
Abstract
Ionizing radiation is effective to treat malignant pelvic cancers, but the toxicity to surrounding healthy tissue remains a substantial limitation. Early and late side effects not only limit the escalation of the radiation dose to the tumor but may also be life-threatening in some patients. Numerous preclinical studies determined specific mechanisms induced after irradiation in different compartments of the intestine. This review outlines the complexity of the pathogenesis, highlighting the roles of the epithelial barrier in the vascular network, and the inflammatory microenvironment, which together lead to chronic fibrosis. Despite the large number of pharmacological molecules available, the studies presented in this review provide encouraging proof of concept regarding the use of mesenchymal stromal cell (MSC) therapy to treat radiation-induced intestinal damage. The therapeutic efficacy of MSCs has been demonstrated in animal models and in patients, but an enormous number of cells and multiple injections are needed due to their poor engraftment capacity. Moreover, it has been observed that although MSCs have pleiotropic effects, some intestinal compartments are less restored after a high dose of irradiation. Future research should seek to optimize the efficacy of the injected cells, particularly with regard to extending their life span in the irradiated tissue. Moreover, improving the host microenvironment, combining MSCs with other specific regenerative cells, or introducing new tissue engineering strategies could be tested as methods to treat the severe side effects of pelvic radiotherapy.
Introduction
Radiotherapy is a mainstay in the management of malignant pelvic diseases. However, exposure of healthy tissue within the radiation field to ionizing radiation (IR) remains inevitable and leads to acute and/or chronic symptoms, which have been described as a new pathology by Andreyev et al., namely, “pelvic radiation disease” (4). At present, the number of declared pelvic cancers is increasing, and the advances in the efficacy of tumor control have led to an increase in the number of cancer survivors suffering from treatment-related adverse effects. Paradoxically, modern therapies threaten to increase the burden of chronic toxicity, not reduce it. The gastrointestinal symptoms induced by the chronic toxicity of irradiation have a substantial impact on the patient's quality of life. It is therefore important to understand these complications. Acute reactions impacting normal tissue injury early after irradiation are related to oxidative stress, modification of gene expression, and inflammation altering cells of the microenvironment as well as stem cells. These changes prime the irradiated tissue for a wide range of multifaceted late effects. Moreover, irradiation of the gut induces specific processes due to the presence of microbiota. This review outlines the pathophysiology of intestinal radiation injury and describes the involvement of epithelial, vascular, inflammatory, and mesenchymal compartments, which may result in rising or even catastrophic complications, such as obstruction and perforation of the intestine. Numerous pharmacological molecules have been studied, and stem cell therapy using pleiotropic mesenchymal stromal cells (MSCs) has provided encouraging results for treating the adverse effects of radiotherapy. Moreover, research is being carried out to use MSCs associated with innovative tissue engineering methods to improve their therapeutic efficacy.
Intestinal Epithelium
The intestinal tract is composed of the small intestine and the colon. The epithelium of the small intestine consists of repeated finger-like invaginations called crypts of Lieberkühn [residence of intestinal stem cells (ISCs)] associated with numerous protrusions/villi, which maximize nutrient breakdown and absorption. Villi are absent from the colon, and crypts are smaller compared with those found in the small intestine. The main function of the colon is the reabsorption of water from feces. Beside their own functions, the small and large intestinal epithelia act as a barrier against the penetration of pathogens. The intestinal epithelium is the most rapidly self-renewing tissue in the human body, with a 3- to 5-day turnover rate (74) throughout life. This process is regulated by ISCs residing near the bottom of the epithelial crypts and maintained by specific signals from the surrounding niche. In the small intestine, there are around four to six ISCs per crypt interspaced with the differentiated Paneth cells. Paneth cells belong to the secretory lineage. They secrete antibacterial components, but above all, they create a home for stem cells by providing signals [e.g., epidermal growth factor (EGF), Notch, and Wnt ligands] important for stem cell maintenance (Fig. 1). Paneth cells are absent from the colon. However, deep crypt secretory cells may represent the colon counterparts of Paneth cells (136).

Schematic illustrations of colonic crypt in normal conditions (left) or after receiving an increasing dose of irradiation (middle and right). In normal condition, two putative populations of stem cells exist: the CBCs located in the deep crypt (green cells) and the quiescent/reserve population (+4 stem cell) located above (blue cells). The crypt unit is surrounded by pericryptal cells and mesenchymal cells including myofibroblasts. The confined microenvironment is known as the stem cell niche that generates signals involved in stem cell maintenance and progeny differentiation according to a gradient of molecules. Stem cells give rise to transit-amplifying cells (red cells) that differentiate into enterocytes, enteroendocrine, and goblet cells. According to the irradiation dose, +4 stem cell can acquire Lgr5+ and proliferative phenotype (middle). Moreover, modification of the concentration gradient of molecules secreted by the niche, by modulation of gene expression or cell disruption following intestinal damage, can participate to modify the stem cell function. In this case, the number of proliferative cells increases. The resultant changes can have consequences on tumorigenesis, fibrosis, growth, and the regenerative capacity of the epithelium. At a dose of irradiation that induces apoptosis of all intestinal stem cells (ISCs), the epithelium cannot be renewed (right).
Two models of ISC identity have been proposed: the label-retaining cells (LRC)+4 model and the crypt base columnar (CBC) cell model. The LRC+4 model was proposed by Potten (133). Recent studies have focused on the identification of markers of cells located at the +4 position [B-lymphoma moloney murine leukemia virus insertion region-1 (Bmi1), telomerase reverse transcriptase (Tert), homeodomain-only protein homeobox (Hopx), and leucinerich and immunoglobulin-like domains protein 1 (Lrig1)] and defined these cells as slowly cycling/quiescent cells. Bmi1 was the first stem cell marker investigated by lineage tracing and was reported to mark rare, slowly cycling cells at the +4 cell position, uniquely in the proximal small intestine. The CBC model revealed the existence of immature, cycling cells wedged between Paneth cells at positions 1–4 (32). Determined by a microarray experiment on human colonic cell line, Lgr5 was revealed to be a good CBC marker (9,172). It has been demonstrated that isolation of single Lgr5+ cells in an adequate culture condition gives rise to a self-organizing mini-gut comprising all the differentiated intestinal cell types (145).
