
Abstract
Bone nonunion treatments pose a challenge in orthopedics. This study investigated the joint effects of using mesenchymal stem cell (MSC) sheets with local injection of stromal cell-derived factor-1 (SDF-1) on bone formation. In vitro, we found that migration of MSCs was mediated by SDF-1 in a dose-dependent manner. Moreover, stimulation with SDF-1 had no direct effect on the proliferation or osteogenic differentiation of MSCs. Furthermore, the results indicated elevated expression levels of bone morphogenetic protein 2, alkaline phosphatase, osteocalcin, and vascular endothelial growth factor in MSC sheets compared with MSCs cultured in medium. New bone formation in fractures was evaluated by X-ray, micro-computed tomography (micro-CT), hematoxylin and eosin (H&E) staining, Safranin-O staining, and immunohistochemistry in vivo. In the rat bone fracture model, the MSC sheets transplanted into the injured site along with injection of SDF-1 showed significantly more new bone formation within the gap. Moreover, at 8 weeks, complete bone union was obtained in this group. In contrast, the control group showed nonunion of the bone. Our study suggests a new strategy involving the use of MSC sheets with a local injection of SDF-1 for hard tissue reconstruction, such as the healing of nonunions and bone defects.
Keywords
Introduction
Despite increasing insight into fracture healing and delayed healing, nonunion formation remains a clinical problem and is a frequent complication in orthopedic surgery. It has been estimated that 5–10% of all long bone fractures have delayed union or develop into a nonunion 1 . Many strategies have been developed to treat bone nonunion, but no consensus on optimal management has emerged thus far. Autologous bone grafts or free vascularized bone grafts are still considered the gold standard for treatment2–4. However, the slow healing of allografts and high donor site morbidity 5 often result in additional clinical problems. Moreover, the possible sources of graft material are limited, especially if previous harvests have been performed.
Cell-based treatment, which is based on bone tissue engineering and has been the focus of widespread research interest in recent years, is considered a promising treatment for certain diseases. Many groups have reported success in healing bone defects with different types of cells 6 , such as mesenchymal stem cells (MSCs). MSCs are multipotential cells that can differentiate into various lineages, such as bone, adipocytes, cartilage, and ligaments. They can be isolated easily from the bone marrow and augmented by culture in vitro. It is possible to harvest billions of cells from 10 ml of bone marrow. Accordingly, many investigators have employed a tissue engineering strategy that constructs MSC scaffold composites to improve bone regeneration 7 . However, conventional methods of tissue engineering involving the seeding of cells into biodegradable scaffolds or the injection of isolated cell suspensions present a number of problems, including small quantities of seeded cells, cell loss, and inflammatory reactions with degradation of the scaffolds. To overcome these limitations, several groups have attempted to use the cell sheet technique for tissue engineering, as it can provide highly efficient cell delivery and maintain intact cell-to-cell contact8,9. In addition, the scaffold-free tissue engineering technique is beneficial in that it eliminates the inflammatory reactions related to the use of biological materials 10 . Using cell sheet engineering can overcome the limitation of conventional tissue engineering approaches such as using isolated cell injections and scaffold-based technologies 11 . It has been confirmed in a rat model that MSC sheets can form new bone after transplantation 12 . For bone regeneration, the cell sheet can be lightly layered and attach strongly to the sites of large bone defects, and the osteoblasts attached to the mineralized layered cell sheet may mimic the deposition of extracellular matrix of bone tissue in vivo 13 . Therefore, we explored the effect of MSC sheet techniques in order to seed cells for bone regeneration.
As several researchers investigate tissue-specific organ repair or regeneration in the liver 14 , heart 15 , skin 16 , and bone17,18, MSCs can migrate to the site of injury and contribute to tissue regeneration. This migration is mediated by stromal cell-derived factor-1 (SDF-1, also called CXCL12), the expression of which is upregulated at sites of injury. SDF-1 is a chemokine that regulates stem cell homing 19 by binding to its receptor, CXCR4 20 , and has been reported to play important roles in the repair or regeneration of tissue by contributing to the recruitment of circulating or resident CXCR4-expressing MSCs. Moreover, SDF-1 also increases the survival and growth of CXCR4-expressing stem cells, such as MSCs, both in vitro 20 and in vivo21,22. It has been demonstrated that local delivery of SDF-1 is effective for recruiting MSCs and endothelial progenitor cells (EPCs), both of which were beneficial for bone regeneration 23 . Local injection of SDF-1 appears to be effective because it has dose-dependent effects on the migration of MSCs, and local application helps to prevent the systemic side effects of drugs.
On the basis of these observations, we hypothesized that the use of MSC sheets with local injection of SDF-1 would enhance bone regeneration in fracture sites compared with the effects of MSC sheets alone, MSCs (dispersed in medium) with SDF-1, SDF-1 alone, MSCs alone, or no treatment (control condition) in a rat model. Our results suggest new strategies for using SDF-1 and MSC sheets to promote successful bone regeneration.
Materials and Methods
Fabrication of MSC Sheets
Primary rat MSCs were isolated and cultured on the basis of the methods described previously 22 . Briefly, young adult male Sprague–Dawley rats (Experimental Animal Center of Zhejiang Academy of Medical Sciences, Hangzhou, P.R. China) were euthanized with CO2, and the femurs and tibias were then removed. The bones were washed in α-minimal essential medium (α-MEM) supplemented with 10% penicillin/streptomycin (Gibco, Grand Island, NY, USA). The bone marrow cavities were flushed with 10 ml of basal medium. The isolated marrow was centrifuged (400 × g), resuspended in 10 ml of α-MEM supplemented with 10% fetal bovine serum (FBS; Gibco), seeded in 75-cm2 tissue culture flasks, and cultured in a humidified atmosphere of 5% CO2 at 37°C. The medium was changed every 3 days to remove hematopoietic and other unattached cells. The adherent cells were cultured until confluence, and then MSCs were passaged after digestion with 0.25% trypsin–ethylenediaminetetraacetic acid (EDTA; Gibco). MSCs at passages 1 and 2 were used for cell sheet engineering.
