
Abstract
Hepatocyte transplantation is a promising alternative therapy for the treatment of hepatic failure, hepatocellular deficiency, and genetic metabolic disorders. Hypothermic preservation of isolated human hepatocytes is potentially a simple and convenient strategy to provide on-demand hepatocytes in sufficient quantity and of the quality required for biotherapy. In this study, first we assessed how cold storage in three clinically safe preservative solutions (UW, HTS-FRS, and IGL-1) affects the viability and in vitro functionality of human hepatocytes. Then we evaluated whether such cold-preserved human hepatocytes could engraft and repopulate damaged livers in a mouse model of liver failure. Human hepatocytes showed comparable viabilities after cold preservation in the three solutions. The ability of fresh and cold-stored hepatocytes to attach to a collagen substratum and to synthesize and secrete albumin, coagulation factor VII, and urea in the medium after 3 days in culture was also equally preserved. Cold-stored hepatocytes were then transplanted in the spleen of immunodeficient mice previously infected with adenoviruses containing a thymidine kinase construct and treated with a single dose of ganciclovir to induce liver injury. Engraftment and liver repopulation were monitored over time by measuring the blood level of human albumin and by assessing the expression of specific human hepatic mRNAs and proteins in the recipient livers by RT-PCR and immunohistochemistry, respectively. Our findings show that cold-stored human hepatocytes in IGL-1 and HTS-FRS preservative solutions can survive, engraft, and proliferate in a damaged mouse liver. These results demonstrate the usefulness of human hepatocyte hypothermic preservation for cell transplantation.
Introduction
Currently, liver transplantation is the gold standard treatment for hepatic failure, hepatocellular deficiency, and genetic metabolic disorders. However, this invasive procedure is expensive, associated with high morbidity, and is limited by the shortage of transplantable organs. Biotherapy, such as the use of a bioartificial liver or hepatocyte transplantation, could be an alternative for transient or permanent liver metabolic support. Hepatocytes isolated from livers that are not suitable for orthotopic transplantation could provide an additional chance for patients on waiting lists and a unique opportunity for those who are not eligible for liver transplant. The development of approaches for hepatocyte preparation in specialized centers with the aim of providing on-demand sufficient amounts of good quality hepatocytes could broaden the scope of these therapies (42). As the hepatocyte source (i.e., donors) is inherently unpredictable, it is necessary to establish the best strategy to identify and prepare the recipient patients and to handle and dispatch the cells to be injected. Hepatocyte cryopreservation and banking is theoretically the most adapted practical solution. Freezing/ thawing protocols have been considerably improved over the years, and now cryopreserved hepatocytes are viable, with some metabolic activities close to those of fresh hepatocytes (e.g., CYP) for at least a short period after thawing (28,44). However, poor attachment to culture substrates is often reported, thus limiting their long-term in vitro use and questions their ability to adhere to the hepatic sinusoidal endothelium after transplantation (24,47,49). Indeed, there is evidence that cryopreserved hepatocytes are less efficient than fresh hepatocytes for transplantation (9,33). Alternatively, freshly isolated hepatocytes could be cold stored in conditions similar to those used for liver preservation before transplantation (17,19,30,36,39. This approach has been widely explored in experimental settings using different organ preservative solutions or defined culture media and isolated (2,34,37,40,41 or cultured hepatocytes (8,26,39), mainly of animal origin. Most of these studies focused on the recovery of the cell metabolic capacities after rewarming, with the aim of providing a convenient cell source for in vitro drug discovery platforms. To our knowledge, the engraftment and repopulation of a damaged liver by cold-stored human hepatocytes has never been tested. We thus studied the impact of hypothermic preservation in three clinically safe (i.e., drug-agency approved) and animal product-free preservative solutions on the viability and in vitro functionality of human hepatocytes and also after transplantation in mice with liver damage (32). Hypothermic preservation slows down cell metabolism and leads to structural membrane damage and reduction of the ATP intracellular pool (16,30,46); therefore, different solutions have been elaborated to counteract these problems. For this study we chose three cold storage solutions designed for use in organ transplantation (13). The University of Wisconsin (UW) solution was developed in the late 1980s to prevent cell swelling, intracellular acidosis, injury from oxygen-free radicals, and to maintain the ATP levels (45). Since then, it has become the standard solution for the preservation of most organs for transplantation and also of many cell types in suspension, including human hepatocytes shipped for transplantation in clinical trials (15,19). It is now considered as the reference for clinical applications. Hypothermosol-FRS (HTS-FRS) is an improved formulation of the intracellular-like solution Hypothermosol (6) that suppresses free radical formation in cells undergoing prolonged hypothermic preservation (29) and preserves human hepatocyte integrity and metabolizing functions (36). We also included Institut Georges Lopez 1 (IGL-1), another organ preservative solution that contains polyethylene glycol (PEG), as colloid, and has a high extracellular sodium/potassium ratio. These features should improve liver preservation (4). IGL-1 was found to be as efficient and safe as UW while less expensive (10); however, it has never been used for cold preservation of isolated human hepatocytes.