ISCs give rise to progenitor cells named transit-amplifying cells, which rapidly divide and differentiate into six subtypes of cells: enterocytes, neuroendocrine, goblet, Paneth, Tuft, and M cells. The two latter are rarely forming descendants of Lgr5+ stem cells. Proliferating progenitors migrate upward while they differentiate into nutrient-absorbing enterocytes and secretory cells producing mucins (goblet cells) or hormones (enteroendocrine cells). The intestinal epithelial barrier consists of enterocytes tightly connected via intercellular junctions. Enterocytes also secrete cytokines and chemokines, which trigger the inflammatory response, as a second line of defense against luminal contents. In addition, goblet cells secrete the mucus layer that protects the mucosal surface from antigens and helps maintain the intestinal barrier function. Tuft cells are a secretory cell lineage. They secrete prostaglandin precursors and possibly perform a sensory role for luminal contents. M cells reside in the specialized epithelium overlying Peyer's patches. They capture and transport antigens to immune cells in the patches. To maintain the integrity of the intestinal epithelium, ISCs must self-renew, proliferate, and differentiate to populate the epithelium. The regulation of ISCs occurs within a discrete microenvironment confined to the crypt base, known as the stem cell niche. The stem cell niche is a complex and dynamic milieu that adapts in response to environmental stress. This niche is made of and influenced by nearby proliferating and differentiating epithelial cells as well as by surrounding mesenchymal cells. Although epithelial cells and mesenchymal cells are separated by the basement membrane, epithelial–mesenchymal cross talk is required to maintain the stem cell niche. The microenvironment surrounding the stem cell zone consists of enteric neurons, endothelial cells (ECs), smooth muscle cells (SMCs), intraepithelial lymphocytes, macrophages, and fibroblasts/myofibroblasts together with the extracellular matrix (ECM). The niche generates signals that favor the existence of a stem cell in its undifferentiated state and also provides an optimal microenvironment for the production of differentiated progeny. The paracrine secretion of molecules by the niche component acts according to a gradient to their target cells. The main signaling pathways involved in niche maintenance are Wnt, bone morphogenetic protein (BMP), Hedgehog, and Notch (18) (Fig. 1).
Pathophysiology of Intestinal Radiation Damage
The exposure of the abdominopelvic region to IR, such as that received during radiotherapy, is associated with the development of a number of symptoms that may limit the course of the cancer therapy. The clinical expression of the gastrointestinal radiation toxicity results from multiple and sequential organ dysfunctions, which have been recently recognized as a new pathology called “pelvic radiation disease” by Andreyev et al. in 2010 (4). Intestinal toxicity is a multifactorial problem, related not only to the dose and method of radiation delivered but also to the intrinsic processes within the responding tissues as regard cellular injury (individual genetic background and comorbid factors such as obesity, smoking, diabetes, vascular disorders, and inflammatory bowel disease), although their exact contributions are unknown.
Tissue response to radiation exposure is considered a continuum between very acute and chronic forms. Acute radiation damages, often occurring within 3 months after radiotherapy, are characterized by diarrhea, incontinency, bleeding, and abdominal pain (6). Acute radiation symptoms generally improve over time, but in 10% of cases, the chronic symptoms associated with the cancer treatment persist. Chronic radiation damages appear between 18 months and 6 years after radiotherapy. Chronic dysfunction may follow without pause from the acute symptoms or may arise de novo months, years, or even decades later (56). Chronic damage after radiation exposure is associated with an aggravation of the acute symptoms (in frequency and importance) (193) associated with the formation of fistulas, local abscesses, perforations, and, in severe cases, occlusion that can lead to mortality (56).
The pathogenesis of intestinal radiation damage is complex. Generally, it is ascribed to the effects of IR on the cell cycle of ISCs. However, the increasing knowledge of radiation biology evidences that all cell types are sensitive/modified by IR, in particular the stromal compartment, ECs, and SMCs (129). The enteric nervous system (ENS), composed of an extensive and elaborate network of neurons involved in the physiological functions of the gastrointestinal tract, such as motility and mucus secretion, has also been demonstrated to play a critical role in intestinal radiation injuries (179). The tissue response to radiation exposure involves tissue necrosis and ischemia, chronic and self-maintained scarring process with excessive collagen deposition. These processes induce fibrosis and, later, organ dysfunction leading ultimately to occlusion.
Inflammatory Reaction, Oxidative Stress, and Cell Injury
When tissue is exposed to IR, radiochemical cell damage can occur by either direct or indirect action. The direct effect of IR consists of the particles/ions physically breaking the macromolecules. The indirect effect of IR consists of the production of highly reactive free radicals from the radiolysis of water and in the early activation of nitric oxide synthase (NOS) enzymes. These early biochemical modifications are responsible for the most important effects in the cells; however, it has been demonstrated that oxidative stress endures late after the initial exposure. Indeed, the oxidative stress is transmitted to the daughter cells and spreads to nonirradiated cells by intercellular mechanisms (8). Among the multitude effects, IR also disrupts the mitochondrial functions. Under normal conditions, the mitochondria participate in the regulation of the physiological reactive oxygen species (ROS) that are involved in the essential signaling pathways involved in numerous cellular functions, such as cellular differentiation and proliferation. Thus, mitochondrial dysfunction significantly contributes to the disturbance of the oxidation–reduction reactions that determine the cellular redox environment. Moreover, modifications of the antioxidant defense system have been demonstrated after intestinal irradiation (71). In response to IR, the immune cell recruitment (polymorphonuclear cells and macrophages) participates in the maintenance of the oxidative stress in tissue.
Reactive oxygen and nitrogen species (ROS/RNS) produced after IR are able to diffuse and to interact with biological targets causing damage to DNA, proteins, and lipids. The direct and indirect effects of IR initiate a series of biochemical and molecular signaling events that may repair the damage or induce cell phenotypic modifications. It is now well known that radiation induces DNA lesions, including damage to nucleotide bases, as well as DNA single- and double-strand breaks. DNA strand breaks result in the activation of multiple transduction pathways that activate transcription factors such as p53 and nuclear factor κB (NF-κB) (36). Moreover, lipid peroxidation results in the activation of genes such as those encoding JNK and C-Jun, which could upregulate other factors, such as NRF2. The transcription factor activation results in the upregulation of an extensive list of target genes, among which are proinflammatory cytokines [tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), and IL-6], chemokines [IL-8 and monocyte chemoattractant protein-1 (MCP-1)], cell adhesion molecules (integrins and selectins), stress response genes, and cell surface receptors (157,158). A consequence of the gene upregulation is that a wide range of biologically active proteins accumulate and target the submucosa; they not only induce tissue injury but also provide a positive feedback to amplify the initial damage caused by radiation.
Depending on the importance of the damage induced by IR, cell death by apoptosis or necrosis or mitotic catastrophe can be observed. Cycling ISCs have been described as highly radiosensitive, a property that is positive for stem cells, preventing the accumulation of deleterious genome changes (133), although resulting in intestinal barrier disruption and impaired mucosal regeneration. Radiosensitivity has been used in numerous studies to analyze adult ISC biology and, particularly, the regenerative process.
The intestinal epithelium has a considerable ability to survive and regenerate following injury. Irradiation-induced injury models have contributed to a better understanding of the mechanisms of the regeneration process, revealing the existence, in the lower regions of the crypt, of “regular” stem cells (Lgr5+ CBC) involved in the day-to-day renewal of the epithelium and “reserve” stem cells (LRC+4) mobilized after epithelial injuries. Some early progenitors can be recalled to the stem cell pool after irradiation (23,114,167,190) (Fig. 1). The fact that distinct or overlapping ISC populations with stem cell potential exist has been a matter of intense debate. Recent data evidenced for the high plasticity of cells within the epithelium, demonstrating that intestinal crypts are dynamic structures composed of diverse cell populations, including transit-amplifying progenitors, which carry some potential to assume ISC function when loss of a vital pool threatens the mucosa (Fig. 1). Such plasticity within the crypts suggests an important role of a niche-induced conversion of progenitors to functional stem cells. However, depending on the dose of irradiation, when CBC and reserve stem cells are both depleted, severe crypt loss ensues, leading to gastrointestinal failure.