To create the MSC sheet, MSCs were seeded on culture dishes at a density of 4 × 104 cells/well and cultured in α-MEM supplemented with 10% FBS, 1% penicillin, streptomycin, and 50 μg/ml vitamin C (Sigma-Aldrich, St. Louis, MO, USA) at 37°C in a humidified atmosphere with 5% CO2 for approximately 1 week until cell sheet formation. The cell sheet was then lifted using a scraper (Fig. 1A). Four cell sheets were digested with collagenase (KeyGEN, Nanjing, P.R. China) for quantification of cell number.

(A) Cultured mesenchymal stem cells (MSCs) were lifted as a cell sheet using a scraper. (B) A transverse osteotomy was performed in the proximal one third of the tibia, and the periosteum at the fracture site was removed. (C) A cell sheet was transplanted onto the osteotomy site.
Proliferation of MSCs Stimulated by SDF-1 In Vitro
In cell proliferation assays, cells were cultured in either maintenance medium (control group) or in maintenance medium supplemented with various concentrations of recombinant rat SDF-1α (PeproTech Inc., Rocky Hill, NJ, USA). Cell number was determined using a cell counting kit-8 (CCK8) according to the manufacturer's instructions (Dojindo Laboratories, Kumamoto, Japan). The growth curves of MSCs cultured at different concentrations were drawn, and each experiment was repeated three times.
Real-Time Quantitative PCR of MSCs and MSC Sheets
We measured the gene expression levels of bone morphogenetic protein 2 (BMP-2), alkaline phosphatase (ALP), osteocalcin (OCN), and vascular endothelial growth factor (VEGF) to determine the osteogenic and angiogenic potentials of the cells. RNA was isolated from cells cultured in medium (MSC group), cell sheets (MSC sheet group), SDF-1-induced MSCs (SDF-1 group), and SDF-1-induced MSC sheets (SDF-1 sheet group) using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's protocol and converted to cDNA using a commercial kit (SuperScript II Reverse Transcriptase; Invitrogen) according to the manufacturer's protocol. Each group occupied four wells in multiwell plates. Realtime quantitative polymerase chain reaction (RT-qPCR) was performed with a SYBR Premix ExTaq Kit (TaKaRa, Otsu, Japan) on an iQTM5 multiplex real-time fluorescence quantitative PCR instrument (Bio-Rad, Richmond, CA, USA) according to the procedures described previously 12 . The nucleotide sequences of primers and probes for rat cDNAs are listed in Table 1. 18S ribosomal RNA was used as an internal control to adjust for the differences between samples. Thermal cycle conditions were as follows: 1 min at 95°C for activation of the Universal Mixture AmpliTaq Gold Polymerase (Invitrogen), followed by 45 cycles of 10 s at 95°C for denaturation and 25 s at 62°C for annealing and extension.
Nucleotide Primers Used for Real-Time Quantitative Polymerase Chain Reaction

ELISA
SDF-1 was quantified in cell culture supernatants using an enzyme-linked immunosorbent assay (ELISA) kit (IBL-America, Minneapolis, MN, USA) according to the manufacturer's protocol. To obtain the cell culture supernatants, equal numbers of MSCs were seeded in the same volume of medium in culture plates of different sizes. After 24 h, cells were divided into two groups: the MSC group and the MSC sheet group. The medium was changed after the MSC sheet had formed to remove residual growth factors. Supernatants were collected 24 h after medium was replaced. The results are presented as means ± standard error of the mean (SEM).
Migration Assay
The effects of exogenous SDF-1α on the migration of MSCs were evaluated using a Transwell system (Corning; Corning, NY, USA). For exogenous SDF-1α, 3 × 104 cells in 100 μl of serum-free α-MEM were placed within the upper chamber. To observe migration, 600 μl of SDF-1α (200 ng/ml; PeproTech Inc.) in serum-free α-MEM was added to the lower chamber. To observe chemokinesis, SDF-1α was also added to the upper chamber. For further confirmation, the cells in some groups were preincubated with an antagonist of SDF-1, AMD3100 (200 ng/ml; Sigma-Aldrich), for 30 min at 37°C. In a separate chemotaxis assay, the upper chambers were loaded with 5 × 104 cells in 100 μl of serum-free MEM, and the lower chambers were loaded with 600 μl of SDF-1α at different concentrations (PeproTech Inc.). We used SDF-1α at 0, 100, 200, and 500 ng/ml according to the manufacturer's instructions.
After incubation for 18 h, the upper surface of the filters was scraped free of cells and debris. Cells that had migrated through the filter were fixed in 4% paraformaldehyde (PFA; Sigma-Aldrich) and subjected to crystal violet staining (Sangon Biotech Co. Ltd., Shanghai, P.R. China). The total number of cells in five randomly selected fields on the lower surface of the membrane was counted under a light microscope (100×; Leica, Wetzlar, Germany). The data were expressed as mean cell number/high-power field (HPF) ± standard deviation (SD) and subsequently analyzed to identify statistically significant differences between experimental groups. Data were confirmed in three independent experiments.