Materials and Methods
Chemicals
Culture media Ham-F12 and William's medium E, additives, and collagenase type IV were purchased from Sigma-Aldrich (Saint-Quentin Fallavier, France), fetal calf serum from Lonza (Levallois-Perret, France), and cytokines/ growth factors from Peprotech (Neuilly sur Seine, France). Hepatocytes in suspension were preserved in IGL-1 solution (Institut Georges Lopez, Lissieu, France), University of Wisconsin solution (UW; VIASPAN; Bristol-Myers Squibb, Rueil Malmaison, France), or HTS-FRS (BioLife Solutions Inc., Bothell, WA, USA).
Human Hepatocyte Isolation
Liver samples were obtained either after resection for medical reasons (primary tumor or metastasis) or from donors when the liver was considered unsuitable for organ transplantation. The use of human liver tissues to prepare hepatocytes for research purposes was approved by the French National Ethics Committees and local legal instances. Informed consent was obtained from each patient or family prior to surgery. Information on the donors of the livers used in this study is shown in Table 1.
Characteristics of the Human Liver Donors

Hepatocytes were isolated by using a two-step perfusion protocol and cultured as described previously by our laboratory (7,11,14,38). Briefly, several veins apparent on the cut edge of the lobectomy were used for sequential perfusions with a washing buffer, with a calcium-chelating buffer made according to Pichard et al. (38), and then with a collagenase solution. After gentle disruption of the tissue and filtration, the post-collagenase homogenate was centrifuged at low speed (50 × g for 5 min) to pellet the hepatocytes. Hepatocytes were then resuspended in “short-term culture medium” (38) supplemented with 2% heat-inactivated fetal calf serum, and seeded at 2.6 × 105 viable cells/cm2 on collagen type I-coated wells (Becton Dickinson, Pont De Claix, France). After overnight attachment, the medium and unattached cells were removed, and fresh “long-term medium” (38) was added. All experiments were carried out in duplicate.
Cold Storage
Hepatocytes were resuspended at the indicated concentrations in the preservative solutions in 50-ml polycarbonate tubes (Becton Dickinson) and left undisturbed for 12, 24, 48, or 96 h at 2-6°C. At the different time points, cells in preservative solutions were diluted with four volumes of “short-term culture medium,” washed once again with 50 ml “short-term culture medium,” and classically centrifuged at 200 × g for 5 min before seeding.
Cell Viability
Viability was estimated based on the trypan blue exclusion method. After cold storage, hepatocytes in suspension were washed twice in “short-term culture medium” (38) and incubated at room temperature with 0.4% trypan blue (Invitrogen, Cergy Pontoise, France) for 5 min. For each sample, the percentage of viable cells was calculated in duplicate.
Cell Attachment Measurement
After cold storage, hepatocytes were washed in “short-term culture medium” and seeded at the same concentration as freshly isolated hepatocytes (FIH) (plated at a density calculated based on their viability before cold storage). Their attachment was estimated by measuring the total amount of proteins recovered after overnight plating. Wells were washed twice with PBS (Invitrogen) to remove unattached cells and medium, and then cells were detached and lysed in reporter lysis buffer (Promega, Charbonnieres, France) through three freezing/thawing cycles. Protein concentration was determined using the bicinchoninic acid (BCA) method (Interchim, Montluçon, France) according to the manufacturer's recommendations. Absorbance at 562 nm was measured using a Mithras LB940 microplate reader (Berthold Technologies, Thoiry, France), and the total protein concentration per well was calculated and expressed as the percentage of the protein concentration of freshly isolated hepatocytes.
RNA Analysis by RT-PCR
Total RNA was extracted from human hepatocytes (primary cultures) or mouse tissues using the TRIzol reagent, according to the manufacturer's recommendations (Invitrogen), and 500 ng of total RNA was reverse transcribed using random hexaprimers and the MMLV Reverse Transcriptase Kit (Invitrogen). Primer sequences are in Table 2. When possible, primer pairs were designed from different exons to avoid false-positive results in case of contamination by genomic DNA.