It was long believed that the stem/progenitor radiosensitive epithelial cell compartment was a determinant in intestinal radiation toxicity and that tissue regeneration depended on the survival of stem/progenitor cells. Indeed, injection of R-spondin, a ligand of the Lgr5 receptor, before irradiation protects mice against the gastrointestinal syndrome (16). However, the increasing knowledge of radiation biology demonstrates that various cell types are modified extremely rapidly after irradiation and may influence the chronicity of the late damages. During the very early stages after irradiation, while the intestinal mucosa seems to have a normal structure, numerous studies describe a cascade of events beginning in the mucosa and the adjacent compartments that participate in the destruction of the mucosa. Even if endothelial apoptosis as the initiator event of intestinal radio-induced damage is currently under debate (129,148), it is admitted that EC apoptosis participates in the tissue damage induced after irradiation. Indeed, in irradiated plasminogen activator inhibitor-1 (PAI-1)–/– mice, a decrease in the intestinal damage associated with reduced endothelial apoptosis has been observed (1). Moreover, endothelial-specific PAI-1 deletion affects the radiation-induced intestinal toxicity, thus demonstrating the endothelium-dependent mechanism of intestinal injury (138). After irradiation, the vascular endothelium acquires a proinflammatory, prothrombotic, and antifibrinolytic phenotype (Fig. 2). The phenotype induced after irradiation in ECs is characterized by the increased expression of cell adhesion proteins [vascular cell adhesion molecule 1 (VCAM-1), intracellular adhesion molecule 1 (ICAM-1), platelet and endothelial cell adhesion molecule 1 (PECAM-1), and E- and P-selectins] favoring bloodstream leukocyte transmigration and increase in platelet/EC interaction. Moreover, EC exposure led to the expression of several proinflammatory mediators such as IL-6 and IL-8 (61,112,117). These studies demonstrated that adhesion molecules, proinflammatory molecules, and chemoattractant molecules expressed on ECs after irradiation play a crucial role in the recruitment of leukocytes and in the inflammatory process, having consequences on the regenerative process. ECs play a key role for vascular tone by secretion of vasoconstrictor and vasodilator molecules that act on SMCs. During early events following irradiation, the vascular tone is modified. In particular, it has been demonstrated that irradiation disturbs the bioavailability of nitric oxide (NO) because of the effects of ROS on the endothelium (72). Another major dysfunction of the ECs is their loss of thromboresistance characterized by aggregation of platelets and development of platelet-fibrin clots associated with adhesion of leukocytes (178). The underlying molecular mechanisms involved the increase of various factors, such as the von Willebrand factor, platelet-activating factor, tissue factor, and the decrease in the anticoagulant thrombomodulin. Downregulation of thrombomodulin stimulates fibrogenesis and decreases fibrinolysis. Platelet aggregation, observed after irradiation, elicits an inflammatory response by the production of free radicals and the release of inflammatory molecules such as the potent profibrosing mediator transforming growth factor-β (TGF-β). Indeed, platelets contain a high amount of TGF-β (119).

Schematic representation of the intestinal alterations induced after irradiation. This view highlights the complexity of radio-induced pathology with the interconnections of events: modification of the epithelial cell compartment, activation of the vascular compartment, and initiation and perpetuation of the inflammatory process, leading ultimately to fibrogenesis.
Disruption of the Intestinal Epithelial Barrier
In normal conditions, the intestinal epithelial barrier is not impermeable, permitting fluid transport (108) and communication between the mucosal immune system and the commensal flora. The intestinal epithelial barrier is maintained by intracellular junctional complexes, such as tight junctions (TJs), adherent junctions, and desmosomes. TJs are composed of transmembrane proteins, claudins, occludin, and junctional adhesion molecules (JAMs) interacting with cytoplasmic proteins, such as zonula occludens-1 (ZO-1), to maintain dynamic structures with the cell cytoskeleton, thus determining paracellular spaces.
Disruption of the epithelial barrier after irradiation is related to the received dose. High doses of irradiation lead to complete loss of ISCs, thus entailing crypt loss. At lower doses, there is no sterilization of the crypts, but structural disruption occurs, which has been associated with increased intestinal permeability (IP). An increased passage of tracer in the extracellular spaces was observed in rat ileum after 5-Gy irradiation (132,156). Perturbations in the mucosal barrier function have been reported in breast cancer, sarcoma, and colorectal cancer patients treated with different chemotherapy agents or after radiotherapy (37,44,48,118,130).
Numerous studies have demonstrated a decrease in the TJ complexes that may disturb the epithelial barrier and increase the IP. Studies have shown that IR increases IP through a combination of epithelial TJ disruption and insufficient replacement of villus epithelium (42,118,155). Among the various molecular components of TJs, claudins play a critical role in tightening cell–cell contacts. Alteration of claudins is closely associated with inflammatory pathologies as well as radiation-induced intestinal injury (90,107,153,185,196). Studies have demonstrated that claudin-3 and claudin-4 decreased, whereas claudin-2 expression increased after irradiation (124,153). Moreover, epithelial barrier dysfunction allows infiltration and proliferation of bacteria in the mucosa (Fig. 2). This bacterial invasion induces the infiltrating mononuclear cells (MNCs) to release proinflammatory cytokines and ROS (45). A correlation between increased IP and inflammatory response has been described in patients with ovarian carcinoma (113), suggesting a role of the proinflammatory cytokines secreted after irradiation, such as IL-1β and TNF-α, in the increased IP. The interaction between the epithelium and inflammation is a two-way process. Intestinal epithelial cell treatment with TNF-α increases the expression and activation of the myosin light-chain kinase (MLCK), associated with an increase in paracellular permeability (104,177). Once activated, the MLCK phosphorylates MLC, leading to a contraction of the actin cytoskeleton and the opening of the TJs. In contrast, interferon-γ (IFN-γ) has a direct effect on junction proteins inducing endocytosis of occludin, JAM-1, and claudin-1 through macropinocytosis (82,170). Furthermore, in an animal model with constitutively increased IP, an inflammatory process was observed, suggesting that dysfunction of the epithelial barrier results in the recruitment or activation of immune cells (161).
