
Abstract
Spinal cord injury (SCI) can cause severe traumatic injury to the central nervous system (CNS). Current therapeutic effects achieved for SCI in clinical medicine show that there is still a long way to go to reach the desired goal of full or significant functional recovery. In basic medical research, however, cell transplantation, gene therapy, application of cytokines, and biomaterial scaffolds have been widely used and investigated as treatments for SCI. All of these strategies when used separately would help rebuild, to some extent, the neural circuits in the lesion area of the spinal cord. In light of this, it is generally accepted that a combined treatment may be a more effective strategy. This review focuses primarily on our recent series of work on transplantation of Schwann cells and adult stem cells, and transplantation of stem cell-derived neural network scaffolds with functional synapses. Arising from this, an artificial neural network (an exogenous neuronal relay) has been designed and fabricated by us—a biomaterial scaffold implanted with Schwann cells modified by the neurotrophin-3 (NT-3) gene and adult stem cells modified with the TrkC (receptor of NT-3) gene. More importantly, experimental evidence suggests that the novel artificial network can integrate with the host tissue and serve as an exogenous neuronal relay for signal transfer and functional improvement of SCI.
Keywords
Introduction
Following spinal cord injury (SCI), it is inevitable that numerous ascending and descending nerve fiber tracts are lesioned causing partial or total disruption of sensory/motor neural circuits below the injured site in the spinal cord. In view of this, attempts to promote the regeneration of nerve fiber tracts across the injured site, reconstruction of synaptic connections, bridging the axotomized neuronal fibers, and remolding the neuronal circuits by transplanting nerve tissues or stem cell-derived neurons have become the major strategies to repair the injured spinal cord. In the past two decades, progress has been made in basic and clinical research in SCI, and arising from this, many therapeutic strategies have been designed and developed for the treatment of SCI1,2. In this review, we will highlight the effectiveness of a novel therapeutic strategy for SCI by transplantation of combined biomaterial constructs seeded with the neurotrophin-3 (NT-3) gene-modified Schwann cells (SCs) together with adult stem cells, retinoic acid (RA)-induced stem cells, or NT-3 receptor TrkC gene-modified stem cells.
Transplantation of SCs for Treatment of Sci
SCs, myelinating cells of the peripheral nervous system, are capable of secreting various neurotrophic factors (NTFs), such as NT-3, nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), ciliary neurotrophic factor (CNTF), glial cell-derived neurotrophic factor (GDNF), basic fibroblast growth factor (bFGF), etc. These NTFs have been shown to promote survival of injured neurons and axonal regeneration. SCs can also produce extracellular matrix-like laminin, fibronectin, integrin, and cell adhesion molecules, such as neural cell adhesion molecule (NCAM) 3 , all of which play a role in axon growth. Additionally, SCs synthesize interleukin-6 (IL-6), which promotes neuronal survival, and growth associated protein-43 (GAP-43), which induces regeneration and sprouting of injured axons.
Because of these biological characteristics, SCs are deemed essential to creating favorable microenvironments for peripheral nerve repair. The limited self-repair capability of the central nervous system (CNS) may be attributed to its lack of SCs. Di Giovanni transplanted injured neurons from the spinal cord lesion site into a peripheral nerve environment 4 . Remarkably, it was found that the injured neurons could completely recover like normal neurons in terms of electrophysiological and external morphological features 4 . It was reported that when autologous sciatic nerve segments were grafted into the injury site of the spinal cord, some axons originated from intrinsic CNS neurons and extended into the grafts, but no obvious functional improvement was observed 5 . Although endogenous SCs can translocate and migrate into the injured spinal cord and contribute to self-repair, the effect is regarded to be rather insignificant6,7. Notwithstanding, SCs are undoubtedly the most widely studied cell type in designing a cell transplantation therapy for SCI. There is mounting evidence indicating that grafted SCs could promote axonal regeneration and remyelination8–11. Nerves with segmental demyelination often develop significant secondary axonal loss 12 . Compared with other cells, the advantage of SCs lies in wrapping the regenerating axons in myelin sheath13,14. Remyelination helps axonal survival and signal conduction 15 . It has been reported that axonal signals to the SCs dictate which axons are to be myelinated and how much myelin is to be produced 16 . Meanwhile, the axon–SC interaction provides guides for axonal regeneration17,18. We investigated the effects of collagen or gelfoam seeded with SCs on the structural and functional recovery of completely transected spinal cord 19 . Three months after transplantation, the voluntary locomotor function of hindlimbs was partially restored. Some neurons in the sensorimotor area of the cerebrum and the red nucleus (RN) of the midbrain were shown to be retrogradely labeled with fluorogold (FG), suggesting that their axons had regenerated through the injured site and into the caudal spinal cord. Associated with the above finding was the occurrence of calcitonin gene-related protein (CGRP)- and 5-hydroxytryptamine (5-HT)-positive nerve fibers in the injury/graft sites. Moreover, 5-HT-positive nerve fibers were found caudal to the injury/graft sites, suggesting that some descending fibers had regenerated and crossed the injury/graft site of the spinal cord 19 . In addition, transplanted SCs may promote neuronal survival in the sensorimotor area of the cerebrum and the RN of the midbrain 20 . The survival of these neurons is necessary for axonal regeneration 21 .