Animal Model
An experimental rat tibia osteotomy model was made as described previously 12 . The animals were treated in accordance with standard ethical guidelines, and the experimental protocols were approved by the Zhejiang University Ethics Committee. The rats were anesthetized with pentobarbital (3.5 mg/100 g weight; Sinopharm, Shanghai, P.R. China) administered intraperitoneally. A lateral incision (1.5 cm) was made on the proximal site of the tibia, and the muscle was divided longitudinally to expose the tibia. The periosteum was removed from the proximal to the distal site of the tibia to the extent possible. A transverse osteotomy was then performed from front to back in the proximal one third of the tibia using an oscillating mini saw (Fig. 1B). A small incision was then made on the medial aspect of the knee, and the patella was deflected laterally to expose the tibiae, where a small hole was drilled. A 21-gauge needle was then inserted through the hole into the distal end of the tibia, resulting in loose fixation. After suturing the cut, 2 ml of SDF-1α solution (200 ng/ml) was injected into the fracture area. After 1 week, 2 ml of SDF-1α (200 ng/ml) was injected again into the implantation site. One hundred forty osteotomy sites of 70 rats were transplanted with the MSC sheet alone (n = 11), with the MSC suspension (MSCs in medium, the number of cells is the same as the quantity in MSC sheets) alone (n = 11), with SDF-1α alone (n = 11), with the MSC sheet and injected with SDF-1α (n = 11) (Fig. 1C), with the MSC suspension (MSCs in medium) and injected with SDF-1α (n = 11), or implanted with an MSC sheet consisting of green fluorescent protein (GFP)-MSCs and injected with SDF-1α (n = 4). Untreated osteotomy sites served as controls (n = 11). Unprotected weight bearing was allowed immediately after the operation. Data on all experimental groups are shown in Table 2.
Experimental Groups

Real-Time PCR Analysis
For gene expression analysis, three tibia specimens were used for each group to evaluate BMP-2, ALP, OCN, and VEGF gene expression at 2 weeks after surgery. Three bone samples incorporating the individual fracture region were obtained from the tibia of each rat. The sequences of primers for target genes and the internal control gene are listed in Table 1. The extraction of total RNA from bone samples, synthesis of cDNA, and real-time PCR quantification were performed according to the methods described above.
Image Examination
Bone union was evaluated from X-ray photographs and micro-computed tomography (micro-CT). X-ray photographs were taken with animals under anesthesia at 4 and 8 weeks postoperatively to evaluate callus formation and bridging bone formation at the fracture site. Radiographs were taken with a dual track molybdenum/rhodium+ Mo target mammography machine (22 kV, 250 mAS; GE Healthcare, Fairfield, CT, USA). Gray values of the fracture sites and healthy tibias were analyzed using medical image analysis software [ImageJ 1.43u; National Institutes of Health (NIH), Bethesda, MD, USA], and the scales were set between 0 and 255. In addition, the therapeutic results after 4 and 8 weeks were determined through three-dimensional (3D) reconstruction of micro-CT images (μCT 40; Scanco Medical, Brüttisellen, Switzerland). The rat tibias were scanned from the proximal to the distal direction along the long axis of the tibia. A small animal scanning setting (70 kVp, 114 mA, 200-ms integration time) was used for the micro-CT scanning, resulting in an image pixel size of 9.8. The operator used a 2D preview X-ray (scout view) to determine the region of interest (ROI) to be scanned. This region consisted of 201 slices starting from the first slice containing the 10-mm distance to proximal bone fracture and moving distally throughout the whole fracture area. The 3D images were smoothed using a Gaussian filter and segmented following the specimen-specific global thresholding. On the basis of the bone images, the bone volume (BV, mm3), tissue volume (TV, mm3), and bone surface (BS, mm2) were measured. The percent bone volume (BV/TV, %) and bone surface density (BS/TV, mm−1) were analyzed to determine bone quantity.
Histological Preparation and Immunohistochemical Analysis
Samples were fixed in 10% PFA, decalcified with 10% EDTA (pH 7.4) for 4 weeks, and embedded in paraffin. Sagittal sections (7 mm thick) from each sample were prepared, stained with hematoxylin and eosin (H&E; Sigma-Aldrich) and Safranin-O (Sigma-Aldrich), and examined under a light microscope (Leica, Wetzlar, Germany). After H&E staining, tissue sections were randomly chosen and examined under a light microscope. Digital images were analyzed by Image-Pro Plus (IPP) 6.0 software (Media Cybernetics Inc., Acton, MA, USA). The percentage of newly formed bone area in the fracture area was calculated following a previous report wherein percentage of new bone tissue area = new bone area/fracture area × 100 % 24 . The fracture area was defined from the histologically visible cut edge of the cortical bone, and new bone tissue was identified by the woven structure.
For immunohistochemical (IHC) assessment, bone sialoprotein (BSP) and osteopontin (OPN) expression were determined according to the following procedure. In addition, IHC staining for CD34 was performed to analyze angiogenesis. Deparaffinized sections were washed with phosphate-buffered saline (PBS) and incubated with 0.3% hydrogen peroxide (Sigma-Aldrich) for 20 min to block endogenous peroxidase activity; this was followed by incubation with 5% bovine serum albumin (BSA; Sigma-Aldrich). The sections were then incubated overnight at 4°C with optimal concentrations of primary antibodies for BSP (1:100; Abcam, Cambridge, MA, USA), OPN (1:200; Abcam), or CD34 (1:200; Abcam). After washing three times with PBS, sections were incubated with biotinylated secondary antibody (polyclonal antibody; Zhongshan Biotechnology Co. Ltd., Zhongshan, P.R. China) for 20 min before incubation with horseradish peroxidase-conjugated avidin-biotin complex (ABC) (Zhongshan Biotechnology) for a further 20 min, followed by buffered 3,3′-diaminobenzidine tetrahydrochloride (DAB) (Zhongshan Biotechnology) as a chromogen. Sections were then counterstained with hematoxylin, dehydrated with increasing concentrations of ethanol solutions xylene, and mounted with coverslips. Stained sections were randomly chosen and examined under the light microscope.
For BSP and OPN, three randomly selected nonoverlapping images from every section were measured. The mean optical density (OD) of BSP and OPN staining was estimated by Image-Pro Plus 6.0 software (Media Cybernetics). For CD34, 10 different fields within the regenerated tissue were randomly selected from each section, and the number of CD34+ vascular endothelial cells (ECs) per field was counted. The mean of 10 values was used to determine the blood vessel density 25 .