Microbiota and Radiation-Induced Intestinal Injuries
The human gastrointestinal tract contains 1–2 kg of bacteria living in symbiosis with the host. The microbes obtain nutrients from the human host and protection from competing species by the immune system. However, the microbiota also benefits the host through different interactions by playing a role in the development and homeostasis of the intestinal epithelium, instructing the immune system and exerting an antioxidant activity. For example, germ-free animals display impaired epithelial proliferation in their crypts and have rudimentary mucosal-associated lymphoid tissues (173). The host distinguishes signals from microorganisms through transmembrane receptors. The well-known receptors are the toll-like receptor (TLR) family and the nucleotide-binding oligomerization domain receptor family (52,137). Bacteria are recognized by the TLRs, present at the external membrane of epithelial and immune cells, leading to the activation of NF-κB (41,137). Consecutively, activation of NF-κB ensures the development of the innate response but also participates in epithelial wound repair (48,73). The deleterious effect of IR on the epithelial barrier has been thought to be exacerbated by penetration of the gut microbiota into the tissue. The role of microbiota in initially driving or perpetuation of intestinal inflammation after irradiation has been highlighted by the use of antibiotics that generally reduces radiation-induced mortality in animal models. However, the efficacy of antibiotic treatment varies depending on the model used (22,88). Various studies demonstrated that germ-free mice show better survival than mice with conventional microflora at an equivalent radiation dose (35,111,127). The authors demonstrated that a protein called fasting-induced adipose factor (FIAF) decreases in germ-free mice, leading to an increase in endothelial and lymphocyte apoptosis. This study highlights the importance of microbial regulation not only on the epithelial but also on the endothelial compartment to modulate intestinal radiation toxicity. However, because germ-free animals are known to have an impaired epithelial turnover (168), the involvement of microbiota in intestinal radioprotection must be reevaluated. Other studies demonstrated that pretreatment with bacterial component, TLR4 or TLR5 agonists, protects gut mucosal tissue against radiation-induced damage (24,140). In the same way, the use of a probiotic, through the TLR2 pathway, improves the severity of the intestinal damage after irradiation (34). Clinically, a phase II study demonstrated the efficacy of a nutritional supplement based on probiotics in preventing radiation-related gastrointestinal disorders (146).
Dysbiosis is the global imbalance of bacterial populations in the gut in various disorders such as radiation bowel disease. The gut microflora can have various radiation sensitivities. Moreover, modifications of the microenvironment after IR can influence microbial colonization. Various studies identified quantitative changes of specific groups of gut microbiota after pelvic radiotherapy (85,106,176). Although these studies are difficult to interpret due to the difference of microbiota between healthy individuals and cancer patients treated with chemotherapy or radiochemotherapy, it seems that radiotherapy decreases the microbial heterogeneity. Husebye et al. demonstrated that late radiation enteropathy leads to Gram-negative bacilli overgrowth and impaired intestinal motility (80). Dysbiosis also causes alteration of the expression and distribution of intercellular TJs (103), leading to the penetration of antigens participating in the chronicity of intestinal inflammation (7). Moreover, a pilot study investigated the correlation between fatigue, diarrhea, and microbial dysbiosis induced after pelvic cancer therapy (176). The epithelial mucus layer is a protective factor that contributes to intestinal integrity and is also regulated by bacteria. Moreover, bacteria induce secretion of numerous anti-inflammatory molecules such as short fatty acid, secondary bile acid, and butyrate, and modification of the microbial profile can exacerbate the intestinal inflammation. Altogether, these studies demonstrated that microbiota not only is involved in challenging the intestinal immune system after epithelial barrier disruption but could act at different levels to participate in intestinal radiation toxicity. However, TLR signaling pathways have an important role in the epithelial regenerative process and could be an interesting therapeutic target for protection against radiation injury.
Inflammation and Immunity in Radiation-Induced Damage
The excessive production of reactive free radicals generated immediately after IR is considered as the first proinflammatory signal. Subsequently, the radio-induced inflammatory process extends to innate and adaptive immune responses contributing significantly to the radiation injuries (53) (Fig. 2). Moreover, there is now an incontrovertible evidence of interaction between the immune system, in particular mast cells (MCs), and the ENS in the genesis and development of structural and functional damages induced after abdominal irradiation. Although the inflammatory process is necessary for normal scarring, tissue response to irradiation is considered as a chronic and self-maintaining scarring process leading to fibrosis.
Production of reactive free radicals activates cell death and proinflammatory molecule signaling pathways in irradiated microvascular and mucosal compartments. By targeting the vascular endothelium, irradiation induces, in a short time, modification of the phenotype, resulting in the migration of leukocytes into the damaged intestine for clearing the tissue. The epithelial cell death induced by irradiation leads to impaired barrier function and pathogen penetration into the mucosa. Neutrophils, possessing phagocytic and antimicrobial capacities, are the first cells that migrate to the injury sites. They eliminate dead cell debris to facilitate tissue repair; however, they also secrete high levels of antipathogenic enzymes, such as myeloperoxidase, which generate oxidative stress. This process adds to the oxidative stress induced by the radiolysis of water and reduces the regenerative capacities of the tissue. In rodent models, localized single-dose or fractionated radiation leads to an influx of neutrophils 1 week after irradiation and lasts up to 26 weeks (54,66). This influx is associated with increased expression of molecules having strong neutrophil chemoattractant activity after colorectal irradiation (14,20). Human biopsies from the rectal mucosa of patients at 2 and 6 weeks during the course of ongoing radiotherapy for prostate carcinoma showed an increased number of neutrophils (77). These observations lead us to consider neutrophils as deleterious in the radio-induced inflammatory process. However, in a model of radiation proctitis, in MC-deficient mice, Blirando et al. reported an acute increase in the influx of neutrophils associated with less colorectal damage (19). Another study also reported the importance of neutrophils influx in the radiation healing process after skin irradiation (51). Therefore, the detrimental or useful properties of early neutrophil infiltration after irradiation need to be elucidated.
The inflammatory microenvironment present in the irradiated tissue can induce the recruitment of circulating monocytes as well as the activation of resident macrophages. Depending on the inflammatory microenvironment, monocytes can differentiate into type 1 (M1/proinflammatory) or type 2 macrophages (M2/anti-inflammatory). After irradiation, numerous studies reported mucosal macrophage invasion in human tissue as well as in animal models (14,20,77). However, further investigations are needed to characterize the subtype of the macrophages induced after irradiation. Macrophages also play a key role in the resolution of inflammation by clearing the tissue and secreting feedback signals that switch off the inflammatory response. Moreover, the epithelial breakthrough induced after intestinal irradiation facilitates the entry of pathogens, leading to macrophage activation that could secrete proinflammatory cytokines detected in mucosa such as IL-1β, IL-8, and IL-6 (14,20).
In humans, lymphocyte infiltration is a common feature after radiotherapy in the subacute and chronic phases (147), but it is difficult to assess their functional significance. Recently, in a rat model of colorectal irradiation, the depletion of T cells induced a reduction of mucosal injury and colonic muscle contractibility (14). In the same model, an infiltration of CD4 and CD8 T cells in colonic mucosa was shown as soon as 7 days after irradiation. Although the percentage of CD8 T cells returned to the control level 3 weeks after irradiation, CD4 T cells persisted in the colonic mucosa for 2 months. Following antigenic stimulation, naive CD4 T cells are able to differentiate into different effector T-cell subsets (TH1/TH2/TH17/Treg) with specific features such as cytokine production and functional properties. A TH1/TH2 immune imbalance has been shown to play a determining role in many inflammatory diseases and to potentially be involved in the chronicity of radiation-induced lesions. It has been demonstrated that fractionated irradiation localized to the colorectum induces expression of the TH2-specific transcriptional factor GATA3 for 6 months postirradiation (66). Moreover, intestinal irradiation in a TH1-deficient environment accelerates fibrogenesis (98). In an abdominal irradiation model, an increase in the Treg population, with impaired ability to control effector T cells, has been reported (14,17). The lack of capacity of Tregs to induce IL-10 secretion after irradiation can contribute to uncover the immune tolerance in the intestine. Until recently, the TH17 population had not been studied, and it has now been demonstrated that colorectal irradiation in rats specifically induces CD4 TH17 cells (13). Targeting the pathways of TH17 differentiation after irradiation can therefore be of therapeutic interest.