SCI can lead to a series of pathophysiological changes. Results from our studies have shown that combined treatment may be a more effective therapy for SCI, the reason being the outcome of any single therapy has not been satisfactory. Though transplanted SCs alone can protect the injured neurons and promote axonal regeneration, they could not migrate into the host tissue; hence, regeneration outside of the injury/graft site of spinal cord was limited. Glial scarring, axonal growth inhibitors, and myelin-associated inhibitors have been implicated in diminishing the intrinsic regenerative capacity when SCs were grafted alone 7 . It is therefore desirable to identify new targets that may be coupled with SC transplantation to facilitate new nerve fiber regeneration22,23.
In our studies, we grafted SCs into the hemisection site of the spinal cord. Both the numbers of surviving neurons and nitric oxide synthase (NOS)-positive neurons in the Clarke's nucleus (CN) on an injured side were increased; on the other hand, shrinkage of soma areas in neurons remained evident 24 . L-Arginine can form nitric oxide (NO) with NOS. It is not certain whether NO derived from L-ariginine via NOS had any effect on the survival of injured neurons in CN. N-Methyl-D-aspartate (NMDA) receptor antagonist (MK801) and SCs were then applied together to observe their effect on the survival of CN neurons and expression of NOS. It was found that after combining SCs with MK801, the survival of injured CN neurons was greatly improved; moreover, shrinkage of neuronal somas became less evident. Furthermore, NOS activity in surviving neurons was decreased, and the shrinkage of their somas was prevented in the SC + MK801 group. The results indicate that the combination of SCs and MK801 can promote the survival of CN neurons and prevent the shrinkage of CN and its neuronal soma, which may be due to the decrease of NOS activity 25 .
Transplantation of Neural Stem Cells for Treatment of Sci
Neural stem cells (NSCs) are multipotent self-renewing stem cells that can differentiate into the main phenotypes of CNS cells (neurons, astrocytes, and oligodendrocytes) in vitro and in vivo26,27. NSCs were first identified in the subventricular zone (SVZ) of mice in 1989 26 and were isolated from the mouse striatal tissue and SVZ for the first time in 199228,29. The major function of NSCs is replacing apoptotic cells and repairing the injured nerve tissues. The NSCs' capability of self-renewal and differentiation has made them potential candidate cells for treating SCI.
We seeded NSCs in a type I collagen scaffold and grafted them into the spinal cord that had undergone a total transection at the T10 level in rats 30 . Sixty days after transplantation, the results showed the following. (1) The grafted NSCs could survive and migrate into the injured spinal cord tissue, wherein some of them differentiated into glial fibrillary acidic protein (GFAP)-, neurofilament-200 (NF-200)-, and GAP-43-positive cells. GAP-43 is a unique phosphorylated glycoprotein in neurons and plays a key role in neurogenesis, development, axonal regeneration, and synapse formation. This suggests that NSC-derived neuron-like cells can form long extending processes. (2) In the NSC-transplanted group, some neuronal somata in the RN and the inner pyramidal layer of the sensory and motor cortex were retrogradely labeled by the fluorescent tracer FG. (3) Degenerating features of the injured spinal cord were diminished. (4) Locomotor function of the hindlimbs in the rats with injured spinal cords was improved. The results demonstrate that grafted NSCs can differentiate into neurons and glial cells, which may alleviate secondary pathogenic damage, preserve injured neurons, and promote the recovery of locomotor function in rats with totally transected spinal cords 30 .
It was then surmised that a major mechanism of NSC therapy for SCI would be replacement of the injured or dead neurons, so that the function of signal transfer is restored. On the other hand, while locomotor function of hindlimbs is partly recovered, NSCs transplanted alone had a rather low rate of differentiation into neurons. Grafted NSCs differentiate preferentially into glial cells, especially astrocytes 31 . The limited survival of the implanted cells and the low rate of neuronal differentiation of NSCs are two major challenges for their use in cell transplantation therapy. In light of this, other strategies including combined transplantation of two types of cells, application of NTFs, gene therapy, predifferentiation, or biomaterial scaffolds to promote neural differentiation, and synaptic formation have been considered and systematically explored.