Detection of Donor Cells Contributing to New Bone Formation
We prepared an additional experimental group (GFP-MSC group) to verify whether the donor cells in the cell sheet contributed to bone formation around the fracture site by detecting MSCs transduced with the GFP gene. The lenti-virus-based GFP-transfected MSCs (GFP-MSCs) were purchased from Cyagen Biosciences (Guangzhou, P.R. China). Reagents and culture methods were as described for primary MSC culture after resuscitation of frozen GFP-MSCs (Cyagen). MSC sheets and the rat model were prepared by the method described above. At 2 and 8 weeks after transplantation, half of the samples were initially observed using the Maestro In-Vivo Imaging System [Cambridge Research & Instrumentation, Inc. (Cri), Woburn, MA, USA]. The remaining samples were decalcified using 10% EDTA/PBS for 30 days and embedded in paraffin. The tibiae were cut into sagittal serial sections and mounted on glass slides. The slides were then mounted in glycerol/PBS (9:1) containing Hoechst 33258 (2.5 mg/ml; Sigma-Aldrich) to identify the cell nuclei for confocal laser scanning microscopy (CLSM; TCS SP5; Leica).
Statistical Analysis
The results are expressed as means and SDs and were examined by one-way analysis of variance (ANOVA). Data analyses were performed with SPSS software (version 15.0; IBM, Armonk, NY, USA). Tukey's post hoc tests were used for multiple comparisons. In all analyses, p < 0.05 indicated statistical significance.
Results
Effects of Different Concentrations of SDF-1 on MSCs and MSC Sheets In Vitro
Growth curves were plotted to examine cell proliferation (Fig. 2A). After 1–7 days in culture, the MSCs showed time-dependent growth in all the samples. There were no marked differences in cell numbers between MSCs treated with SDF-1 and controls.
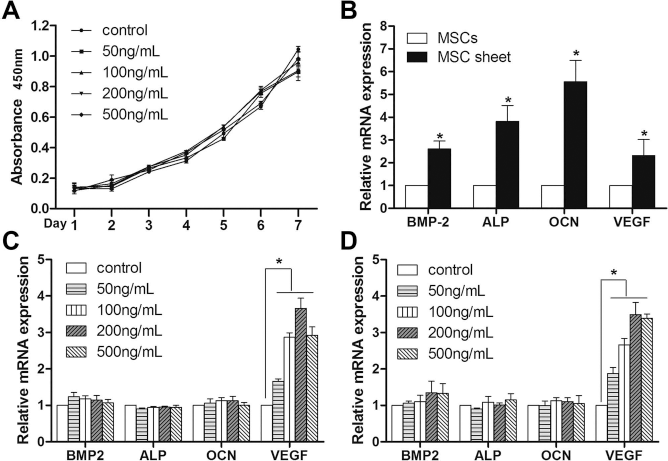
In vitro (A) growth curves indicating the effects of different concentrations of stromal cell-derived factor-1α (SDF-1α) [• 0 ng/ml (control), ▪ 50 ng/ml, ▴ 100 ng/ml, ▾ 200 ng/ml, and ♦ 500 ng/ml] on the proliferation of MSCs. SDF-1α had no significant effect on cell proliferation at any of the different concentrations. (B-D) Real-time quantitative polymerase chain reaction (PCR) data for bone morphogenetic protein 2 (BMP-2), alkaline phosphatase (ALP), osteocalcin (OCN), and vascular endothelial growth factor (VEGF) in vitro. MSC sheet compared with MSCs (B), SDF-1-induced MSCs compared with noninduced MSCs (control) (C), and SDF-1-induced MSC sheet compared with noninduced MSC sheet (control) (D). The groups treated with SDF-1 showed significantly higher expression of VEGF than those of control groups; no statistically significant upregulation occurred in the expression of BMP-2, ALP, or OCN in MSCs or MSC sheets following treatment with SDF-1 (*p < 0.05, compared with controls).
A single MSC sheet from a 60-mm dish had a total cell number of approximately 4 ± 3 × 106 cells. The MSC sheets expressed much higher levels of BMP-2, ALP, OCN, and VEGF than did the cultured MSCs (Fig. 2B). This result indicated that cell sheets have the potential for osteogenesis as well as angiogenesis. Moreover, no significant upregulation was observed in the expression of BMP-2, ALP, or OCN in MSCs (Fig. 2C) or MSC sheets (Fig. 2D) following treatment with 50, 100, 200, or 500 ng/ml of SDF-1. These observations indicate that SDF-1 has no direct effect on the osteogenesis of MSCs or MSC sheets. However, SDF-1 at all concentrations examined had increased expression of VEGF (Fig. 2C and D). In addition, the results showed dosage-dependent effects in all groups, especially at a concentration of 200 ng/ml. These observations suggested that SDF-1 may induce vascularization of MSC sheets.
Secretion of SDF-1 by MSCs
SDF-1 in the MSC supernatant led to a significant increase in the level of SDF-1 in the MSC sheet supernatant compared with the MSC group (174.4 ± 12.3 vs. 23.8 ± 3.6 pg/ml) (Fig. 3).

Concentration of SDF-1 in MSC supernatant. There was a significant increase of SDF-1 in MSC sheet supernatant compared with supernatant of nonseeded MSCs. Results are shown as means ± standard error of the mean (SEM). n = 6 (*p < 0.01, compared with the MSC group).
Effects of SDF-1 on MSC Migration
We performed in vitro chemotaxis assays to examine the migration of MSCs toward exogenous SDF-1α. Compared with the control group, the group with the addition of SDF-1α to the upper chamber of the Transwell system showed a significantly increased number of migratory MSCs (p < 0.05). However, these increases were counteracted by adding SDF-1α at the same concentration to the upper and lower chambers or by pretreatment of MSCs with AMD3100, an antagonist of SDF-1 (p < 0.05) (Fig. 4A).

