
Abstract
In vitro liver conservation is an issue of ongoing critical importance in graft transplantation. In this study, we investigated the possibility of augmenting the standard pre-transplant liver conservation protocol (University of Wisconsin (UW) cold solution) with the phenothiazines chlorpromazine and promethazine. Livers from male Sprague-Dawley rats were preserved either in UW solution alone, or in UW solution plus either 2.4, 3.6, or 4.8 mg chlorpromazine and promethazine (C+P, 1:1). The extent of liver injury following preservation was determined by alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities, the ratio of AST/ALT, morphological changes as assessed by hematoxylin-eosin staining, apoptotic cell death as determined by ELISA, and by expression of the apoptotic regulatory proteins BAX and Bcl-2. Levels of glucose (GLU) and lactate dehydrogenase (LDH) in the preservation liquid were determined at 3, 12, and 24 h after incubation to assess glucose metabolism. Oxidative stress was assessed by levels of superoxide dismutase (SOD), reactive oxygen species (ROS), and malondialdehyde (MDA), and inflammatory cytokine expression was evaluated with Western blotting. C+P augmentation induced significant reductions in ALT and AST activities; the AST/ALT ratio; as well as in cellular swelling, vacuolar degeneration, apoptosis, and BAX expression. These changes were associated with lowered levels of GLU and LDH; decreased expression of SOD, MDA, ROS, TNF-α, and IL-1β; and increased expression of Bcl-2. We conclude that C+P augments hypothermic preservation of liver tissue by protecting hepatocytes from ischemia-induced oxidative stress and metabolic dysfunction. This result provides a basis for improvement of the current preservation strategy, and thus for the development of a more effective graft conservation method.
Introduction
Because liver transplantation is the principal treatment for end-stage liver diseases, the temporal limitation imposed on the number of viable donor organs by the current method of liver graft preservation constitutes a major problem for patients with this disease. Liver preservation consists of the methods employed to minimize in vitro damage to tissue in advance of engraftation. For decades, this was achieved through the maintenance of low environmental temperatures (0–4°C) that function to decrease the metabolism and decelerate the degeneration of liver function 1 . More recently, organ preservation solutions such as the University of Wisconsin (UW) cold storage solution have emerged to augment static cold storage (SCS) 2 ; UW accomplishes this by reducing oxidative injury and metabolic disorder 3 . Despite widespread adoption of UW augmentation, however, cell damage and loss continue to limit the duration of graft preservation possible with the current methodology 4 .
The neuroleptic drugs chlorpromazine and promethazine (C+P), both derivatives of the drug phenothiazine, have been used for their anti-psychotic and sedative effects since the 1950s 5,6 . Often administered in combination 6 –9 , C+P appear to exert their effects on the nervous system through the inhibition of carbohydrate oxidation 8,10 ; several studies have reported beneficial antioxidant effects achieved with these agents 7,9,11,12 . Additionally, recent unpublished data from our group have demonstrated that phenothiazines applied after stroke suppress brain metabolism and decrease peroxide production.
The impaired metabolic state with interrupted mitochondrial activity to which liver grafts are subjected in advance of transplantation causes disrupted energy production, resulting in ischemic injury 13 . Under these ischemic conditions, oxygen deprivation impairs oxidative metabolism, significantly limiting ATP production and leading to an overproduction of reactive oxygen species (ROS) 14 that enhances cellular injury. Because of the depressive effect of phenothiazines on oxidative metabolism and injury, we hypothesized that addition of C+P to the UW solution would result in reductions in ROS production and liver cell damage/loss, leading to improved liver preservation. Therefore, we evaluated the effect of phenothiazine-supplemented conservation solution on the rat liver using multiple indices of liver function, measurement of morphological changes, and assessment of the associated oxidative and inflammatory mechanisms in which we expected the phenothiazines to intervene.