Combination of SCs and NSCs
Although some NTFs, such as BDNF, GDNF, NT-3, and bFGF, have been demonstrated to promote the survival of NSCs and induce the differentiation of precursor cells into neurons in vitro 32 , the effect of application of these factors on NSCs in vivo is not clear. More importantly, since many of these factors have a relatively short half-life in vivo, it is difficult to maintain a long-term effect on transplanted NSCs in vivo. Although there are no data available on how long SCs can survive after transplantation, it has been reported that transplanted SCs can survive in the host spinal cord for at least a few months13,33. Because SCs are capable of secreting various kinds of NTFs, it was speculated that if SCs and NSCs were to be cotransplanted into an injured spinal cord, more NSCs would differentiate into neurons to replenish the loss. By the same logic, the grafted cells would act as a bridge and rebuild the neural pathway, and promote neural regeneration and ultimately functional recovery. On the basis of this assumption, we cocultured SCs and NSCs in vitro to explore how SCs might influence the survival and differentiation of NSCs 34 . The results showed that the number of surviving NSCs and differentiated neuron-like cells was significantly increased. The cytoplasmic processes extending from the neuron-like cells were obviously longer than the control group. Cocultured SCs and NSCs have grown to make physical contacts with each other in at least three different configurations: (1) cell body-to-cell body contacts, (2) cell body contacting cytoplasmic process, and (3) contacts between cell processes 34 . These features support that SCs could promote the survival and differentiation of NSCs. We next cografted SCs and NSCs into the hemisected site of the spinal cord. At 30 days after surgery, the number of surviving NSCs and differentiated neuron-like cells was significantly increased in the cografted group, while the frequency of GFAP-positive cells decreased, when compared with the control group (NSCs only) 35 . Some grafted NSCs exhibited long neurites, and some of them expressed acetylcholinesterase. It is noteworthy that some of the grafted NSCs were distributed in the white matter of the spinal cord, where they differentiated into myelin-forming cells. Taken together, the results indicate that the cografted SCs could increase the survival rate of NSCs and promote their differentiation into neuron-like cells 35 .
Cotransplantation of NTF Gene-Modified SCs and NSCs
Apart from causing the neuronal death, axotomy, demyelination, and injury to the spinal cord also activate the secretory activity of astrocytes, causing production cytokines that inhibit nerve regeneration and form glial scars 36 . Moreover, the injury site often lacks sufficient cytokines necessary for promoting nerve regeneration, such as NTFs. Therefore, enhancing the local NTF levels would conceivably provide an environment that may offer necessary protection for damaged neurons from the effects of secondary injury and, hence, be conducive for axonal regeneration. NTFs are a group of proteins that regulate the survival of neurons, axonal growth, synaptic plasticity, and synthesis of neurotransmitters37,38, and these include neurotrophins, fibroblast growth factor (FGF), insulin-like growth factor (IGF), transforming growth factor (TGF), etc. Neurotrophins act by binding to two kinds of membrane receptors: p75 neurotrophin receptor (p75NTR) and tyrosine kinase receptors (Trk). p75NTR regulates three major signaling pathways: NF-κB, Jun kinase, and Rho. The binding of neurotrophins and p75NTR can promote the extension of neurites, but in the absence of neurotrophins the growth cone collapses39,40. Additionally, the p75NTR regulates apoptosis and cell death 41 . We have reported that NT-3 improves the neural differentiation of skin-derived precursors induced by RA through p75NTR-dependent signaling pathways2,42. The Trk family includes three subtypes: TrkA, TrkB, and TrkC. The major neurotrophins include the following: NGF, which binds with TrkA; BDNF and NT-4/5, which bind with TrkB; and NT-3, which primarily binds with TrkC. NT-3 also binds to TrkB and TrkA, but with a lower affinity. In most neurons, the interaction between neurotrophins and Trk receptors increases the survival of neurons43–45. It is generally accepted that NGF mainly affects the sensory neurons, while BDNF affects a small range of neurons. It is well documented that NT-3 plays an important role in not only the development and differentiation of neurons but also the survival of injured neurons and axon regeneration 46 . NT-3 has been shown to influence the gene expression of microtubule-associated protein 2 (MAP2), which is thought to be involved in active axonal outgrowth during the growth of the nervous system47,48. After SCI, the corticospinal tract (CST) axons hardly regenerate, but these axons exhibit sprouting in response to NT-349–51. Fortun et al. used adeno-associated virus (AAV) vectors for delivery of the NT-3 gene. Following injection into the triceps of the SCI animals, a significant increase in NT-3 levels was detected in the cervical enlargement of the spinal cord. Interestingly, only moderate astrogliosis and less CST axonal retraction and/or enhanced sprouting of CST axons were found. It was reported that the delivery of NT-3 via retrograde transport of AAV from triceps to cervical motor neurons had led to reduced functional loss and anatomical reorganization of the CST following injury 52 . Remarkably, other trophic factors, such as IGF, failed to induce corticospinal fiber regeneration after SCI 53 . Application of NT-3 can also prevent the atrophy of neurons in the CNS and promote axonal regeneration and hindlimb functional recovery in SCI rats54–57. Other studies had shown that NT-3 regulated the differentiation of NSCs into neurons through extracellular signal-regulated kinase (ERK) signaling pathway. Inhibition of ERK resulted in a significant decrease in the number of NSCs that differentiated into neurons 58 . In a separate in vitro study, it was reported that BDNF and NT-3 act as axonal guidance factors during growth of the spinal neurons 59 . At present, there are two major routes whereby NTFs are administered into the CNS. This is achieved through direct injection of exogenous NTFs or gene therapy by delivering NTF genes into the injured CNS. Since NTFs have a short half-life in vivo and are hardly able to cross the blood–brain barrier (BBB), the effect of exogenous NTFs cannot be sustained in the long term. We have, therefore, opted for NT-3 gene therapy. For this, we transduced the NT-3 gene into SCs, so that they overexpressed NT-3 along with other cytokines. In vitro, more NSCs differentiated into neuron-like cells when cocultured with the genetically modified SCs 60 . On the basis of this approach, we cografted NT-3-transduced SCs (hereafter referred to as NT-3-SCs) and NSCs into SCI rats whose spinal cords were transected, and a segment 2 mm in length between T9 and T10 was removed 61 . Two months after surgery, results showed that (1) some NSCs could differentiate into neuron-like cells at the injured site of the spinal cord, and the highest percentage of differentiation occurred in the NT-3-SC + NSC group. (2) In the grafted group, there were 5-HT-, CGRP-, and substance P (SP)-positive nerve fibers within the injured site. Some FG-labeled cells were found rostral to the injured site of the spinal cord, the RN, and the inner pyramidal layer of the sensorimotor cortex (SMC) of the cerebrum. (3) The grafted cells could enhance the survival of injured neurons in the inner pyramidal layer of the SMC, RN, and CN of the spinal cord. (4) The grafted cells could decrease the latency and increase the amplitude of cortical somatosensory evoked potential (CSEP) and cortical motor evoked potential (CMEP), which showed that they could partly promote structural and functional recovery of SCI rats 61 . It can be concluded that all of this was attributed to the graft of overexpressing NT-3-SCs, which therefore promote the survival and differentiation of NSCs, axonal regeneration, and functional improvement.