Effects of SDF-1 on MSC migration. (A) Effects of exogenous SDF-1 on the migration of MSCs. (a) Migratory cells under control conditions (–/–). (b) Migratory cells with SDF-1α added to the lower chamber only (–/SDF-1). (c) Migratory cells with SDF-1α added to both the upper and lower chambers (SDF-1/SDF-1). (d) Migratory cells that had been preexposed to AMD3100 for 30 min and with SDF-1α added to the lower chamber only (AMD3100/SDF-1). The presence of SDF-1 significantly increased the numbers of migratory MSCs compared with the nontreated control group. These increases were offset by adding SDF-1 to the upper chamber at the same concentration or pretreating MSCs with AMD3100. (B) Effects of different concentrations of SDF-1 on the migration of MSCs. (e–h) Migratory cells with 0, 100, 200, and 500 ng/ml SDF-1α added to the lower chamber. With the increase of SDF-1, the number of migrating MSCs increased, especially at a concentration of 200 ng/ml. Abbreviations: MSCs, mesenchymal stem cells; HPF, high-power field; U, upper chamber; D, lower chamber. Scale bar: 50 μm (*p < 0.05; n/s indicates no significant difference).
To investigate whether SDF-1 can induce MSC migration in a dose-dependent manner, the cells were exposed to different concentrations of SDF-1α. The results indicated that the numbers of cells following treatment with SDF-1α at 100, 200, and 500 ng/ml were significantly increased compared with the controls (p < 0.05). However, there was no difference between the 200 ng/ml group and the 500 ng/ml group (p > 0.05) (Fig. 4B).
Gene Expression Profiles In Vivo
This experiment was performed to investigate whether bone fracture regeneration could be related to the expression of ALP, OCN, BMP-2, and VEGF. Significantly elevated levels of OCN, ALP, BMP-2, and VEGF were observed in MSC sheets/SDF at 2 weeks compared with the other five groups (Fig. 5). The SDF and MSC/SDF groups showed upregulation of ALP, BMP-2, OCN, and VEGF expression, which was similar to that observed in the MSC sheet group and significantly higher than that observed in the control and MSC groups. There were no obvious differences between the MSC and control groups.

BMP-2, ALP, OCN, and VEGF gene expression of new bone tissue in different groups 2 weeks after surgery. The group treated with MSC sheet/SDF showed a higher expression of ALP, OCN, BMP-2, and VEGF compared with other groups. SDF, MSC/SDF, and MSC sheet showed a significant upregulation of ALP, BMP-2, OCN, and VEGF expression compared with the MSC and control groups (#p < 0.05, compared with all other groups; *p < 0.05, compared with the MSC and control groups).
Image Analysis
Radiographs taken at 4 weeks showed more callus formation around the fracture site in SDF, MSC/SDF, MSC sheet, and MSC sheet/SDF groups than in control and MSC groups (Fig. 6A–F). The cortical gap was present in all groups except in the MSC sheet/SDF group, and the fracture line was fuzzy and bridging callus formation was observed in the MSC sheet/SDF group (Fig. 6F). The amount of bridging bone formation increased with time in all groups. At 8 weeks after surgery, control and MSC groups showed less callus formation at the fracture site, and the cortical gap was still present, resulting in established nonunion (Fig. 6G and H). In the SDF, MSC/SDF, and MSC sheet groups, more bridging callus formation was seen at the fracture site, and the gap almost disappeared (Fig. 6I–K). In the MSC sheet/SDF group, the cortical gap disappeared, indicating bone union, and showed enhanced consolidation of the fracture site (Fig. 6L).
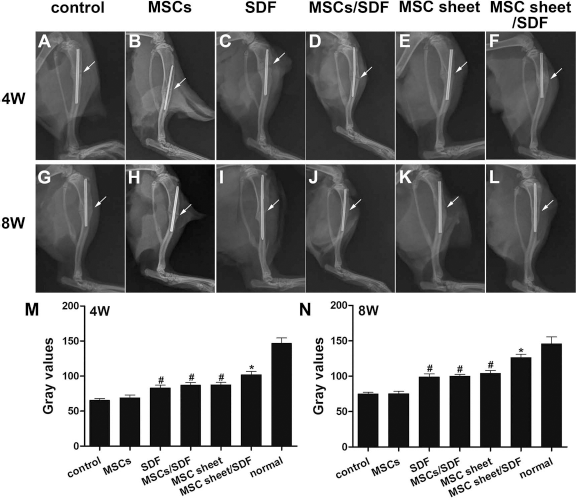
Radiographs and gray values of fracture sites at 4 and 8 weeks after surgery in each group (A–F and G–L, respectively). At 4 weeks, all groups except the MSC sheet/SDF group showed a radiolucent area around the osteotomy site at 4 weeks, and the callus formation was obvious in the MSC sheet/SDF group. At 8 weeks, the control and MSC groups represented a nonunion; faint callus formation was seen at 8 weeks in the SDF, MSC/SDF, and MSC sheet groups; and bone union was achieved in the MSC sheet/SDF group. White arrow indicates the osteotomy site. (M and N) Gray values of each group at 4 and 8 weeks after surgery, respectively. The MSC sheet/SDF group showed a higher gray value compared with the other groups. The gray values in the SDF, MSC/SDF, and MSC sheet groups were significantly higher than those of the control and MSC groups (#p < 0.05, compared with control and MSC groups; *p < 0.05, compared with the other five groups).