Sequences of Primer Pairs Used for Quantitative and Semiquantitative RT-PCR Analysis

Quantitative PCR was performed using the Roche Molecular Biochemicals Light Cycler system (Roche Diagnostic, Meylan, France) with the following program: one step at 95°C for 10 min, 55 cycles of denaturation at 95°C for 15 s, annealing at 65°C for 15 s, elongation at 72°C for 15 s. The amplification specificity was verified by determining the product melting curve. Relative expression values were calculated with the LightCycler480 software (Roche Diagnostics) and normalized to the expression of ribosomal protein, large, P0 (RPLP0). RNA from human muscle cells (kindly provided by Dr. G. Carnac, INSERM U1046, Montpellier, France) was included as negative control.
Human liver chimerism in transplanted mice was analyzed by semiquantitative PCR using the Taq Platinum set (Invitrogen) and a Techne TC-512 gradient thermo-cycler (IMLAB, Lille, France) with the following program: one step at 94°C for 2 min, cycles of denaturation at 94°C for 30 s, annealing at 60°C for 30 s, and elongation at 72°C for 30 s, and final step at 72°C for 5 min. Primers are listed in Table 2. PCR products were visualized under UV light after migration in 2% agarose gels containing the SyberSafe DNA gel stain (Invitrogen). The percentage of chimerism was evaluated using RNA samples in which increasing concentrations of human liver RNA (from 1% to 100%) were mixed with mouse liver RNA. The species specificity and efficiency of all primers (including the h/m RPLP0 forward primer that exhibits one mismatch toward human RPLP0 mRNA) were previously validated using mouse and human liver cDNA as templates.
Quantification of Human Albumin, Coagulation Factor VII, and Urea Production and Secretion
Human albumin in the supernatants of hepatocytes or in the serum of chimeric mice following blood sampling from the facial vein was measured using a sandwich ELISA assay according to the manufacturer's instructions (Bethyl Laboratory, Montgomery, TX, USA). Human albumin concentration was calculated by nonlinear regression analysis with the GraphPad Prism 5 software (GraphPad Software Inc., La Jolla, CA, USA).
We adapted a previously described ELISA assay (Diagnostica Stago, Asnieres, France) (7) to quantify the amount of coagulation factor VII in the hepatocyte culture supernatant. The culture medium was used as a negative control.
Spontaneous production and secretion of urea by hepatocytes was quantified using the QuantiChrom Urea Assay Kit (Gentaur, Paris, France), a colorimetric test.
All these analyses were performed in duplicate, and the values in hepatocyte supernatants were normalized to the amount of total protein extracted from the same well and quantified with the BCA method (Interchim, Montluçon, France).
Liver Damage Induction and Human Hepatocyte Transplantation
We used, with some minor modifications, a recently described mouse model in which, following drug-induced liver damage, the liver can be efficiently repopulated by xenogeneic hepatocytes (3,32). Male Balb/C Rag2tm1Fwa Cd3gtm1Amk mice (4-6 weeks old, bred at CIMA, Pamplona, Spain) were intravenously inoculated with 5 × 109 PFU of recombinant adenoviruses expressing herpes simplex virus thymidine kinase (AdTk). In these mice, liver damage is characterized by inflammation accompanied by increased hepatocyte size and enlarged nuclei and provides a growth advantage for transplanted human hepatocytes. The AdTk-GCV effect is maintained for 10 weeks and then is progressively resolved, as indicated by regenerative nodules (32). As soon as the human liver perfusion was planned, a single dose of 50 mg/kg ganciclovir (GCV; Roche) was intraperitoneally administered to induce subacute liver damage. Human hepatocytes were isolated at the production site (Montpellier, France) and were cold preserved in HTS-FRS or IGL-1 at 1 × 106 cells/ml on ice during shipment to the transplantation site (Pamplona, Spain). Thirteen to 15 h after isolation, 1.5 × 106 viable human hepatocytes suspended in 100 μl PBS were inoculated in the spleen of mice. Freshly isolated hepatocytes were not available. As cultured human hepatocytes were previously shown to be a suitable source of cells for the generation of chimeric livers in this model (32), hepatocytes from the same donor were cultured in long-term medium for 7 days and were transplanted in a group of mice used as controls.
Then animals received intravenously 100 μl of human AB serum (Sigma-Aldrich) and were treated with antibiotics at 25 mg/ml in drinking water (enrofloxacin; Bayer HealthCare, Barcelona, Spain) for 1 week. Blood was collected by retro-orbital bleeding once per week, and cell engraftment was followed by quantifying serum human albumin (ELISA assay; Bethyl Laboratories Inc.), starting at 3 weeks posttransplantation when the injected human serum was cleared. The serum levels of alanine and aspartate transaminase (ALT, AST) in transplanted mice were quantified using a Cobas C311 analyzer (Roche). The study protocol was approved by the Animal Experimentation Ethics Committee of the University of Navarra, Spain (permit Nos. 041-07 and 095-09).