Cotransplantation of Genetically Modified NTF-Overexpressing SCs and NSCs Induced by RA
Since the microenvironment of the injured site of the spinal cord is unfavorable for the differentiation of NSCs, predifferentiation of these cells in vitro has been considered an option to increase the rate of differentiation along with cografting with genetically modified NTF-overexpressing SCs. RA, a vitamin A derivative, has various biological effects on growth, development, metabolism, cellular survival, and differentiation. It is also a strong inducing agent as it exerts a synergistic effect with NTFs on neurogenesis and facilitates the neuronal differentiation of stem cells62–64. Although both NTFs and RA play an important role in early neurogenesis, their mechanisms are clearly distinct. The effect of NTFs is usually mediated via membrane receptors, while RA acts by binding with the nuclear receptors, RA receptors (RARs).
We reported that RA in the presence of SCs exerted a robust effect on neuronal differentiation and synaptogenesis of NSCs in vitro 65 . We next cotransplanted NSCs pretreated with RA and NT-3-SCs into the injury/graft site of SCI rats. Results showed that there were significantly more surviving neurons in the SMC, RN, and CN, axonal regeneration, and remyelination. Remarkably, some 5-HT nerve fibers had regenerated through the injury/graft site to reach the cord caudal to the injury/graft site. Apart from glial-like cells, more neuron-like cells were observed in the injury/graft site. Additionally, PSD95 (postsynaptic density 95)-, synapsin-, and nestin-positive cells were detected, suggesting formation of synaptic contacts between the grafted cells. Furthermore, BBB scores were significantly improved. By electron microscopy, regenerating nerve fibers were found to be remyelinated in the injury/graft site of the spinal cord 65 . These results indicate that cotransplantation of NT-3-SCs and RA-preinduced NSCs may mediate partial improvement for SCI, both functionally and structurally.
Cotransplantation of Genetically Modified NTF-Overexpressing SCs and Genetically Modified Receptor-Expressing NSCs
Binding of neurotrophins to Trk results in receptor dimerization by activating the intrinsic tyrosine activity of Trk, and this results in autophosphorylation of the receptor and triggers phosphorylation of its downstream substrate, thereby activating various target signal molecules. Even in the absence neurotrophins, Trk in high density is triggered by interactions with each other, which leads to autophosphorylation 66 . Each Trk receptor has been reported to control three major signaling pathways: PI3K/Akt, mitogen-activated protein kinase (MAPK), and phospholipase C gamma (PLC-γ)67–70. Axonal growth is linked to assembly and disassembly of actin filaments. Binding of neurotrophins to Trk results in axonal extension by actin depolymerizing factor (ADF)/cofilin regulation in a signaling pathway that decreases phosphorylation of these actin-binding proteins in order to stimulate neurite growth71,72. Our study has shown the beneficial effect of cografted NSCs and NT-3-SCs over NSCs and unmodified SCs. Research in vitro showed that NT-3 could increase the expression of TrkC in NSCs so that more NSCs would differentiate into neurons60,73. Castellanos et al. found that the survival rate of genetically modified TrkC-NSCs could be up to 100% with application of NT-3 74 . Conversely, a lack of NT-3 or TrkC could cause the survival rate to decrease by 30–50%. The combined treatment reduced the differentiation of transplanted cells into glial cells.