The gray values of the fracture sites and healthy tibias from all groups reflected similar results (Fig. 6M and N). The MSC sheet/SDF group showed the highest gray values at both 4 and 8 weeks. However, at 4 weeks, the gray values of the normal tibia were significantly higher than those of the MSC sheet/SDF group. At 8 weeks, the gray values of MSC sheet/SDF group were close to those of the normal controls. The gray values in the SDF, MSC/SDF, and MSC sheet groups were significantly higher than those of the control group and the MSC group. There were no significant differences in gray values for any two groups in the SDF, MSC/SDF, and MSC sheet groups. Additionally, no statistical differences were found between the control group and the MSC group.
After 3D reconstruction, micro-CT images were used to analyze the morphology and cortical continuity of regenerative new bone. The 3D images of all groups differed at both time points. The control group and the MSC group remained primarily separate over the study duration, indicating nonunion (Fig. 7). At 4 weeks, fractures treated with SDF-1, MSC/SDF, or MSC sheets showed significantly reduced cortical fracture gaps, and newly regenerated bone tissue partially bridged the gap (Fig. 7C–E and I–K). The MSC sheet/SDF group had dense calluses, and the osteotomy gap almost disappeared (Fig. 7F and L). At 8 weeks, the SDF, MSC/SDF, and MSC sheet groups showed improved cortical continuity, and the bridging bone was seen in the fracture site (Fig. 7O–Q and U–W). The MSC sheet/SDF group showed complete bone union; the gap disappeared, and the cortical bone was connected (Fig. 7R and X). In terms of BS/TV and BV/TV values, the MSC sheet group revealed a significantly higher value than that in the other groups at both points. Micro-CT also showed a significantly higher value in the SDF, MSC/SDF, and MSC sheet groups compared with the MSC or control group at 4 and 8 weeks. Although the bone quantity parameter in the MSC group was above that in control, no statistical significance existed between them (Fig. 7Y and Z).

(A–X) Micro-CT images of the fracture sites in each group at 4 or 8 weeks after surgery (A–L and M–X, respectively). (Y, Z) Statistical analysis of BV/TV (%) and BS/TV (mm−1). More hard tissues formed in the MSC sheet/SDF group than in the other groups at all time points (A–X). MSC sheet/SDF increased the bone quantity at all time points (Y, Z). White arrow indicates the osteotomy site (*p < 0.05, compared with other groups; #p < 0.05, compared with the MSC group and controls).
Histological and IHC Assessment
Histological analysis at 4 weeks after implantation indicated no bridging bone formation at the fracture site in the control and MSC groups. Although newly formed bone tissue was observed around the fracture sites in the SDF, MSC/SDF, and MSC sheet groups, the cortical fracture gaps were still present in the SDF, MSC/SDF, and MSC sheet groups. In addition, the regenerative tissues in the SDF, MSC/SDF, and MSC sheet groups were filled with chondrocytes and fibrous tissue. Newly formed woven bone tissue united both the distal and proximal sites in the MSC sheet/SDF group (Fig. 8A, 4W). At 8 weeks, there was no continuous bone tissue at the fracture site in the control and MSC groups. These gaps resulted in nonunion of the tibia. Newly formed woven bone tissue was observed in the SDF, MSC/SDF, and MSC sheet groups. However, in the MSC sheet group, some cancellous bones were replaced by cortical bone. In contrast, the MSC sheet/SDF group showed complete bridging of the fracture site, indicating the formation of hard bone and complete bony union (Fig. 8A, 8W). The newly formed bone area in the MSC sheet/SDF group was significantly higher than that of the other five groups at both 4 and 8 weeks. The new bone tissue area in the MSC sheet, MSC/SDF, and SDF groups was significantly larger in the MSC and control groups at 4 and 8 weeks. However, no significant difference was found for any two groups among the MSC sheet, MSC/SDF, and SDF groups at 4 and 8 weeks (Fig. 8B).

Histological analysis of fracture sites in each group 4 and 8 weeks after surgery. (A) Hematoxylin and eosin (H&E) staining and Safranin-O staining (w: woven bone, f: fibrous tissue, and arrow: chondrocytes). At 4 and 8 weeks postsurgery, remarkably more regenerated new bones had formed in the MSC sheet/SDF group than in the other five groups. Nonunion occurred in the MSC and control groups. (B) Comparison of newly formed bone areas in each group. The results of percentage of the newly formed bone area were significantly higher in the MSC sheet group than those in the other five groups at 4 and 8 weeks. A similar result was shown in the MSC sheet, MSC/SDF, and SDF groups compared with the MSC and control groups at both points (*p < 0.05, compared with the other groups; #p < 0.05, compared with the MSC and control groups; scale bars: 1 mm).
IHC staining was performed for OPN and BSP, which are osteogenic markers for bone regeneration. The results showed increased matrix deposition in new bone tissue and active bone remodeling in the fracture site. Although groups with implanted MSC sheets or injection of SDF-1 showed positive staining for OPN and BSP, the staining was significantly more intense and covered a larger area in the MSC sheet/SDF group (Fig. 9A). A similar result was shown in the OD value. The protein expressions of BSP and OPN in the MSC sheet/SDF group were much higher than that in other groups. In addition, the expression of bone-related markers in the MSC sheet, MSC/SDF, and SDF groups was significantly increased compared to that in the MSC and control groups (Fig. 9B).