Immunohistochemistry
Immunohistochemical staining was performed using the EnVisionTM+ System (Dako, Glostrup, Denmark) according to the manufacturer's recommendations. Paraffin liver sections (3 μm thick) were sliced, dewaxed, and hydrated, and some of them were stained with hematoxylin (Panreac, Catelar del Valles, Spain) and eosin (H&E) (Merck, Darmstadt, Germany) for routine histology. If necessary, heat-induced antigen retrieval (AR) was performed by incubating sections at 95°C in 0.01 M Tris-1 mM EDTA buffer (pH=9) (Sigma-Aldrich) in a Pascal pressure chamber (S2800; Dako) for 30 min. Proteolytic-induced AR was performed by incubating sections in 20 μg/ml proteinase K (Sigma-Aldrich) at 37°C for 30 min. Endogenous peroxidase was blocked with 3% H2O2 (Panreac) in deionized water for 10 min. Sections were washed in TBS-0.05% Tween 20 (TBS-T) (Fluka, Madrid, Spain) and incubated with primary antibodies [anti-hCK-18, 1:100; M7010 (Dako); anti-albumin, 1:8,000 (Bethyl Laboratories)] at 4°C overnight. After rinsing in TBS-T, sections were incubated with the relevant secondary antibodies (Dako). Peroxidase activity was revealed using DAB+ (K3468; Dako), and sections were lightly counterstained with Harris hematoxylin (Panreac). Finally, they were dehydrated in a graded series of ethanol, cleared in xylene, and mounted in DPX (Sigma-Aldrich). Slides were scanned (Montpellier RIO Imaging facilities, INM Montpellier, France) using a Nanozoomer Slide Scanner (Hamamatsu Photonics, Massy, France), and virtual slides were viewed using the NDP.view software (Hamamatsu Photonics).
Statistical Analysis
Experiments on each liver were carried out in duplicate. Results were expressed as the mean ± SEM. Oneway repeated measures analysis of variance (ANOVA) followed by the Dunnett's post hoc test multiple group comparison was used to analyze group differences. Data were analyzed by GraphPad Prism software version 5 (GraphPad Software Inc.). Values of p < 0.05 were considered significant (p < 0.05, p < 0.01, p < 0.001).
Results
Impact of Hypothermic Preservation on Viability, Morphology, and Attachment of Human Hepatocytes
To determine the optimal cold-storage conditions for human hepatocytes, first we assessed the effect of the concentration at which cells were cold stored on their viability. To this aim, human hepatocytes were stained with trypan blue after 16 h of cold storage in three different preservative solutions (UW, IGL-1, and HTS-FRS). Overall, cold storage reduced cell viability compared to freshly isolated hepatocytes (FIHs) (Fig. 1A), and the deleterious effect was strongest (viability around 40%) at the lowest cell concentration (0.1 × 106 cell/ml). At intermediate cell concentrations (1-10 × 106/ml), viability ranged from 48% to 62%, depending on the storage solutions. IGL-1 solution allowed a slight but significantly better recovery of viable human hepatocytes than UW. The trypan blue exclusion method is based on cell membrane integrity and does not portend cell functionalities. Conversely, cell attachment to a substratum is an energy-demanding process indicative of cell vitality. Cell attachment was thus assessed in parallel to viability measurements, assuming that hepatocytes that attach and spread on the substrate are viable and functional. Hepatocytes that had been cold preserved in suspension at different concentrations (see Fig. 1A) for 16 h were thus seeded on collagen type I-coated plates at a density calculated based on their viability before cold storage (i.e., FIH viability). Their ability to form a monolayer was assessed by phase contrast microscopy after overnight culture. Hepatocytes exhibited the same characteristic polygonal morphology before and after cold storage in the three solutions (Fig. 1B), and cell confluence varied depending on the storage conditions (cell concentration and solution), as observed for cell viability (Fig. 1A). Based on these results and on literature data (36), for the subsequent experiments, hepatocytes were cold stored in suspension at a concentration of 1 × 106 cells/ml. We then examined the impact of the cold storage duration on cell viability and attachment (Fig. 1C). For all hepatocyte samples (isolated from three different livers with viabilities of 78%, 95%, and 84%), cell viability declined progressively with the storage time (from 72% after 12 h down to 35% after 96 h of storage) in all solutions. No difference was observed concerning the three cold storage solutions, except after 24 h of hypothermic preservation when cells stored in IGL-1 or HTS-FRS showed a significantly better viability than cells preserved in UW. Finally, human hepatocytes were seeded on collagen type I-coated plates before and after cold storage, and plating efficiency was evaluated 16 h later by quantifying the total protein amount per well (Fig. 1D). As with cell viability, the cell attachment declined progressively over storage time, from 100% after 12 h down to 13% after 96 h of cold storage, but without any significant difference between the three preservative solutions. The difference observed between changes in viability and attachment can be explained by the fact that cells with little membrane damage may be stained blue, but are still viable and have the potential to repair and recover. Overall, this experiment indicates that viable hepatocytes after cold storage in the three preservative solutions retain their adhesive properties.