The results when taken together indicate that applying both NT-3 and its high-affinity receptor TrkC may be the strategy of choice for the effective treatment of SCI. In order to cograft SCs and NSCs with genetically enhanced expression of NT-3 and TrkC (NT-3-SCs and TrkC-NSCs), we have constructed recombinant adenoviral vectors carrying the human NT-3 or TrkC gene (Ad-NT-3/Ad-TrkC) in the hope that NT-3-SCs would promote the survival and differentiation of TrkC-NSCs. First, we cocultured NT-3-SCs and TrkC-NSCs in vitro and found that NSCs showed the tendency to become neuron-like cells compared to the control. PSD95 and synapsin were detected, indicating de novo formation of synapses among the differentiated neuron-like cells. We next grafted a piece of gelfoam preloaded with these gene-modified cells into the lesion gap at the T10 segment of the spinal cord that was transected. Two months later, results showed (1) a significantly improved relay of CMEP and CSEP. Strikingly, the hindlimb deficits were ameliorated. (2) Increased levels of neuronal differentiation of NSCs, synaptogenesis (including inhibitory/type II-like synapses), and myelin formation of SCs, in addition to neuroprotection and outgrowth of 5-HT nerve fibers in the lesioned spinal cord, were observed. (3) Elevated expression of laminin, a proneurogenic factor, and decreased presence of chondroitin sulfate proteoglycans (CSPGs), major inhibitors of axonal growth and neuroplasticity, were detected 75 . The results have convincingly demonstrated that the interaction between NT-3 and TrkC plays a key role in improving the therapeutic effect. It is tempting to speculate that the axonal regeneration and neuronal differentiation following combined treatment with NT-3 and TrkC would ultimately lead to a reorganization of the intraspinal cord neural circuitry. This would provide the underlying morphological basis for consideration of designing therapeutic strategies for SCI 75 .
Transplantation of Neural Network by Tissue Engineering
In current surgical interventions for treatment of SCI, it is well recognized that there is poor, if any, regeneration of axons through the injury epicenter from the supraspinal motor neural pathways, including those that emanate from the motor cortex (such as the CST), midbrain (such as the rubrospinal tract), or brain stem (such as the reticulospinal tract). There is an apparent lack of anatomical evidence demonstrating the existence of reestablished neural circuits; furthermore, the functional improvement is hardly encouraging 76 . In severe injury that causes large cavitation, the physical barrier may completely impede the potential intrinsic nerve regeneration. Therefore, the use of a neuroengineering approach would be a promising therapy and, indeed, an option to overcome the above-mentioned obstacle in SCI repair. This takes into consideration some of the advantages of this strategy, i.e., a biological function-oriented engineering endeavor involving the combination and interaction of cells, bioactive molecules, and biological scaffolds. The lesion gap/cavity is located in the injury epicenter of the spinal cord. Buoyed by the recent anatomical evidence that the axons of propriospinal neurons (PSNs) are able to regenerate, it was surmised that spontaneous bridge formation at the lesion site might be capable of forming new intraspinal circuits77–79. Stem cell transplantation has been utilized to form interneurons that can serve as a bridge to restore communication between the brain and remote spinal cord80–84. Through a combination of biomaterials and bioactive molecules, this strategy may be further engineered to repair extensive tissue loss in order to relay signals across the lesion epicenter.
An adverse microenvironment as represented by cavitation and accumulation of various inhibitors of axonal regeneration is the main outcome in the lesion area in SCI. In this connection, biological scaffolds have been designed and fabricated to fill the cavities, and improve the local microenvironment by the cells and bioactive molecules attached to them. There are two kinds of biological scaffolds: natural materials and synthetic materials. Natural materials include collagen, hyaluronic acid (HA), agarose, alginate, fibrin, etc., and most of them are nontoxic, biocompatible, and mostly biodegradable 85 . Houweling et al. grafted collagen containing BDNF into the lesion site of the spinal cord, which enhanced the locomotor performance 86 . Paino and Bunge constructed SC-collagen implants and grafted them into cystic cavities formed after SCI; it was compatible with the host, and SCs promoted axonal elongation and remyelination 87 . On the other hand, the poor mechanical strength, variation, and even immunogenicity of natural materials have precluded their wider usage and progress. The plasticity of synthetic materials has overcome this, and furthermore, the synthetic materials have allowed for large quantity production. The nondegradable synthetic materials include silicone, poly(2-hydroxyethyl methacrylate) (PHEMA), poly(N-(2-hydroxypropyl) methacrylamide) (PHPMA), etc. Although it has been reported that these materials could induce regeneration of nerve fibers through the lesion area 88 , the poor biocompatibility and compression of axons for a longer duration usually require a second surgery to remove the implant. Poly(D, L-lactic acid) (PLLA) and poly(D, L-lactic-co-glycolic acid) (PLGA) are two typical representatives for degradable synthetic materials. They are widely used as temporary extracellular matrices in tissue engineering. PLLA scaffolds with BDNF can enhance axonal regeneration in the lesion area 89 . In our previous study, PLGA scaffolds were constructed and seeded with NT-3-SCs and TrkC-NSCs in vitro. Two weeks later, the artificial neural construct had facilitated neuronal differentiation of NSCs that established connections and exhibited synaptic activities 90 . However, when the cell-PLGA scaffold (artificial neural construct) was grafted into the injury/graft site of the SCI rats, limited regeneration capacity for axons was found 91 . It has been reported that the pH value of PLGA degradation solution is in a narrow range of 4.1–4.5 from 2 to 20 weeks in vitro 92 . In comparison with PLGA scaffold transplants, gelatin sponge (GS) scaffold transplants reduce the acidic environment of the injury/graft site of the spinal cord 93 . Therefore, our results suggest that the GS scaffold may be an ideal bioconstruct to serve as a tissue biomaterial for SCI 93 .
