer pair (5′-ATCAACGACCCCTTCATTGACC-3′ and 5′-CCAGTAGACTCCACGACATACTCAGC-3′). GAPDH level was used as the internal standard. SYBR™ green dye (Applied Biosystems, Foster City, CA, USA) was used to monitor the replication of PCR products. The results are reported as 2−AACt values.
Immunofluorescence Staining and Histology
For tissue staining, mice were transcardially perfused with 0.9% saline followed by 4% PFA. Spinal cords and spleens were removed and fixed in 4% PFA for 24 h and then cryoprotected in 30% sucrose overnight at 4°C. Tissues were then sectioned, mounted onto slides, and stored at −80°C until use. For cell culture staining, cells were grown upon coverslips, fixed with 4% PFA, and mounted onto slides after staining. All samples were blocked with 1% bovine serum albumin (BSA) in PBS supplemented with 0.3 % Triton X-100 for 1 h. Samples were then incubated with primary antibodies overnight at 4°C, washed, and then incubated with secondary antibodies at room temperature (RT) for 1 h. Finally, samples were stained with DAPI. Control samples stained only with secondary antibodies were also generated to determine the degree of nonspecific binding. Some samples were stained with Oil red O to identify accumulated lipids. Samples were imaged with a Nikon Ti-E microscope. All images were captured using identical exposure times and settings. Staining intensity quantification was performed with ImageJ 1.48v and standardized to staining controls.
Statistical Analysis
Statistical data were analyzed using SPSS 22 (StatSoft, Tulsa, OK, USA). Statistical significance between two groups was evaluated using the Student's unpaired t-test, while analysis of variance (ANOVA) was used to assess groups containing multiple comparisons. Results are reported as mean+standard error of the mean (SEM). Statistical significance was set at p < 0.05.
Results
Effect of Neural Stem Cell-Conditioned Medium Upon Macrophage Phenotype and Activation State
Our previous work showed that infiltrating macrophages are the majority cell type within lesion areas and directly contribute to secondary injury via promotion of inflammation 24 . As such, we wanted to determine how NSC-M affected macrophage behavior. Cerebral cortex NSCs were prepared from 14-day-old fetal mice, and BM-DMs were prepared from 8- to 10-week-old mice. BM-DMs were treated with either NSC-M or CON-M for 72 h. Our initial goal was to ensure that NSC-M did not alter the phenotypic traits of the macrophages. As Figure 2A shows, BM-DMs treated with either NSC-M or CON-M maintained F4/80 expression, a ubiquitous macrophage marker. One of the major activities of macrophages responding to sites of SCI is the clearance of myelin debris, which results in toxicity within the SCI lesion site if not removed 24 . Thus, we performed phagocytic assays by incubating BM-DMs cultured in the presence of either CON-M or NSC-M with myelin debris for 30 min, and then washed away noningested myelin debris. Cells were then stained with Oil red O or MBP. Figure 2B shows that macrophages stained positive with Oil red O, which stained engulfed lipids, and MBP, a major myelin protein component 24 . Moreover, ELISA results showed that BM-DMs treated with myelin debris in the presence of either standard macrophage medium, CON-M, or NSC-M contained similar levels of phagocytosed MBP (Fig. 2C). As such, our results indicate that NSC-M does not affect the phagocytic capacity of BM-DMs.

NSC-M affects M1 activation without affecting other phenotypic traits. (A) BM-DMs were incubated with either NSC-M or CON-M for 72 h and then stained for F4/80 using immunocytochemistry. Scale bars: 10 μm. (B) BM-DMs were cultured in the presence of either NSC-M or CON-M, and then incubated with brain-derived myelin debris for 30 min. Cells were then stained with either the lipophilic dye Oil red O or the anti-MBP antibody at 1:200 dilution. Scale bars: 10 μm. (C) BM-DMs pretreated with macrophage culture medium (Mϕ medium), CON-M, or NSC-M for 48 h and then incubated with myelin debris for 1 h. Intracellular MBP was assayed by ELISA to determine the quantity of phagocytosed myelin debris (n = 3). (D) BM-DMs pretreated with CON-M or NSC-M for 48 h and then stained for F4/80 (red). Scale bars: 20 μm. Cell length was quantified by measuring cells along their longest axis (n = 3). (E) BM-DMs were incubated with 10 ng/ml IFN-γ for 24 h, followed by treatment with either NSC-M or CON-M for 48 h. Cells were stained for CD86 and CD206 and then analyzed via flow cytometry (n = 3). Results are displayed as mean ± standard error (SE). ∗p < 0.05. NSC-M, neural stem cell-conditioned medium; BM-DMs, bone marrow-derived macrophages; CON-M, unconditioned control medium; MBP, myelin basic protein; IFN-γ, interferon-γ.