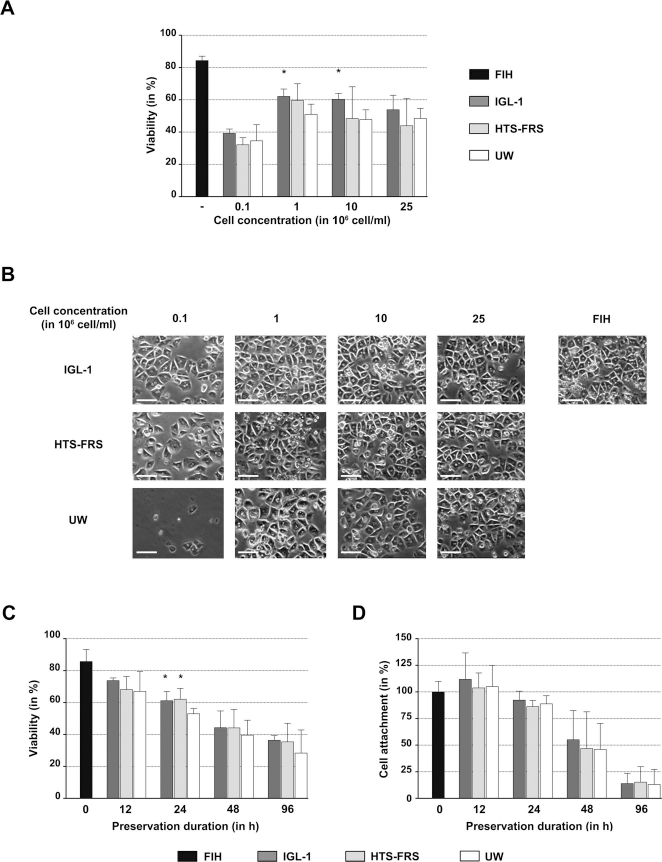
Impact of cold preservation on human hepatocyte viability and attachment. Different concentrations (0.1, 1, 10, and 25 × 106 cells/ml) of human hepatocytes were preserved in suspension at 2-8°C using different organ preservation solutions (IGL-1, HTS-FRS, and UW) for 16 h. (A) Cell viability was then assessed by using the trypan blue exclusion method (freshly isolated hepatocytes, FIH). (B) Hepatocytes were then seeded on collagen type I-coated plates according to their viability before hypothermic preservation, and their morphology and confluence were visualized by phase contrast microscopy after 16 h in culture. (C, D) Hepatocytes were preserved in suspension (1 × 106 cells/ml) at 2-8°C in the different organ preservation solutions for up to 4 days. At different time points (12, 24, 48, 96 h) cells were washed, and (C) their viability was assessed by using the trypan blue exclusion method, and (D) their attachment capacity was evaluated by seeding on collagen type I-coated plates according to their viability before hypothermic preservation and by quantifying the total protein amount per well after 16 h in culture. Results are expressed as the percentage of the value measured in cells plated immediately after isolation (time 0). Statistical analysis was performed using an ANOVA followed by the Dunnett's post hoc test multiple group comparison, with UW value as a reference (*p < 0.05). Scale bar: 100 μm.
Phenotype and Functionality of Human Hepatocytes After Cold Preservation
The deleterious effect of hypothermia/hypoxia, such as apoptosis and mitochondrial dysfunction, can be revealed and/or amplified during the rewarming period (12,39,43). A viability of 60% is the consensual acceptable limit for the use of isolated hepatocytes in clinical settings; thus, we were mostly interested in analyzing the phenotype of hepatocytes selected for this criteria. Therefore, the functionality of human hepatocytes after 12 and 24 h of hypothermic preservation in the different solutions was assessed after return to normoxic temperature and 3-day culture and compared to that of hepatocytes seeded directly after isolation.