Despite great efforts being made by many, the injured nerve fibers fail to regenerate through the lesioned area so that the normal neural pathway can be reestablished after SCI. In view of this, we put forth another strategy, namely, transplantation of stem cell-derived neural network scaffolds with functional synapses constructed from NSCs, NTFs, and biomaterial scaffolds by neural tissue engineering techniques. The objective was to ascertain if any of the ascending and descending axons, including regenerating CST, would establish synaptic contacts with NSC-derived neural network and, if so, whether it would serve as a relay “station” or “center” to conduct signals from the brain to hindlimbs through the injury/graft site of the spinal cord. This is based on the premise that under normal physiological conditions the PSNs and interneurons could also serve as relay neurons for conduction of signals. In addition to descending signals from the brain, movements of limbs are regulated by spinal locomotor circuitries. In higher vertebrates, locomotor circuitries include three major cell types: excitatory interneurons that receive reticulospinal inputs for generating the rhythmic output, which is then distributed to all neurons on the same side of the body; two inhibitory interneurons, one contralateral and one ipsilateral, which control alternation of the left and right motor rhythms; and output motor neurons 94 .
PSNs and interneurons are fundamental for varying degrees of spontaneous recovery in SCI. Studies of PSNs in SCI have found spontaneous repair of the spinal cord, which continues 1 month after injury. This is because the PSNs that bypass the lesion mediate spontaneous functional recovery by reconnecting the injured axons and eventually reorganize the relay connections 95 . Bareyre et al. reported that 12 weeks after transection of CSTs in rats, long PSNs that bridged the lesion area were maintained and arborized on lumbar motor neurons, creating a new intraspinal circuit relaying cortical input to its original spinal targets 77 . We maintain that the above-mentioned results have provided a solid theoretical foundation for transplantation of stem cell-derived neural network scaffolds with functional synapses. Once the stem cell-derived neurons form connections with injured neuronal circuits, the artificial neural network would serve as a relay resembling interneurons to conduct signals.
Abematsu et al. reported that stem cell-derived neurons could integrate into the host and become a new intraspinal relay to reinstate neural communication between the segments above and below the lesioned area of the spinal cord 80 . Regeneration of CST was rather limited, but synapse formation was observed inside the implants. Using a mouse model of SCI, transplantation of NSCs together with administration of valproic acid (VPA), a known antiepileptic and histone deacetylase inhibitor, promoted the differentiation of transplanted NSCs into neurons and migrated into the rostral and caudal areas of the injury/graft site. About 20% of the surviving cells differentiated into MAP2-positive neuron-like cells and extended their processes 80 . As no obvious axon reextension was found, it was assumed that transplant-derived neurons reconstructed the disrupted neuronal circuits in a relay manner. To test this hypothesis, the axon-sparing excitotoxin NMDA was injected at 12 weeks after SCI into the epicenter of the injured site in mouse spinal cords and in the control animals to ablate local neurons in the gray matter. In uninjured mice, NMDA injections had no significant effect on hindlimb function. However, NMDA injections completely reversed both spontaneous and treatment-provoked functional recovery of hindlimb movement in the SCI model mice, indicating that both endogenous and transplant-derived local neurons indeed play an important role in the restoration of hindlimb motor function 80 . Lu et al. grafted NSCs expressing green fluorescent protein (GFP), which were embedded into fibrin matrices containing growth factor cocktails, to the lesion sites of severe SCI 84 . Extending axons of the grafted cell-derived neurons covered remarkable distances and formed abundant synapses with host cells. Axons from the host also extended into the implants. These functional stem cell-derived neurons (relay cells) were significant to locomotor functional recovery, for the recovery disappeared when the injury/graft site was transected again. Although it was discovered in a follow-up replication study that there was a discrepancy in continuous bridging of neural tissue between rostral and caudal segments, the basic finding that transplants of NSCs embedded in fibrin resulted in robust extension of axons that formed plenty of synapses with the host has been confirmed 96 .
Inspired by these studies, we assumed that the mechanism of cell transplantation therapy lies not only in replacing the lost cells but also in providing an optimal environment for endogenous recovery and in remolding of the neural network. With this consideration, we constructed a functional neural network using a three-dimensional (3D) GS scaffold to explore whether it could form synapses and integrate into the host as a functional graft 97 . As distinct from the previous experiments whereby stem cells were transplanted immediately after SCI, we cocultured NT-3-SCs and TrkC-NSCs in the GS scaffolds, which were incubated for 14 days prior to transplantation. Scanning electron microscopy (SEM) showed frequent cell contacts between long processes of different cells including SC-like cells and neuron-like cells. Moreover, NF-positive fibers from GFP-positive NSC-derived neurons were ensheathed by myelin basic protein (MBP)-positive myelin sheaths. Immunoexpression of PSD95 (a marker of postsynaptic component) and synaptophysin (SYP; a presynaptic marker), along with the transmission electron microscopy (TEM), revealed that many synaptic-like contacts were established between long processes of different cells. Furthermore, NSC-derived neuron-like cells were immunopositive for choline acetyl transferase (ChAT), glutamate, or γ-aminobutyric acid (GABA), indicating that these synaptic-like structures have properties for synaptic transmission necessary for neuronal communication. More importantly, spontaneous postsynaptic currents were recorded by whole-cell patch clamp. These results lend strong support to the notion that GS scaffolds can provide a biologically favorable microenvironment for neural restoration and differentiation in order to form a neural network with synaptic function.