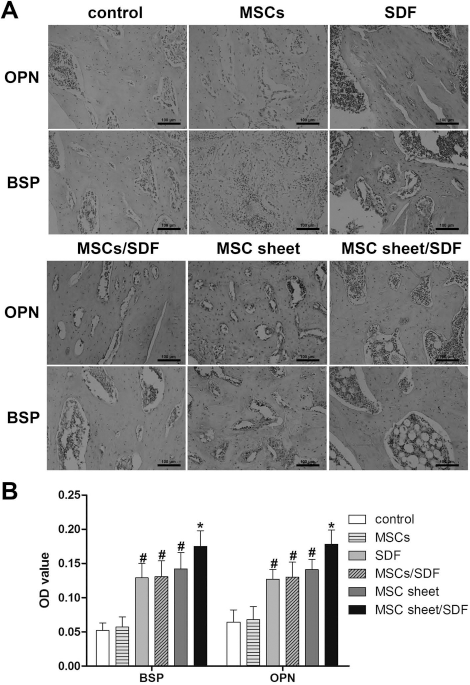
(A) Immunohistochemical staining for osteogenic markers in each group 4 weeks after surgery. OPN, osteopontin; BSP, bone sialoprotein. At 4 weeks, immunoreactivity of BSP and OPN was discernible and there was no significant difference between the MSC and control groups. (B) Quantitative analysis of bone-related markers in each group. SDF, MSC/SDF, MSC sheet, and MSC sheet/SDF groups showed stronger immunoreactivity than the other two groups, but a significantly higher optical density (OD) value of BSP and OPN staining was detected in the MSC sheet/SDF group than in the others (*p < 0.05, compared with the other groups; #p < 0.05, compared with the MSC and control groups; scale bars: 100 μm).
IHC staining for CD34 showed that markedly more CD34+ vascular ECs were present in the MSC sheet/SDF group than in the other five groups. Moreover, the density of blood vessels in regenerative tissue treated with the MSC sheet, MSC/SDF, or SDF was significantly higher than in MSC and control groups (Fig. 10).

(A) Immunohistochemical staining images of CD34+ vascular endothelial cells in each group after 4 weeks (arrow: CD34+ cells). (B) Vascular density within regenerated tissue 4 weeks after treatment in each group. There were greater numbers of CD34+ vascular endothelial cells in the MSC sheet/SDF group than in the other groups (*p < 0.05, compared with other groups; #p < 0.05, compared with the MSC and control groups; scale bar: 100 μm).
Detection of Donor Cells
The transplanted GFP-MSCs were easily identified within the operative site by their green fluorescence at both 2 and 8 weeks postoperatively (Fig. 11). These observations indicated that the cells from cell sheets were present in the regenerating tissue at least 8 weeks after transplantation and that the donor cells proliferated, differentiated, and finally formed bony connections.

Tracking of donor cells in vivo. Fluorescence images obtained with a small animal in vivo fluorescence imaging system (A, C) and confocal microscopy (B, D). Green fluorescent protein (GFP)-MSCs were identified by green fluorescence in the operative site [scale bars: 75 μm (B, D); arrows: green fluorescence area].
Discussion
We demonstrated that MSC sheets transplanted into bone fractures along with local injection of SDF-1 greatly promoted fracture regeneration. The results confirmed the following: (1) MSC sheets can be produced easily from MSCs isolated from bone marrow and can improve bone healing; (2) the SDF-1/CXCR4 chemokine axis plays an important role in the migration of MSCs in vitro and promotes the trafficking of MSCs to sites of bone injury in vivo; and (3) injection of SDF-1 affects the osteogenesis and angiogenesis of MSC sheets. In summary, these observations suggest a new clinical strategy for treating fractures and delayed bone unions or bone nonunions.
MSC Sheets Represent an Ideal Means of Achieving Scaffold-Free Bone Formation
Our results showed that transplanted MSC sheets significantly enhanced bone regeneration in all groups compared with controls. Importantly, the availability of adequate cells was found to promote the repair process. We successfully incorporated about 7 × 106 MSCs into 60-mm culture dishes, and the sheets thus obtained were transplanted directly into the lesions. It is difficult to achieve this cell density with other techniques. In addition, both the adhesion molecules on the cell surface and the interactions among cells remained intact, resulting in good MSC survival and proliferation 12 . This effect in maintaining cell viability may be partly explained by the fact that although we implanted with the same quantity of MSCs into the fractural area in the MSC and MSC/SDF groups, more newly formed bone was observed in the MSC sheet and MSC sheet/SDF groups. In addition, the present study revealed significantly more SDF-1 in conditioned media obtained from MSC sheet culture supernatant than in that from MSCs, which is consistent with the suggestion that SDF-1 secretion occurs only when direct contact between bone marrow-derived stromal cells (BM-MSCs) is possible 26 . SDF-1 is known for its effect on the recruitment of CXCR4+ cells. This recruitment may increase the number of cells participating in the healing process. Furthermore, neovascularization at the fracture site is also as a key factor in fracture healing. The hypoxia that occurs after a fracture regulates the microenvironment, including the expression of VEGF 27 . Ito et al. reported that VEGF is critical for the reconstitution of the cortical surface of autografts and can induce a new vascular network around the fracture, thus leading to bone regeneration 28 . According to our results, the MSC sheets showed higher levels of VEGF and osteogenic gene mRNA expression (BMP-2, ALP, and OCN) than did the MSCs. Therefore, we concluded that the MSC sheets secreted VEGF around the bone lesions, which subsequently induced neovascularization and bone regeneration. These characteristics suggested that the MSC sheet functions similarly to fresh periosteum, which has been shown to efficiently enhance repair of bone defects 29 . This was further confirmed by the results of gene expression, X-ray, micro-CT, histological, and IHC analyses of the MSC sheet group in vivo. Therefore, the results of the present study, along with those reported previously30,31, confirm that MSC sheets can promote bone healing.