Our next goal was to determine if NSC-M affected the activation state of the macrophages. Figure 2D shows that NSC-M enhanced M2-like morphology, as illustrated by the increased process length of treated cells27–29. Of the two canonical macrophage types, M2 is considered to be restorative and anti-inflammatory, while M1 is proinflammatory 30 . Since NSC-M enhanced M2 morphology, we next determined if NSC-M could reduce M1 activation in response to external stimuli. M1 activation was induced by culturing BM-DMs with IFN-γ, a potent M1 activator, for 24 h, followed by NSC-M and CON-M treatment for 48 h. Flow cytometry analysis showed that NSC-M significantly reduced expression of the M1 macrophage marker CD86 in BM-DMs treated with IFN-γ when compared to CON-M, while the M2 marker, CD206, remained relatively unchanged (Fig. 2E). Our data suggest that NSC-M can mitigate macrophage M1 activation.
NSC-M Affects the Release and Expression of Inflammatory Cytokines by Macrophages
Since our initial results indicated that NSC-M reduced M1 activation in macrophages, we wanted to determine its affect on inflammatory response. We examined how NSC-M affected the transcriptional expression of IL-6, IL-10, IL-12, and iNOS. Using qPCR, we found that NSC-M was able to substantially reduce LPS-stimulated expression of proinflammatory IL-6, IL-12, and iNOS to almost basal levels (Fig. 3A–C). Moreover, treatment with NSC-M maintained the expression of the anti-inflammatory cytokine IL-10 (Fig. 3D). Taken together, these results strongly suggest that NSC-M possesses a strong anti-inflammatory ability.

qPCR analysis of various macrophage-produced cytokines. BM-DMs were incubated with either CON-M or NSC-M for 12 h and then incubated with or without 10 ng/ml LPS for 6 h. The transcriptional expression of (A) IL-6, (B) iNOS, (C) IL-10, and (D) IL-12 was then determined using qPCR. Results are displayed as mean ± standard error (SE). n = 3, ∗p < 0.05. CON-M, unconditioned control medium; NSC-M, neural stem cell-conditioned medium; IL, interleukin; iNOS, inducible nitric oxide synthase; BM-DMs, bone marrow-derived macrophages; qPCR, quantitative real-time polymerase chain reaction; LPS, lipopolysaccharide.
Effect of NSC-M on In Vivo Models of SCI
Since we had recently shown that SCIs are heavily infiltrated with BM-DMs 24 , we began our investigation determining how NSC-M modulates inflammatory cytokine expression within SCI sites. Female C57BL/6J mice (8 to 10 weeks old) were subjected to spinal cord contusion between the T9 and T10 vertebra. Twenty-four hours after injury, animals were injected with either CON-M or NSC-M intraperitoneally. After 1 week of treatment, expression levels for IL-1b, IL-6, IL-10, and IL-12 within spinal cord tissues were determined via qPCR for both injury and marginal areas (Fig. 4A). For IL-1b, IL-6, and IL-10, NSC-M substantially reduced expression when compared to CON-M (Fig. 4B–D). Interestingly, NSC-M-treated samples expressed higher levels of IL-12 over CON-M in the injury area, while CON-M samples exhibited higher expression in the marginal areas (Fig. 4E).