In these experiments, fresh (FIHs) and cold-preserved hepatocytes were seeded based on their viability to establish a confluent cell monolayer in order to avoid the cell dedifferentiation observed in subconfluent human hepatocyte primary cultures (21,27). Q-PCR analysis (Fig. 2A) revealed that the mRNA expression of HNF1α, HNF4α, C/EBP4aL and C/EBPβ (liver-enriched transcription factors that are critical for the highly differentiated hepatocyte phenotype and function) was well maintained after 12 h of cold storage. After 24 h, a slight, but not significant, decrease (due to the interindividual variability among liver donors) compared to FIHs was observed. Similarly, the ability of FIHs and cold-stored hepatocytes to synthesize and secrete coagulation factor VII and urea (the clinical biological parameters usually followed in patients with liver disease) in the extracellular medium after 3 days of culture was preserved, but albumin secretion was significantly decreased after 24 h of cold storage (Fig. 2B). Overall, these results demonstrate that cold preservation up to 12 h does not dramatically affect the phenotype and synthetic functions of human hepatocytes in primary culture, regardless of the preservative solution used.

Phenotype and functionality of human hepatocytes cultured after hypothermic preservation. Hepatocytes were seeded on collagen type I-coated dishes directly after isolation (FIH) or after cold preservation in suspension in the different solutions for 12 or 24 h and cultured for 3 days. (A) The mRNA levels of hepatic genes encoding liver-enriched transcription factors (HNF1α, HNF4α, C/EBPα, and C/EBPβ) were quantified by qRT-PCR and expressed as the percentage of their expression in FIHs. (B) Hepatocyte synthetic functions were assessed by quantifying the secretion of albumin, coagulation factor VII, and urea in the medium. Statistical analysis was performed using an ANOVA followed by the Dunnett's post hoc test multiple group comparison, with FIH value as a reference (*p < 0.05).
Transplantation of Cold-Preserved Human Hepatocytes
We then investigated whether cold-preserved hepatocytes in IGL-1 and HTS-FRS solutions could be used for cell transplantation. For this purpose, we used a simplified mouse model of liver injury (32) that relies on the rapid induction of liver damage in a temporally controlled manner following a single intraperitoneal injection of GCV at high dose in immunodeficient mice infected with AdTk. The experimental protocol is described in Figure 3A.
In this pilot study, two independent transplantation experiments (Experiment I: mice with mild liver damage—ALT mean value between 300 and 450 IU/ml; Experiment II: mice with sustained liver damage—ALT mean value between 500 and 800 IU/ml) were carried out. For each experiment, hepatocytes from a single liver were used after cold storage in IGL-1 or HTS-FRS (one group of mice for each solution; three to seven mice/group) or after culture (only for Experiment II). Hepatocyte viability after cold storage at the time of the intrasplenic injection (TO) ranged between 62% and 70%, corresponding to a recovery between 72% and 97%, when referred to the FIH viability (Table 3). Four mice died from surgical complications at day 2 postgraft in Experiment I (Table 3). The remaining mice survived until sacrifice at week 9 after transplantation, whereas some of the animals included in Experiment II died around week 5 after transplantation due to illness related to the liver injury (Table 3). Transplantation outcomes of Experiments I and II are presented in Figures 3 and 4. Engraftment was monitored over time by measuring human albumin in the recipients’ blood. Its concentration was very variable, even within the same group and in animals that received hepatocytes from the same liver. Overall, human albumin serum concentration gradually increased between weeks 3 and 9 posttransplantation. However, it was much lower in animals included in Experiment I (mild liver injury) (Fig. 3B) than in those in Experiment II (high liver injury) (Fig. 3C), as expected. The presence of human hepatocytes within the host liver was confirmed by assessing the mRNA expression of several human hepatic markers (Fig. 4A, B). Based on the albumin production, hepatocytes that had been cold stored in IGL-1 and HTS-FRS showed equivalent engraftment capacity in both Experiments I and II. Indeed, the levels of circulating human albumin increased in a comparable manner in both HTS-FR and IGL-1 groups, reaching around 20 μg/ml and 720 μg/ml in Experiments I and II, respectively, at the end of the experiment (week 9). In contrast, human albumin production by human hepatocytes transplanted after 7 days in culture (D7) was significantly lower and remained stable over time (Fig. 3C). Repopulation of mouse livers by human hepatocytes was evaluated also by immunohistochemistry on a small number of animals (Fig. 4C). Five weeks after cell graft, human hepatocytes could be identified with antibodies against human albumin and CK18 (data not shown) in the liver lobes. They were organized in small clusters, reflecting their selective expansion after successful engraftment. Four weeks later (9 weeks after transplantation), the clusters of human hepatocytes were larger, suggesting several rounds of cell division in a subset of transplanted cells. At this stage, in H&E-stained sections, clusters of human hepatocytes could be clearly differentiated from mouse hepatocytes by their size and the paler cytoplasm, as previously described (31,32,48). Native regenerative nodules were also visible (Fig. 4C). Human hepatocytes were not found in the lungs, and only some residual hepatocytes were observed in the spleen (Fig. 4D), especially after injection of D7 hepatocytes, as revealed by the detection of human hepatic markers in this organ (Fig. 4B). In conclusion, cold-preserved human hepatocytes retain their ability to engraft and repair a damaged liver, whatever the preservative solution, and display normal biological functions, as shown by the albumin production over a 9-week period after transplantation. Cultured human hepatocytes can also engraft and proliferate in damaged livers, as previously shown (32), but not as efficiently as cold-preserved hepatocytes.