In our further studies of the neural networks in vivo 97 , a 2-mm cord segment and associated spinal roots were completely removed at the T10 spinal cord level, and the preconstructed neural networks (NT-3-SCs + TrkC-NSCs) were grafted to fill up the tissue gap. At the end of the eighth week, animals exhibited an average BBB score of 8.09 ± 0.94. Axons from the host regenerated into the implants and formed synapses between transplanted NSC-derived neurons and host neurons in the epicenter of the injury/graft site. Axons both from host or NSC-derived neurons were ensheathed by myelin sheaths. Synapses were observed between two transplanted neurons, or between transplanted neurons and host neurons. Our results strongly suggest that the NSC-derived neural network had integrated into host neural networks and formed a relay to conduct signals from the brain to the hindlimbs through the injury/graft site of the spinal cord 97 . Thus, SCs and interactions between NT-3 and TrkC in a favorable microenvironment, as provided by the GS scaffolds, are pivotal to successful construction of a tissue-engineered neural network.
Because of the potential ethical issues associated with transplanting NSCs, clinical application is currently limited. Bone marrow-derived mesenchymal stem cells (BM-MSCs), however, are readily available, and the cells may then be induced to proliferate in vitro. In autologous transplantation, immunological rejection and ethical problems may be obviated; hence, it has been used for bone, cartilage, tendon, and muscle repair 98 . Because of this, MSC transplantation has attracted the attention of many researchers and has been used in many studies99–101, including ours, in which NT-3-SCs and TrkC-MSCs were embedded in the GS scaffolds to build an artificial synaptic connection 102 .
Transdifferentiation of MSCs into neurons is still controversial, although many in vitro studies103,104 and transplantation studies105,106 showed neuronal properties of MSC-derived cells under different inducing environments. A major concern is that these transdifferentiated neuronal cells failed to incorporate into the host neural network and hence were void in function. Nevertheless, neuronal electrical activities have been recorded from MSC-derived neurons in several electrophysiological studies104,107. Using SCs to induce neural differentiation of MSCs, we compared the differentiation rates in 2D culture 108 and in 3D PLGA multiple-channel conduit culture 109 . Results showed the potential neuron-inducing effect of 3D culture. Considering that the host niches may exert a profound influence on stem cells 110 , we used alternative 3D GS scaffolds with good cytocompatability and histocompatibility 111 to deliver cells. To apply the above-mentioned findings in SCI repair and in a separate study, we took a combinatory strategy by coculturing MSCs, genetically engineered to overexpress TrkC, with SCs that overexpressed NT-3 in 3D GS scaffolds for in vitro induction and a transplantation study. Fourteen days after coculturing with SCs in the GS scaffolds, MSCs differentiated into cells immunopositive for a cholinergic biomarker, which accounted for 84.64 ± 0.94% of the total MSCs grafted 102 . The high incidence of cholinergic neuron-like cells occurred only in the NT-3-SC + TrkC-MSC group. The majority of neuron-like cells in the GS scaffolds expressed PSD95 and SYP. Synaptic structures and some vesicles in the presynaptic component were observed between cells by electron microscopy. The vesicles might represent less mature or some unique synaptic types in neuron-like cells transdifferentiated from MSCs. Spontaneous postsynaptic currents (sPSCs) generated by MSC-derived neuron-like cells were detected by whole-cell patch clamp 102 . The results indicate that MSC-derived neuron-like cells within the scaffold may have synaptic transmission potential as required for neuronal communication.
Following a complete transection of the spinal cord, and the removal of a 2-mm segment of the cord at the T10 level, the lesion gap was filled by MSC-derived synaptic connections as constructed in our study. Remarkably, at 8 weeks after surgery, the tissue cavities were noticeably reduced in the injury epicenter, indicating a tissue-support effect of gelatin implants. A large number of NF-positive nerve fibers of varying lengths were observed to traverse through the injury/graft site, especially the epicenter. These nerve fibers had replaced large cavities and bridged the two ruptured stumps of the spinal cord. Synapses were observed between two adjacent transplanted neuron-like cells, or between transplanted neuron-like cells and host neurons 102 . The synaptic structure was characterized by the presence of a few vesicles in the presynaptic component that might contain acetylcholine. 5-HT-positive fibers were also observed to form connections with the transplanted neuron-like cells in areas rostral to the injury/graft site 102 . Because neither CST fibers nor 5-HT descending fibers could pass through the injury/graft site, it was determined that descending fibers from the brain first cross through the relay, and thereafter through PSNs in T10-L1 to then excite the anterior horn motoneurons at the L2-L4 level to finally regulate the movement of the hindlimbs.