NSC-M treatment is anti-inflammatory in vivo. (A) General location of marginal tissues relative to injured area. Female C57BL/6J mice between 8 and 10 weeks of age were subjected to spinal cord contusion between the T9 and T10 vertebras. Twenty-four hours after injury, animals were IP injected with 1 ml of either CON-M or NSC-M, with subsequent injections every 24 h. After 1 week of treatment, marginal and injured spinal cord tissues were collected. Expression levels for (B) IL-1β, (C) IL-6, (D) IL-10, and (E) IL-12 were determined via qPCR. Results are displayed as mean + standard error (SE). n = 4, ∗p < 0.05. CON-M, unconditioned control medium; NSC-M, neural stem cell-conditioned medium; IL, interleukin; qPCR, quantitative real-time polymerase chain reaction; IP, intraperitoneal.
We next determined how NSC-M affected microglia and macrophage activity via immunohistochemistry (IHC). Injured spinal cords were examined at 1, 2, and 8 weeks after injury for iNOS and ionized calcium-binding adapter molecule 1 (IBA1). IBA1 is a potent indicator of microglia and macrophage activation within tissues 31 . While IBA1 was similarly expressed in both CON-M- and NSC-M-treated animals, NSC-M treatment reduced iNOS expression at each time point (Fig. 5A and B). It also appeared that iNOS expression within the lesion site is not related to microglia activation, considering the minimal overlap in IBA1 and iNOS staining in the marginal areas. This suggests that infiltrating BM-DMs are responsible for the detected iNOS expression, since we have previously shown that BM-DMs are the only type detected within lesion sites 24 .

In vivo iNOS expression. (A) SCI animals were treated with either NSC-M or CON-M for 8 weeks. Injured sites were examined at 1, 2, and 8 weeks after injury for iNOS and IBA1 via immunofluorescence staining of collected tissues. Scale bars: 400 μm. (B) Results were quantified by measuring iNOS+/IBA1+ signal density within the injured area with ImageJ 1.48v. Staining controls were used for background subtraction. Results are displayed as mean ± standard error (SE). n = 3, ∗p < 0.05. SCI, spinal cord injury; iNOS, inducible nitric oxide synthase; IBA1, ionized calcium-binding adapter molecule 1; CON-M, unconditioned control medium; NSC-M, neural stem cell-conditioned medium.
NSC-M Treatment Improves Motor Function Recovery Following Spinal Cord Injury
We next examined if the anti-inflammatory properties of NSC-M facilitated functional recovery following SCI. Female mice (8 to 10 weeks old) were subjected to spinal cord contusions. Twenty-four hours after injury, animals received 1 ml of either CON-M or NSC-M via IP injection. Locomotor recovery was assessed for 8 weeks using the BMS 23 . As seen in Figure 6A, NSC-M-treated animals presented better locomotion 2 weeks after injury. Additionally, NSC-M-treated animals maintained higher levels of recovery through week 8. Injury sites were also examined 15 weeks after injury via IHC. Tissues were stained with anti-GFAP, an astrocyte marker, and anti-Mac-2 (galectin-3), which we have previously shown can be used to identify infiltrating macrophages 24 . Our results indicated that NSC-M-treated animals had significantly reduced injury areas (Fig. 6B and C). Consequently, our results show that NSC-M promotes functional recovery within animals suffering from SCI.
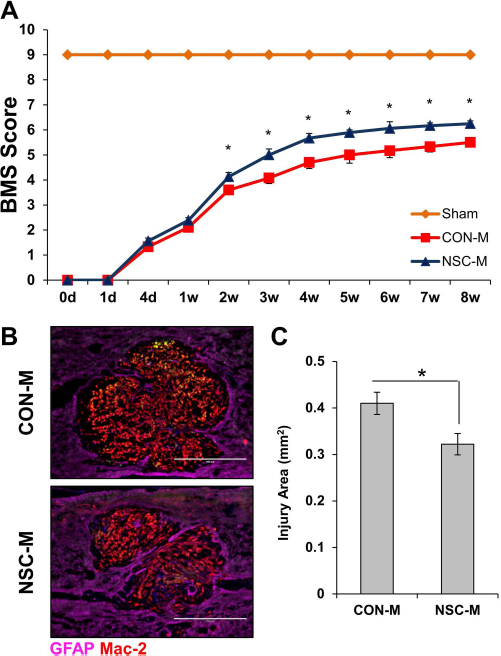
NSC-M improved motor function recovery and tissue repair. (A) Twenty-four hours after SCI, mice received daily IP injections of 1 ml of either CON-M (n = 16) or NSC-M (n = 16). Locomotor function was assessed for 8 weeks using the Basso mouse scale (BMS). (B) Injury sites at 15 weeks after injury were assessed via immunofluorescence staining for GFAP (purple) at 1:500 dilution and Mac-2 (red) at 1:500 dilution. Scale bars: 400 μm. (C) Quantification of injury site area was conducted by calculating GFAP-areas in each injury site across six sections at 100× magnification. Three mice were analyzed for each condition for a total of 18 sections per condition (n = 3). Results are displayed as mean + standard error (SE). ∗p < 0.05. CON-M, unconditioned control medium; NSC-M, neural stem cell-conditioned medium; GFAP, glial fibrillary acidic protein; IP, intraperitoneal.