Engraftment of cold-preserved human hepatocytes in mice infected with AdTk after induction of mild liver damage by injection of ganciclovir. (A) Schematic of the transplantation experiments: immunodeficient mice infected with adenoviruses expressing thymidine kinase (AdTk) received intraperitoneally a single high dose of ganciclovir as soon as the human liver perfusion was planned. Isolated human hepatocytes were cultured or preserved in suspension (1 × 106 cells/ml) on ice in the different solutions (IGL-1 and HTS-FRS) and shipped to the transplantation site in Pamplona, Spain. They were injected in the spleen of mice with mild (Experiment I) or sustained (Experiment II) liver damage 13 to 15 h after liver dissociation and cold preservation or after a 7-day culture period (D7). Human hepatocyte engraftment was evaluated by measuring the production/secretion of human albumin in the serum of recipient mice at different time points after transplantation (3 to 9 weeks) in Experiment I (B) and Experiment II (C). Values are the mean albumin concentration per group.

Analysis of chimeric livers in AdTk/GCG-treated mice after transplantation of cold-preserved or cultured human hepatocytes. Human hepatocyte engraftment was evaluated at 9 weeks posttransplantation at the end of Experiment I (A) and Experiment II (B) by semiquantitative RT-PCR analysis of the expression of human hepatic genes in the liver, spleen, lung, and muscle. Human–mouse RNA solutions containing decreasing percentages (from 100% to 1%) of human mRNA were included to estimate the human/mouse liver chimerism. (C) Immunohistochemistry was performed to specifically detect human CK18 (hCK18) and albumin (hAlb) in chimeric livers. The presence of growing human hepatocyte nodules was evidenced at week 9 posttransplantation. Native regenerative nodules were observed after hematoxylin and eosin (H&E) staining. Scale bar: 100 μm. (D) Dissemination of the infused human hepatocytes in the recipients’ body was investigated after sacrifice at the end of the experiment (9 weeks posttransplantation) by immunohistochemistry with antibodies against human CK18 in lung, muscle, and spleen tissue sections. Scale bar: 1 mm.
Transplantation of Cold-Preserved Human Hepatocytes to Rescue Mice With Liver Failure

Hepatocyte viability and mice survival outcomes in two independent transplantation experiments (Experiments I and II). For each experiment, hepatocytes from one single liver were used. Ratios refer to the number of surviving mice/total number of mice at the time point indicated in parentheses.
Discussion
The objectives of this study were to evaluate hypothermic conditions for preserving FIHs in clinically safe preservative solutions and then to determine whether and to what extent cold-stored hepatocytes can repopulate a damaged liver in a mouse model.
Plating efficiency is a valuable criterion of cell vitality and is assumed to correlate with their engraftment capacity in the liver. As described by others, we found that viability and attachment of human hepatocytes after hypothermic storage in suspension were dependent on cell density and on the preservation period. After short hypothermic storage (12 h), hepatocytes in primary culture were as healthy and functional as freshly isolated cells. Previous studies reported comparable gene expression profiles (44) and xenobiotic metabolism capacity and clearance predictions in human hepatocytes in suspension after cold storage and in freshly isolated cells (19,36). We found that human hepatocytes could be cold stored at 4°C for up to 24 h without significant decrease in functional integrity, as assessed by their ability to attach on a substratum and to produce and secrete urea and factor VII, and with a viability higher than 60%, which is the threshold limit value currently accepted as an insurance of hepatocyte quality for biotherapy programs (1). Moreover, we did not observe any clear specific effect of the three storage solutions, although IGL-1 and HTS-FRS seemed to give better results than UW concerning cell survival. These results are in agreement with recent data showing the superiority of HTS-FRS compared to UW for the maintenance of the metabolic activity of human hepatocytes in vitro after 48 h and 72 h of cold storage (36). However, in our study the difference between UW and HTS-FRS was not as significant as in this previous work. Our results are similar to those reported by Gramignoli et al. showing that critical hepatic functions, analyzed in 47 human hepatocyte preparations, were well maintained in hepatocytes after cold storage in UW solution for up to 48 h, although in our hands UW is not as performing as stated in this work (19). The metabolic activity of animal hepatocytes also was reported to be maintained after cold storage in UW solution for 48 h (35). These discrepancies could be related to the age, underlying pathology and medication taken by the liver donors, the impact of the ischemic time on the liver samples and/or species differences (22), and also to the cell dissociation and isolation procedures (collagenases and proteases ratio and activities, composition of the perfusion buffers) that can impact on cell conservation methods (20,34, and unpublished data). Concerning the IGL-1 solution, no other data are currently available regarding its ability to preserve the viability and functionality of human or animal hepatocytes.