MCs have also been associated with diseases characterized by pathological tissue remodeling and fibrosis in different organs including intestinal radiation fibrosis. MCs secrete a plethora of inflammatory, vasoactive, chemotactic, and fibrotic molecules that interact with the immune system and the mucosal microenvironment including the ENS. In human proctitis following radiotherapy, MC hyperplasia has been highlighted in areas of inflammation and collagen deposition (20). Various studies, using MC-depleted animals, demonstrated their importance in late tissue fibrosis after intestinal irradiation even if the role of MCs in acute radiation damage is still controversial (20,197). The significance of various MC mediators in radiation fibrosis is not completely understood. However, in vitro studies demonstrated that MC chymase or tryptase can shift colonic muscularis smooth muscle to a proinflammatory phenotype and that MC-conditioned medium exacerbates the overexpression of proinflammatory genes induced after EC irradiation (19,20). These results describe MCs as putative participants in the global tissue inflammation following irradiation. Moreover, MCs can activate the protease-activated receptor-2 (PAR-2), which is expressed by numerous cells including epithelial cells and nerve fibers. The importance of PAR-2 activation has been demonstrated by the increase in PAR-2 expression after radiation exposure and by the attenuation of the fibroproliferative process in irradiated MC-deficient rats (180). Recently, the involvement of MCs in radio-induced visceral hypersensitivity has also been demonstrated (43). Ketotifen treatment, which specifically blocks MCs, reverses radiation-induced mechanical allodynia. Moreover, the same study demonstrated, in patients developing late side effects of radiotherapy for rectal adenocarcinoma, an increase in the interaction between MCs and nerve fibers.
Fibrosis/Chronic Mesenchymal Cell Activation
Fibrosis is the main late complication of radiation therapy. It can occur several months to years after exposure to IR. Fibrosis is an evolutive process characterized by an excessive accumulation of ECM components, mainly collagen, often causing loss of tissue function (169). Fibrosis is described as an unchecked regeneration process that occurs under chronic injury conditions, caused for instance by bacterial infection, ischemia, or chronic inflammation (159,188). In the intestine, it can lead to occlusion due to the thickening of the intestinal wall and the loss of muscular function.
The characterization of the cellular and molecular mechanisms that initiate and perpetuate fibrosis after irradiation is still under debate. However, the involvement of cells from various intestinal compartments seems to be a matter of consensus. The myofibroblasts, or myofibroblast-like cells, are mainly responsible for the ECM production in radio-induced fibrosis. In the intestine, there are several sources of myofibroblasts. They differentiate from fibroblasts or local equivalents through the activation by various signals and are continuously activated during chronic inflammation. SMCs have been identified as the main cell type responsible for fibrosis in the intestine (182). Pericryptic fibroblasts and myofibroblasts, while able to undergo activation to a profibrotic state, are involved to a lesser extent. They are less abundant, and ulceration of the mucosa further reduces their number. Last, endothelial-to-mesenchymal transition and epithelial-to-mesenchymal transition can increase the pool of myofibroblasts (116,187). In these processes, endothelial or epithelial cells are able to transdifferentiate into mesenchymal cells. This large pool of potentially profibrotic cells makes the intestine particularly susceptible to fibrosis. An imbalance of dialogue between ECs and vascular SMCs can be involved in radiation-induced fibrosis (115). Irradiated ECs orient SMCs toward a profibrogenic phenotype: migration, proliferation, stimulation of synthesis of PAI-1, connective tissue growth factor (CTGF), heat shock protein 27 (HSP27), and collagens I and III.
Various pathways intervene in the differentiation of cells into myofibroblasts and their chronic activation. Irradiation leads to an acute inflammatory response with synthesis of profibrotic cytokines such as IL-4 and IL-13. Subsequently, a chronic inflammatory adaptive response induces differentiation of T lymphocytes toward TH2 or TH17 (depending on the model) that secrete profibrosing cytokines. It has also been reported that the interactions of radio-induced ROS with the EC compartment lead to inflammatory mediator secretion and thrombin synthesis that favor production of the profibrotic cytokine TGF-β. Recently, it has been proposed that the radiation effects to DNA can modify the epigenome of cells. These modifications (named as DNA methylation, histone acetylation, and miRNA synthesis) can be inheritable gene modifications that can induce and also perpetuate fibrogenesis even in the absence of the initial damaging stimulus (184). This hypothesis needs to be investigated and can provide a potential link between the acute radiation response and long-lasting radiation fibrosis.
Numerous signaling pathways, such as Rho, platelet-derived growth factor (PDGF), insulin-like growth factor (IGF), and TGF-β1, have been described to trigger fibrosis. TGF–β1 is considered as the main profibrotic cytokine. Active TGF-β can be processed in two ways: by direct induction of the activator protein-1 (AP-1) family transcription factor by IR and by activation of the latent form of TGF-β stored in the ECM. TGF-β associated with the latency-associated protein (LAP) is constitutively present in the microenvironment. ROS, RNS, and various cytokines generated in chronic radiation disease are involved in the release of TGF-β1 from LAP. Consequently, the activated TGF-β1 binds to its receptors and activates the Smad signaling pathway. The Smad family members activate various target genes (such as CTGF), and some of them are responsible for myofibroblast activation (12,73).
Management/Reduction of Radio-Induced Intestinal Damage
Therapists try to prevent the development of sequelae after radiotherapy by restricting the dose and the volume of healthy irradiated tissue. Innovations in tumor imaging allow precise delineation of the tumor, and use of three-dimensional (3D) conformal radiation therapy (3D-CRT) and intensity-modulated radiation therapy (IM-RT) techniques allows better tumor targeting and reduces the volume of normal tissue located in the irradiation field. However, it has been reported that 90% of patients develop acute side effects, leading to, in the worst cases, the arrest of antitumoral treatment. Moreover, 10% of patients develop late side effects (10 years after radiotherapy), which severely affect their quality of life and entail substantial mortality. There is currently no known permanent cure to treat the side effects of abdominopelvic radiotherapy. In more severe cases, such as intestinal obstruction, perforation, fistulas, and severe bleeding, the surgical resection of injured segments remains the only therapeutic option (139). These interventions are highly complicated by the presence of extensive inflammation and fibrosis leading to dehiscence of colonic anastomoses (57,89). To improve the outcome after surgery, a wide resection of margins (123) is performed, increasing the risk of short bowel syndrome that can entail a need for parenteral nutrition. Preventing the occurrence of radiation enteritis in the first instance represents the best approach to avoid surgical morbidity. Intestinal radiation damage is characterized by injuries of all compartments, compromising their role in the healing process. The multiplicity of targets involved in the physiopathology makes the development of therapeutic strategies extremely complex.
Medication of RT Side Effects
Despite the absence of treatment to counteract the consequences of irradiation of healthy tissue, clinicians use nutritional measures, antibiotics, and pharmacological agents, such as anti-inflammatory drugs. Table 1 lists examples of the available treatments to manage gastrointestinal symptoms. However, further investigations are needed to demonstrate their effectiveness. It is also important to note that, as late radiation gastrointestinal toxicity shares numerous symptoms with inflammatory bowel disease, clinicians emphasized the importance of detecting pelvic radiation disease to rapidly provide appropriate and effective treatments to patients (5,165).