Systemic Administration of NSC-M Reduced Inflammation in the Spleen
Considering that systemic administration of NSC-M was able to have pronounced effects on sites of SCI, we wanted to investigate if NSC-M affected inflammation within other areas. Since the spleen has recently been identified as a possible source of inflammatory monocytes 32 , we investigated iNOS expression within spleen macrophages following SCI. Figure 7 shows that treatment with NSC-M significantly reduced iNOS expression on macrophages 1 week after injury. It would appear that NSC-M might also reduce inflammation systemically.
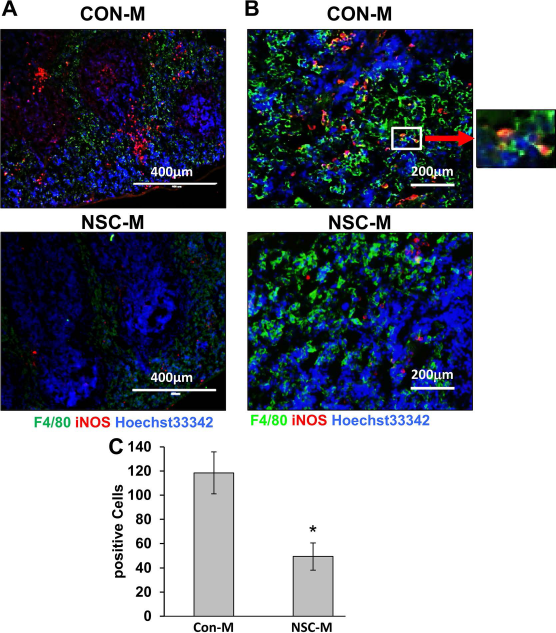
NSC-M reduced iNOS expression in the spleen of SCI-afflicted mice. (A, B) Following SCI, mice received daily injections of either NSC-M or CON-M. After 1 week, spleen tissue was collected and examined for expression of iNOS and F4/80. Scale bars: 400 μm (A), 200 μm (B). Zoomed area is 130 μm across. (C) Quantification of double iNOS+ F4/80+ cells in the spleen was conducted by counting stained cells across six sections per mouse. Results are displayed as mean ± standard error (SE). n = 3, ∗p < 0.05. CON-M, unconditioned control medium; NSC-M, neural stem cell-conditioned medium; SCI, spinal cord injury; iNOS, inducible nitric oxide synthase.
Discussion
The present study demonstrates that NSC-M possesses tremendous research potential for the treatment of SCI. One of our major findings was that NSC-M was able to facilitate locomotive recovery and reduce lesion volume. Future studies will determine if this reduction in lesion size is due to neuroprotection or neuroregeneration. Some have shown that NSCs facilitated recovery by differentiating into various cell types (e.g., neurons, oligodendrocytes, etc.) replacing those lost within the lesion site 33 . However, the extent to which NSCs differentiate and to what lineages is still under debate5,17,34. Furthermore, it has been suggested that NSCs produce their regenerative effects through release of significant quantities of neurotropic factors that stimulate axonal growth independent of differentiation 35 .