Although we observed a decrease in cell viability after 24 h of cold storage, it has to be noted that important synthetic activities required for clinical applications, such as urea synthesis and factor VII secretion, were still well maintained in the surviving cells after 3 days in culture. Urea synthesis, a complex process involving several enzymes, is a good indicator of the degree of mitochondrial preservation. Like others, we found that urea metabolism is better maintained than albumin secretion, which seems more sensitive to long-term cold preservation (19). This could be related to the irreversible cytoskeleton disorganization that is associated with long-term cooling and rewarming and that affects the cell transport dynamics (25,46).
We then evaluated to which extent cold preservation in IGL-1 and HTS-FRS solutions influences human hepatocyte survival and function after transplantation into a suitable animal model. Comparison with freshly isolated cells was not included because our intent was to simulate the clinical situation that occurs when hepatocytes are prepared in specialized centers far away from the transplantation site. For the same reason, the chosen mouse model of AdTk/GCV-induced liver damage (32) was very convenient for performing on-demand transplantation studies, as the availability of human livers is unpredictable. Using this experimental model, we show that human hepatocytes can survive, engraft, and proliferate in a chronically damaged liver after cold preservation in IGL-1 and HTS-FRS solutions. Although it is difficult to measure the exact extent of the contribution of human hepatocytes in the chimeric mouse liver, we think that about 10-15% of hepatocytes were of human origin based on the expression of human-specific liver function markers (by immunohistochemistry and RT-PCR analysis) in the liver of the recipient mice. This estimation is in accordance with the data by Hasegawa et al. (23) showing a high correlation between liver chimerism and plasma concentration of human albumin in recipient mice. Although preliminary (due to the small number of animals included in the study), our data suggest that IGL-1 and HTS-FRS represent alternative conservative solutions useful for biotherapy programs based on hepatocyte transplantation.
IGL-1 previously showed some level of protection against the immune response following kidney and pancreatic islet grafts, possibly due to the “immunocamouflage” provided by PEG interaction with membranes (50). Moreover, according to Ben Mosbah et al., IGL-1 offers a better protection to steatotic livers, which are the main available source of human hepatocytes for liver biotherapy, against the deleterious effects of cold ischemia–reperfusion injury than the UW solution (5). Whether these protective effects are also effective on hepatocytes isolated from such livers is an interesting question that deserves further studies.
Compared to cryopreservation, hypothermia is a milder technique that does not require the use of potentially deleterious chemicals, liquid nitrogen, or special equipment for cell storage and might allow shipping to remote sites for transplantation. Its main disadvantage is the relatively short period of time that cells can be preserved. However, recent advances in preservation methods based either on supercoiling (51) or on the formulation of new solutions better adapted to cold storage has already allowed increasing the preservation time of rat hepatocytes up to 1 week with improved cell survival and function (8,40). The application of these strategies to the preservation of primary human hepatocytes has already given some promising results (39). Hepatocytes may also be preserved by culturing. We show here that our long-term culture conditions favor human hepatocyte survival in monolayer (7,14,18,38,52 and also maintain their ability to engraft and proliferate in a damaged liver microenvironment. However, engraftment and proliferation were less efficient than after cold storage in suspension. Moreover, the culture process introduces additional steps that may increase cell loss after detachment, economic cost, and safety concerns.
In conclusion, we show that primary human hepatocytes can be hypothermically preserved up to 24 h while maintaining a metabolic activity similar to that of freshly isolated cells. This cheap procedure, based on clinically safe and commonly used preservative solutions, allows the easy handling of large quantities of hepatocytes isolated from precious donor livers, optimizing their use in clinical biotherapy programs. Indeed, cold preservation gives the time to deliver cells to distant transplantation centers and facilitates their repeated use when several cell infusions or refilling of an extracorporal bioartificial liver bioreactor are required. Overall, our results demonstrate that human hepatocytes could be successfully cold preserved in clinically safe and commonly used preservative solutions for clinical applications.
Footnotes
Acknowledgements
This work has been funded by the Région Languedoc-Roussillon/Communauté de Travail des Pyrénées. The authors thank Ms. Elisabetta Andermarcher and Ms. Natalie Funakoshi for the professional editing. The authors declare no conflicts of interest.