Treatments for Gastrointestinal Symptoms Associated With Radiotherapy

New Molecules for Improving Regeneration After Irradiation
Studies analyzing the healing process of the intestine suggest that all cells and compartments of the digestive tract are sequentially involved, in particular epithelium/mesenchyme/vasculature/immune and ENS. Clinical and laboratory research, as well as detailed knowledge of the mechanisms of action of the different actors in all tissue compartments, are essential for the development of new therapeutic strategies to prevent radio-induced damage and/or improve tissue regeneration. Table 2 summarizes the different molecules that have been tested to reduce intestinal radio-induced toxicity and details their effects on specific compartments. Although these treatments could potentially reduce irradiation-induced damage, none has been shown to have pleiotropic action, contrary to stem cell-based strategies.
Molecules to Improve Regeneration After Irradiation

TGF, transforming growth factor; EGF, epidermal growth factor; FGF, fibroblast growth factor; KGF, keratinocyte growth factor; IGF, insulin-like growth factor; HGF, hepatocyte growth factor; GLP-2, glucagon-like peptide-2; PGE2, prostaglandin E2; IFN, interferon; VEGF, vascular endothelial growth factor; TLR, Toll-like receptor; NF-κB, nuclear factor-κB.
New Therapeutic Approaches: Cell and Tissue Engineering
MSC Treatment
MSCs were initially identified in the bone marrow, but are also found in other tissues such as cartilage, skeletal muscle, tendon, adipose tissue, circulating blood, and placenta (175). A number of preclinical studies have highlighted structural and functional benefits of MSC transplantation in different pathologies such as models of myocardial infarction, muscular dystrophy, fibrosis, and bone or tendon defects (169). In humans, MSCs have been shown to be an effective therapy for patients suffering from graft-versus-host disease who develop gastrointestinal lesions (92). A phase II clinical study on the effect of MSC treatment on perianal fistulas in Crohn's disease patients has shown promising results (60). Furthermore, long-term follow-up of these patients for 10 years showed that MSC treatment is not associated with adverse effects (69).
Managing Intestinal Radiation Injuries Using MSC Regenerative Therapy
In various animal models, intravenous injections of MSCs have been shown to improve the intestinal function disrupted after irradiation (Fig. 3). The ability of human MSCs to restore the intestinal function (absorption and secretion capacity) was highlighted for the first time in NOD/SCID mice irradiated at a dose of 8.5 Gy in the abdominal region (142,151). Likewise, the goblet cell content, which decreased in colonic crypt cells in a model of colorectal irradiation in rats, was reestablished after MSC treatment. In addition, increased expression of the adherent junction protein (β-catenin) in the colonic mucosa was shown after MSC injection (149). A functional therapeutic benefit of MSC injection was also observed after fractionated irradiation of the prostate. Indeed, MSCs restore erectile function measured by intracavernous pressure in response to electrostimulation of the cavernous nerve (136). A recent study demonstrated that MSC treatment reverses persistent visceral allodynia in rats exposed to colorectal irradiation. This study demonstrates that MSCs reduce the fecal tryptase level and the number of anatomical interactions between MCs and nerve fibers, suggesting their role in stabilizing MCs and reducing activation of nerve fibers (43).

Use of MSCs to improve radio-induced intestinal injury. This figure summarizes the functional benefits induced after MSC treatment (green oval) in various preclinical studies. It also highlights the ability of MSCs to target various intestinal compartments (orange squares). Indeed, MSCs have been reported to induce cellular and molecular events in the epithelial and vascular compartments as well as on inflammatory and fibrosis processes.
The therapeutic benefit of MSC injection on the intestinal structure was also observed. The MSC-treated animals present an increase in the crypt–villus axis compared with those having received abdominal irradiation (30,150,151). Furthermore, after colorectal irradiation inducing irreversible lesions in rats, intravenous injection of MSCs decreases the epithelial injury score in a cell dose-dependent manner (149).
Cellular Events Induced After MSC Infusion
The accelerated reepithelization process after MSC injection of the radiation-exposed intestine is associated with a decrease in epithelial cell apoptosis (142,149,151) and stimulation of epithelial proliferation (30,142,149,151) (Fig. 3). More precisely, these studies demonstrated an increase in intestinal progenitor/stem cells (30,149, 151) with a higher number of Musashi-1, SOX9-high positive, or Bmi1.
MSCs have also been reported to modulate radiation-induced inflammation (Fig. 3). T lymphocytes are involved in intestinal radiation toxicity (14,98), and MSC injection reduces the infiltration and proliferation of CD4 T cells and increases the apoptosis of radiation-activated T cells (14). After colorectal irradiation in rats, we also demonstrated for the first time the predominance of the TH17 subset, largely described for its role in profibrotic diseases and their repression after MSC injection (13). However, MSCs do not modify IL-10 secretion by Tregs after abdominopelvic irradiation. This may be due to the lack of Tregs, generated after irradiation, to produce IL-10 (17). In a minipig model of severe colorectal damage induced by localized irradiation, iterative injections of autologous MSCs decreased the macrophage infiltration associated with wound healing (99). Studies showed a decrease in neutrophils infiltrate in the gut after injection of human MSCs (30,122). In addition, Ono et al. demonstrated a decrease in macrophage infiltrate in the rectum 3 days after injection of human amnion-derived MSCs (122). However, in a rat model with severe radio-induced mucosal damage, injection of rat MSCs induced no modification of innate cell infiltration (14). Importantly, MSC treatment reduces the radiation-induced fibrosis by reducing collagen deposition and expression of col1a1/col1a2/col3a1 (13,99) (Fig. 3).
Molecular Mechanisms of the Therapeutic Benefit of MSC Therapy
The mechanisms involved in the improvement of tissue healing by MSCs vary depending on the type of lesion and tissue damage. A complete understanding of these mechanisms therefore includes the identification of the complex interactions between MSCs and their environment. However, a large number of results published on various experimental models suggest that MSCs act through the secretion of a wide variety of bioactive (angiogenic, mitogenic, and antiapoptotic) molecules (28). Indeed, MSC-conditioned media accelerate the recovery of radio-induced structural damage (59). Moreover, microvesicles and exosomes produced by MSCs also demonstrate a therapeutic benefit in animal models of tissue regeneration, myocardial ischemia/reperfusion injury, hindlimb ischemia, and liver fibrosis (3,96).
To exert their therapeutic benefit, injected MSCs should home to the injury site. A quantitative study revealed that MSCs are able to migrate more abundantly toward the irradiated tissue, including the small intestine (55). The proinflammatory context induced in mucosa after irradiation can attract MSCs to the injury site. Moreover, we also demonstrated in colorectal-irradiated rats that MSC infusion induces plasmatic stromal cell-derived factor-1α (SDF-1α) associated with an increase in endogenous circulating MSCs. We can therefore hypothesize that endogenous MSCs ensure the continuation of tissue regeneration (Fig. 4). The local microenvironment induced after irradiation can be unfavorable to the long-term survival of MSCs. However, several studies demonstrated that the inflammatory microenvironment is necessary to stimulate MSCs in order to achieve their full therapeutic benefit. Indeed, proinflammatory molecules, such as IFN-γ, are very important for stimulating MSC secretion and their regenerative capacities (27).