Our results also indicate that NSC-M is able to elicit anti-inflammatory effects. NSCs have been investigated for their anti-inflammatory potential in several diseases, particularly hemorrhagic stroke20,36. Inflammation is a principal regulator of the SCI secondary injury cascade and can be both beneficial and deleterious. Early on, inflammatory signals attract neutrophils, microglia, and, most importantly, circulating macrophages to the injury site37,38. Monocyte-derived cells clear toxic cellular debris and release factors that may be protective to certain elements of the damaged microenvironment39–41. However, these cells also produce high levels of inflammatory cytokines, such as IL-6, which can be destructive to surrounding cells and prevent regeneration39,42. Our results showed that NSC-M reduced inflammatory activation of BM-DMs, as evidenced by reduced CD86 expression and effects upon morphology, without affecting their ability to phagocytose myelin debris. Moreover, NSC-M was able to suppress expression of the proinflammatory cytokines IL-6, IL-12, and iNOS by macrophages in vitro and IL-1b-, IL-6–, and macrophage-specific iNOS expression in vivo. This apparent anti-inflammatory effect is in contrast to other studies, which showed that mesenchymal stem cell (MSC)-conditioned medium induces a proinflammatory response within rat macrophages 43 , even though other reports indicate that MSCs are generally anti-inflammatory44–46. These opposing results could possibly be attributed to the source of conditioned medium or model species chosen (i.e., mouse vs. rat). It should also be noted that while NSC-M treatment maintained macrophage expression of anti-inflammatory IL-10 in vitro, NSC-M suppressed IL-10 in vivo. The reason for this is still under investigation. Nevertheless, NSC-M seems to be predominantly anti-inflammatory and can reduce macrophage-related inflammatory signaling while maintaining its beneficial phagocytic ability. This could provide a means by which NSC-M promotes regeneration within injured spinal cord tissues.
Since we were able to elicit a significant anti-inflammatory response within the SCI lesion and surrounding areas, it was important to determine if it could also affect peripheral organs. Systemic inflammatory response syndrome (SIRS) can be triggered by distressing events, such as SCI47–49. SCI-induced SIRS can promote distal organ damage due to activation and infiltration of inflammatory cells from the circulation, particularly in the lungs and kidneys48,50,51. In addition, SCI causes brain inflammation associated with cognitive impairment and depressive behavior 52 . Since the spleen is a major source of immune response, it is unsurprising that this organ is also affected by traumatic events 53 . Moreover, the spleen has been indicated as being a principal source of infiltrating macrophages following SCI 54 . When we investigated the expression of the M1 marker iNOS in the spleen 1 week after SCI, we discovered a substantial number of positive cells in CON-M-treated animals, while significantly fewer were detected within NSC-M-treated animals. Our results are comparable to studies utilizing IL-33, which can be either pro- or anti-inflammatory depending on the disease 55 . Pomeshchik et al. showed that IL-33 was highly anti-inflammatory and improved functional recovery in models of SCI, while also showing that iNOS expression was markedly reduced in the spleen 56 . Considering the immunological role of the spleen, there exists the possibility that mitigation of splenic inflammatory activation may contribute to improved SCI recovery.
In conclusion, NSC-M appears to have anti-inflammatory potential, and further refinement could offer an alternative to stem cell engraftment. While the use of stem cell-conditioned medium directly could alleviate stem cell production difficulties and concerns over cell death due to the toxic nature of the SCI lesion 57 , the use of such a heterogeneous mixture could present other clinical difficulties, such as issues with standardization. Moreover, the source of the NSCs, that is, patient derived or allogamous, would need to be considered in order to avoid unintended immune response to foreign material. As such, we believe that future endeavors should focus on identifying specific factors within NSC-M that induce beneficial effects and on translating them into clinical usage. Regardless, our results indicate that NSC-M offers an exciting opportunity for SCI research.
Footnotes
Acknowledgment
This work was supported by the National Institutes of Health (R01GM100474-4 and R01G M07261-11), the National Natural Science Foundation of China (31571408 and 81571395), and the China Scholarship Council (CSC201306280104). The authors have made the following declarations about their contributions: Y.R. conceived and designed the experiments; Z.C., D.B.B., L.S., W.T., and R.D. performed the experiments; Z.C., D.B.B., J.F., X.H., and Y.R. analyzed the data; X.C., Y.X., J.L., J.F., and X.H. contributed reagents/materials/analysis tools; and D.B.B. and Y.R. contributed to the writing of the manuscript. The authors declare no conflicts of interest.