Local Injection of SDF-1 Enhances Regeneration of Bone Fractures
After 4 and 8 weeks, rats injected with SDF-1 showed accelerated new bone formation compared with the control group. This may have been due to the local increment in the SDF-1 level related to in vivo injection, which facilitated homing to the fracture. Several groups reported that local application of exogenous SDF-1 promotes the recruitment of endogenous progenitor cells and bone formation24,32. We assessed the chemotactic capacity of MSCs in vitro using a Transwell system. The number of migrating MSCs was increased by SDF-1 in a dose-dependent manner. However, migration was inhibited by an anti-CXCR4 antibody, indicating that MSCs express functionally active CXCR4 receptors, which are responsible for their migration to sites of injury. A similar study conducted by He et al. indicated that SDF-1-induced migration of MSC was dose dependent in the concentration range of 50 to 250 ng/ml 33 . Meanwhile, taking the SDF-1 secreted from MSC sheets into consideration, we postulated that the optimum concentration of SDF-1α injected into the lesion area would be 200 ng/ml. As higher dosages of SDF-1 have been confirmed to be critical for the mobilization of MSCs from the bone marrow 34 , this burst increase is advantageous. The SDF-1/CXCR4 axis also works in ECs, smooth muscle cells (SMCs), and EPCs35–37 and improves angiogenesis both in vitro and in vivo. It was confirmed in our research that more CD34+ vascular ECs were observed in the groups that were injected with SDF. These cells homing to the lesion site could promote the construction of the vessels in vivo. Moreover, our data also showed higher levels of VEGF mRNA expression in MSCs treated with SDF-1α, which confirmed the results reported previously by Pasha et al. 38 . There may be some association between SDF-1 and VEGF signaling pathways. We speculate that the increased secretion of VEGF could improve the vascularization of the MSCs. However, our results indicated that SDF-1 did not markedly promote the proliferation or osteogenesis of MSCs in vitro. Liu et al. 39 and Nair et al. 40 reported similar results indicating that SDF-1α did not seem to have a major influence on the osteogenic differentiation of MSCs. These observations indicate that SDF-1 has no direct effect on the osteogenesis of MSCs. Furthermore, Jin and Giannobile 41 demonstrated that SDF-1 did not impact de novo bone formation, and it plays a role in the repair process through circulating stem cell recruitment and angiogenesis.
Taken together with these previous reports, our results suggest that SDF-1 influenced the process by which bone fractures are repaired by increasing the number of these cells homing to the lesion and stimulating the angiogenesis of cells rather through its direct effects on the behaviors of host cells, primarily MSCs. The implanted MSC sheets may play a critical role in cellular differentiation.
SDF-1 Combined with MSC Sheets Greatly Promoted Bone Regeneration
Owing to the positive effect of SDF and the MSC sheets on osteogenesis and vasculogenesis, our results showed that the combination of SDF with an MSC sheet induced the strongest bone regenerative response and greatly accelerated bone formation compared with groups treated with MSC sheets or SDF alone. The MSC sheet/SDF group had thick cortical bone tissue and complete bone union. This is the first report indicating that effective transplantation of an MSC sheet with injected SDF -1 has effects on treating bone nonunion.
Although SDF-1 had no direct effect on osteogenesis, our results indicate that, relative to the other groups, new bone formation was significantly enhanced in the group in which MSC sheets were transplanted in combination with SDF-1 injection. This was probably due to the higher expression of BMP-2 in the MSC sheet associated with SDF-1 injection. The SDF-1/CXCR4 axis may regulate a BMP signaling pathway and promote the early osteogenic differentiation of MSCs. BMP-2 is known to be an active osteogenic regulator, which induces orthotopic and ectopic bone formation, and is also known to enhance cell chemotaxis and vasculogenesis42,43. Some groups have confirmed that SDF-1 meditates the osteogenic differentiation of MSCs induced by BMP-2 through the regulation of intracellular Smad and Erk activation44,45. Second, SDF-1 can recruit EPCs and enhance vascularization in vivo. Furthermore, previous studies have indicated that SDF-1 and VEGF have a reciprocal relationship in terms of secretion38,46. It has been confirmed that VEGF is upregulated in the initial stages of bone regeneration. This can enhance bone healing, including improving vasculogenesis, and can result in the conversion of soft cartilaginous callus tissue to hard, bony callus47–49. This may explain why, although no cells were transplanted into the site of injury, increased newly formed bone was also seen at the fracture site in the SDF-1 group. Third, our tracking experiments indicated that MSCs from MSC sheets survived for at least 8 weeks in vivo. In addition, Kortesidis et al. reported that high SDF-1-expressing BM-MSCs increased cellular growth and protected BM-MSCs from interleukin-4 (IL-4)-induced apoptosis 50 . These studies confirmed that SDF-1 could improve the maintenance, survival, and development of MSCs. Taken together, the above points and the results of the present study suggest that our technique of MSC sheet transplantation combined with local injection of SDF-1 greatly enhances bone formation.
It should be noted that this study has some limitations. First, SDF-1 was simply injected around the implantation site. In future studies, we plan to investigate the trend of the SDF-1 concentration in plasma and in the microenvironment around the injury site. This would allow more appropriate treatment via the injection of SDF-1 at different time points. Furthermore, we will also introduce an SDF-1 release system to maintain the SDF-1 secretion at the optimal effective concentration around the fractured bone. Second, the precise mechanisms underlying the cooperative effect of SDF-1 and the MSC sheet should be investigated further based on the present study.
Conclusions
In the present study, both SDF-1 and the MSC sheet contributed to bone regeneration. MSC sheet transplantation combined with the local injection of SDF-1 around the implantation site greatly promoted bone healing. This technique will facilitate the development of strategies for treatment of fracture or delayed bone union or bone nonunion in clinical settings.
Footnotes
Acknowledgments
This work was supported by the Natural Science Foundation of China (Nos. 81401779, 81271356, and 81201408), the Natural Science Foundation of Zhejiang Province (No. LQ14H060001), the Science Technology Program of Zhejiang Province (Nos. 2013C33096 and 2012C23104), the TCM Foundation for Distinguished Young Talents of Zhejiang Province (No. 2012ZQ024), the Key Medical Disease Program of Hangzhou city (No. 20120533Q39), and the Key Science Technology Program of Xiaoshan district (No. 2012234). The authors declare no conflicts of interests.