Schematic representation of MSC behavior after infusion into the blood circulation of colorectal irradiated rats. 1) Exogenously amplified MSCs were injected by intravenous route. 2) MSC infusion following colorectal irradiation demonstrated a high number of MSCs trapped in the lungs initially and for 3 days after injection. This is probably due to the incapacity of the larger MSCs to pass through the lung capillaries. Lung-trapped MSCs have been reported to improve myocardial infarction by secretion of molecules released into the blood circulation and acting through the abscopal effect (93). Such a mechanism can also be responsible for the initial benefit induced by MSCs on colonic epithelial regeneration following irradiation. 3) After MSC injection, a higher amount of SDF-1α chemoattractant molecule is retrieved in the plasma. 4) Associated with the SDF-1α increase in the plasma, an increase in endogenous circulating MSCs has been observed. This result suggests that MSC therapy, using secreted factors, can enhance the pool of endogenous MSCs by mobilizing them probably from organ storage. This endogenous MSC mobilization can indirectly potentiate and prolong the therapeutic effect of exogenous MSCs. This hypothesis could explain the therapeutic benefit of MSCs even if the level of infused MSC engraftment at the injury site is very low or undetectable with current techniques. However, the distribution of exogenous MSCs in the body following their systemic administration has demonstrated that MSCs migrate preferentially to the irradiated area (55). It can be hypothesized that MSCs will engraft in damaged tissue through the same mechanism as leukocytes. 5) Irradiation of the tissue leads to secretion of chemoattractants and proinflammatory molecules such as IL-6, IL-8, TNF-α, and IL-1β that can attract circulating MSCs. Thus, exogenous and also endogenous MSCs may be able to leave the bloodstream and migrate to the damaged tissue.
As detailed previously, radiation-induced intestinal toxicity is complex and involves various cellular compartments. Because of their large panel of molecule expression, MSCs seem to be the ideal candidate to promote intestinal regeneration after irradiation. Even if the underlying molecular mechanisms of action of MSCs are not fully understood, MSCs have multiple therapeutic effects. The most described are antiapoptotic, proproliferating, proangiogenic, and anti-inflammatory. MSCs reduce the radiation-induced apoptosis of epithelial cells (142,149). Even if in vivo MSC antioxidative properties are difficult to highlight, studies using gene-modified MSCs to overexpress the molecules involved in oxidative stress showed superior effectiveness in an intestinal irradiation model (79,191). Saha et al. reported that MSCs can protect irradiated intestine by increasing the levels of R-spondin, KGF, and bFGF protein, which promote proliferation and inhibit apoptosis (142). One study described an increase in the proliferation of crypt progenitor cells, positive for the SOX9 marker, after colorectal irradiation and systemic infusion of MSCs (149) (Fig. 5). This proregenerative effect of MSC injection is Wnt4 dependent. Furthermore, MSCs were demonstrated to promote neovascularization in irradiated small intestine (Fig. 3). The authors demonstrated that the number of ECs and also the naive ECs increases in MSC-treated animals associated with an increase in vascular endothelial growth factor (VEGF) gene expression in the intestine (99) and PDGF in the serum (142). In a pig model, rectal irradiation increased the vascular density in the rectum, associated with an increase in VEGF gene expression in the colon, rectal mucosa, and anus (99). Moreover, numerous studies reported anti-inflammatory effects of MSC treatment in various diseases and after irradiation. Downregulation of proinflammatory cytokines (IL-6, IL-1β, TNF-α, and IL-8) in pigs and IL-17 in rats was observed in the mucosa of colorectal-irradiated animals treated with MSCs (13, 99). Using enzyme-linked immunosorbent assay (ELISA) tests on irradiated intestine and colon, a reduction of IL-6 or IL-1β secretion by MSCs was not detected (14,142). However, the anti-inflammatory cytokine IL-10 increases rapidly following MSC treatment (142). Another study reported a significant increase in IL-10 at 5 days after MSC treatment (64,65). However, 10 days after MSC treatment, this increase was less important. These data can explain the absence of an increase in IL-10 at 21 days after colorectal irradiation in colonic mucosa (14) and the importance of repeated injections of MSCs (99). In addition, we also demonstrated that colorectal irradiation leads to stimulation of the synthesis of endogenous glucocorticoids in colonic mucosa and that MSC injection sustains this anti-inflammatory process over time (14). These data confirm the anti-inflammatory properties of MSCs and highlight their abilities to stimulate existing endogenous processes of intestinal regeneration. MSC injection also reduces radiation-induced fibrosis by decreasing the collagen gene expression associated with the modulation of the matrix metalloproteinase (MMP)-to-tissue-inhibitor of metalloproteinase (TIMP) ratio (13,99). Altogether, these preclinical data suggest that MSCs, by secreting various bioactive factors, reduce severe radiation-induced ulcers by activating areas in the margin of the lesions, leading to stimulation of the endogenous regeneration process. This benefit is effective in multiple cellular compartments (summarized in Fig. 3), leading to an increase in animal survival following irradiation of the abdominopelvic area (30,149,151).

Mesenchymal stromal cell treatment stimulates epithelial proliferation and increases the number of SOX9-high-positive progenitor/stem cells. SD rats are subjected to colorectal irradiation, while MSCs from rat bone marrow were grown in parallel. After irradiation, animals were intravenously injected with 5 × 106 of MSCs. PCNA and SOX9 immunostaining was evaluated 1 or 2 weeks after injection. Representative pictures of PCNA immunostaining (blue staining) 2 weeks after irradiation. Original magnification: 650×. Representative pictures of SOX9 immunostaining (blue staining) 2 weeks after irradiation. Original magnification: 850×. Quantification of SOX9-low- and SOX9-high-expressing cells by crypt. *p < 0.001 versus control groups.
Clinical Outcomes and Irradiation
MSC transplantation was used on compassionate grounds for the first time in the world in 2005 in a severe radiological burn victim hospitalized at the Percy HIA (91). Seven patients who had accidentally received very high doses of irradiation and developed severe radiation-induced burns were treated with repeated local injections of MSCs, in combination with surgical excision of the necrotic areas, followed by autologous skin transplantation or a skin and muscle flap. The results were spectacular, and the patients recovered the use of their limbs, which would have been impossible with conventional treatment (15,91,162). In addition to clinical applications on the skin, MSC injections have been used on compassionate grounds to treat patients who have been overirradiated during radiotherapy for prostate cancer (131). Three patients suffering from rectal bleeding and pain, signs of severe radiation-induced proctitis (grade IV according the SOMA-LENT scale), and who were refractory to symptomatic treatment were treated with MSCs. Although it was difficult to assess the treatment because of its compassionate nature, a decrease in T-cell activation and an increase in the number of Tregs were of particular significance (173). In addition, a decrease in pain sensitivity was experienced by the patients after injection of the cells. One of the patients was even able to stop opioid analgesic therapy during this period. As the beneficial effect of the MSC injection is temporary, repeated injections are required (173).