So far, MSCs were either applied as prototypes due to their immunomodulatory and/or prohomeostatic benefit 112 , or predifferentiated into specific cell types to repair tissue loss105,106. Compared with these studies, our current work provides a new perspective on the function of MSC-derived neuron-like cells, i.e., a combined function of terminally differentiated cells and stem cells. The coexistence of neuronal and mesenchymal properties as observed from in vitro and in vivo studies suggests that MSC-derived neuron-like cells may be a new type of cells with therapeutic effects on SCI. The functional multipotency of MSC-derived neuron-like cells may serve as a unique cell type, serving as a relay for the interrupted neural pathways in injured spinal cords.
Concluding Remarks
The key approach in tissue engineering consists of provision of an interactive environment between cells, scaffolds, and bioactive molecules to promote tissue repair. Experimental evidence derived from our recent series of studies supports that a combined treatment comprising cell replacement, gene therapy, and biomaterial scaffolds is an effective strategy for SCI (Table 1). Along these lines, we have shown that SCs could form myelin sheaths and secrete numerous NTFs. It was found that the interaction of NT-3 and TrkC is essential to promote stem cell survival and neural differentiation. Three-dimensional GS scaffolds as fabricated by us had created a favorable microenvironment for embedded cells to connect with each other and form a functional neural network (exogenous neuronal relay) 113 . It is hoped that such a neural network with potential synaptic function may serve as a relay in the injured spinal cord (Fig. 1). Work is now in progress to further explore the combined treatment for more effective treatment of SCI. This involves (1) construction of an artificial spinal cord-like tissue and (2) 3D printing of an artificial spinal cord graft. In the present constructed stem cell-derived neural network, the stem cell-derived neurons indeed can grow, but their processes extend in a haphazard manner so that the regenerating nerve fibers of the host spinal cord cannot integrate with the grafted neural network properly. The artificial spinal cord-like tissue that we aim to fabricate, however, is composed of a white and gray matter-like structure mimicking the natural spinal cord structure. The white matter-like structure is a gelatin ring seeded with genetically modified CNTF-oligodendrocyte precursor cells (OPCs). Inside the ring, the gray matter-like structure is a gelatin cylinder seeded with NT-3-NSCs and TrkC-NSCs. As for 3D printing of an artificial spinal cord graft, decellularized nerve tissue and hydrogel are the main ingredients of the bio-ink. Various growth factors and NSCs for spinal cord construction can be loaded into the printed scaffolds. All of these methods would be future aims to create more histocompatible neural grafts mimicking the spinal cord, so that the transplants can integrate more effectively into the host tissue.

A schematic diagram depicting the integration of neural stem cell (NSC)- or mesenchymal stem cell (MSC)-derived neural network scaffold preconstructed in vitro with the host neural network with synaptic functional neurons. A 2-mm segment of the spinal cord at the T10 level was removed, and the gap was filled by the neural network scaffold. The NSC- or MSC-derived neural network scaffold has been shown to create favorable microenvironments for neural regeneration and survival. On the one hand, the grafted cells (green) form synaptic contacts with regenerating corticospinal tract (CST), 5-HT nerve fibers (5-HT NF), or fasciculus proprius in the area rostral to the injury/transplantation area; on the other hand, they show synaptic contacts with the interneurons or motor neurons at sites caudal to the injury/transplantation area. The artificial neural network with synaptic contacts appears to integrate well with the host tissue and serve as an exogenous neuronal relay for signal transfer. In other words, the motor command from the brain or proprioception from paralyzed limbs may be transmitted through the NSC- or MSC-derived neural network even if the passage of CST or 5-HT nerve fibers through the injury/transplantation area was to be impeded. Hence, with proper rehabilitation, and with increases in signal transfer through the exogenous neuronal relay, it is envisaged that the motor/sensory function may be improved.
Comparison of Effects of Different Graft Treatments After Spinal Cord Injury
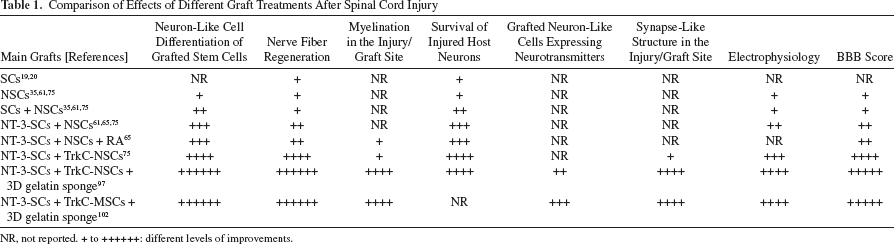
NR, not reported. + to ++++++: different levels of improvements.
Footnotes
Acknowledgments
This research was supported by grants from the Chinese National Natural Science Foundation (Nos. 81330028 and U1301223), National 863 Project (No. 2013AA020106), Foundation of the Education Ministry of China (No. 201300193035), Foundation of Guangdong Province (No. 2011A030300004), and Co-innovation Foundation of Guangzhou (No. 201508020215) to Y.-S. Zeng. The authors declare no conflicts of interest.