Nowadays, MSC treatment of severe radiation-induced colonic lesions is only used in the clinic on a compassionate basis. The absence of side effects after MSC injection as well as a better comprehension of mechanisms of action may, in the future, allow using MSCs as soon as the first symptoms appear. However, the use of MSCs as a prophylactic treatment, before radiotherapy, seems unrealistic since it has been shown that MSCs migrate to the site of the tumor (96) and could therefore be a stroma for cancer cells. Studies demonstrated the ability of MSCs to resist irradiation because of their high antioxidant capacity (31). Moreover, even if irradiation does not impair the immunosuppressive capacity of MSCs in vitro (38), it has been demonstrated that irradiation can modify MSC phenotype, their capacity to differentiate, and their molecule secretion profile (49). These modifications could alter their therapeutic benefit.
Optimizing the Therapeutic Properties of MSCs
Although MSCs have been shown to improve the intestinal regeneration process after irradiation, the low implantation and survival rates of the cells in the injured tissue limit their therapeutic benefit. Various studies demonstrate that this feature is common to various diseases. Consequently, this has prompted efforts to improve the therapeutic properties of MSCs (Fig. 6). First, the best route of administration of cells has been tested. Indeed, local injection of MSCs is preferred wherever possible by surgeons. This method of administration allows fewer cells to be injected, which are directly implanted into the site of lesion, and reduces the spreading of stem cells throughout the body. Other strategies have also been tested to increase MSC homing to the site of lesion: particularly, the combination of MSC injection with a mobilizing molecule or genetic modification of MSCs to overexpress the CXCR4 receptor. The poor viability of the injected MSCs in the tissue can be due to various apoptotic signals. MSCs have a fibroblastic morphology and are attached to the culture plate during their in vitro amplification. The first stress occurs when the cells are detached from the culture dishes for the engraftment procedure. Once MSCs are locally injected into the damaged tissue, their lack of adhesion to ECM can lead to anoikis, which promotes apoptotic signaling. In addition, the innate immune cells like natural killer (NK) cells may also participate in MSC lysis (163). More recent studies demonstrated that preconditioning MSCs with TLR ligands protects them from death by NK cells (189) and enhances their therapeutic potential (70). Moreover, the host inflammatory microenvironment, which generates proinflammatory molecules and ROS, can also trigger apoptotic signaling. Strategies to decrease the apoptosis of MSCs have been successfully realized in animal models. Even if genetically modified MSCs would be difficult to use in the clinic, these experiments shed light on the correlation of the therapeutic benefit and inhibition of MSCs apoptosis. Other studies demonstrated that modification of the host by anti-inflammatory molecules or hyperbaric oxygen, already in clinical use, improves the outcome of MSCs (86,192). In the context of tissue engineering, growing evidence suggests that biomaterials can be used as scaffolds to protect MSCs from the environment and thus increase their life span.
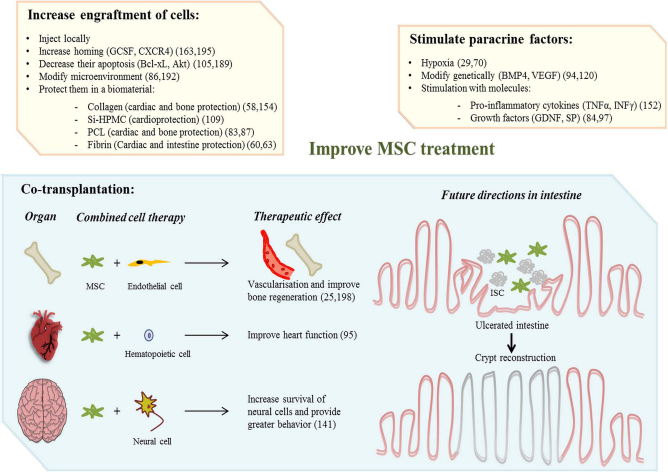
Strategies to improve MSC therapeutic benefits. Upper panels in yellow described the options used to optimize paracrine effects of MSCs. Increasing the engraftment of cells: 1) local delivery of MSCs is preferred wherever possible by surgeons. This method of administration allows fewer cells to be injected, which are directly implanted into the site of lesion, and reduces the spread of stem cells throughout the body, even though no side effects have been shown after MSC injection (164). 2) Engraftment in the injured area can be increased by coinjection of MSCs with mobilizing molecules (163) or genetic modification of MSCs (195). One study demonstrated that preconditioning of animals with hyperbaric oxygen increases MSC engraftment in infarcted myocardium (86). 3) Improving MSC survival by overexpressing antiapoptotic molecules. 4) Protection of the MSCs in biomaterials: cell encapsulation allows the diffusion of nutrients and oxygen, both essential for the survival of cells, while at the same time protects the implanted cells from the host's immune response and avoids the entrance of high molecular weight immune system components such as immunoglobulins and immune cells. The choice of the biomaterial (resolvability and biocompatibility) is very important depending on the characteristic of the tissue. 5) Stimulating paracrine effect: in vitro priming of MSCs by a wide variety of techniques, including hypoxia or molecule preconditioning. Cell engineering could be used to influence cell fate after in vivo injection. This strategy has been largely tested to increase the proangiogenic properties of MSCs by stimulating VEGF expression or to improve the anti-inflammatory capacities of MSCs. Lower panel in blue schematizes the combined cell-based therapies that have been tested in preclinical models to repair bone, heart, and brain (left) and future directions that can be envisaged to repair irradiated intestine (right). Cotherapy with MSCs can broaden the spectrum of action combining the beneficial effects of each cell type. As ISCs are destroyed by irradiation, combining ISCs with the anti-inflammatory and proangiogenic, proproliferative MSCs can improve the structural and functional benefit and accelerate the regeneration process after high doses of irradiation.
The reparative potential of MSCs seems to be mediated by the release of a broad spectrum of soluble factors. This has prompted considerable efforts to enhance these paracrine capacities by various preconditioning strategies listed in Figure 6. Beyond the determination of the best route of injection, protection against apoptotic signals, and stimulation of their paracrine capacities, the question arises of the ability of one type of cells to improve entire organ regeneration. Because of their intrinsic supportive properties, MSCs are the ideal candidate to use in combination with other progenitor cells. Combined cell therapy has been tested in animal models to repair different tissues (Fig. 6). Significant challenges exist in the development of the intestinal mucosa with the successful ability to grow mini-gut structure in vitro into a 3D scaffold. Besides, in vivo engraftment of colonic progenitor cells has been realized in mice (194). Vacanti's lab has achieved encouraging results by seeding epithelial cells on a biodegradable scaffold and implanting them into host animals (67). At present, efforts are necessary to induce ISC growth on a tubular scaffold by using specific molecules or cell types to obtain a complex architecture that resembles the native intestine. This will require multidisciplinary collaboration to bring fundamental stem cell science, biomaterial engineering, and surgeons into closer contact to overcome the complexity of the intestine. Regenerative therapy for radiation injury is an important juncture. These innovative tissue engineering strategies will offer new hope to patients suffering from the severe side effects of radiotherapy.
Footnotes
Acknowledgment
The authors declare no conflicts of interest.