Materials and Methods
Subjects
A total of 48 adult male Sprague-Dawley rats (240–260 g, Vital River Laboratory Animal Technology Co., Ltd., Beijing, China) were used in this study. The experimental protocol was approved by the Animal Care and Use Committee of Capital Medical University (Beijing, China), in accordance with the National Institutes of Health (NIH, Bethesda, MD, USA) Guide for the Care and Use of Laboratory Animals. After the rats were adequately anesthetized, a laparotomy was performed through a midline incision. The distal end of the portal pedicle was ligated when a cannula was placed in the lumen of the portal vein. Rats were then perfused with UW solution (Viaspan; DuPont Merck Pharmaceutical Company, Wilmington, DE, USA, preserved at 4°C) through the cannula into the portal vein using a pump set to operate at 30 ml/min, such that a clear fluid began draining from liver. After perfusion, a standard hepatectomy was performed and livers were dissected into patches weighing 2.0 g. Isolated liver specimens were washed in normal saline (NS). In group 1 (n = 6), specimens were then preserved for 24 h at 4°C in 10 ml UW solution. In groups 2, 3, and 4, (n = 6 each), additional C+P (1:1) was added at 240 μg/ml (2.4 mg C+P) in 0.3 ml of NS, 360 μg/ml (3.6 mg C+P), and 480 μg/ml (4.8 mg C+P), respectively; these doses were determined by us in a preliminary study to produce strong cytoprotection.
Measurement of Liver Function
To evaluate hepatocyte integrity after incubation, 2 ml of preservation liquid was collected from all groups and measured with the COBAS INTEGRA 800 analyzer (COBAS, Mannheim, Germany) using the enzyme multiplied immunoassay technique (EMIT) for detection of aspartate aminotransferase (AST), alanine aminotransferase (ALT), glucose (GLU), and lactate dehydrogenase (LDH). To further evaluate damage to hepatocytes, the AST/ALT ratio was calculated 15 .
Histological Evaluation of Cell Damage
Tissue sections were cut into 4 μm-thick slices and stained with hematoxylin-eosin (H&E). The extent of hepatocyte injury was then determined under light microscopy (Carl Zeiss, Jena, Germany) by evidence of architectural alteration and vacuolization. This evidence was collected by a pathologist in a blinded manner. Four fields per section, totaling 5000 μm2, were analyzed under 100x magnification using a computerized image system (Axiovision release 4.7/05-2008, Carl Zeiss) coupled to an optical microscope (Axioskop40) 16 . The ratio of dead/injured hepatocytes to the total number of hepatocytes was calculated; cell injury/death was indicated by the presence of swelling and vacuolar degeneration. To quantify the severity of hepatic injury, a point-counting method on an ordinal scale was used as follows: a grade of 0 was assigned when there was either minimal or no evidence of injury; a grade of 1 was assigned to cells demonstrating mild injury, consisting of cytoplasmic vacuolization and nuclear pyknosis; a of grade 2 was assigned to cells demonstrating moderate-to-severe injury with extensive pyknosis, cytoplasmic hypereosinophilia, and loss of intercellular borders; a grade of 3 was assigned to cells demonstrating severe necrosis with disintegration of hepatic cords, hemorrhage, and neutrophilic infiltration; and a grade of 4 was assigned to cells demonstrating very severe necrosis with disintegration of hepatic cords, hemorrhaging, and neutrophilic infiltration 17 . In our study, a grade of anything greater than 1 on this scale was defined as hepatocyte damage.
Apoptotic Cell Death Detection by ELISA
In both the control and C+P groups, enzyme immunoassay was used to quantify apoptosis-related DNA fragmentation by detecting cytoplasmic histone-associated DNA fragments (Cell Death Detection ELISA; Roche Diagnostics, Indianapolis, IN, USA), as described previously by us 18 .
SOD and MDA Detection
To quantify the extent of oxidative injury, malondialdehyde (MDA) and superoxide dismutase (SOD) concentrations were measured in homogenized liver using fluorescence detection. In accordance with the manufacturer’s instructions (Jiancheng Biotech Co., Nanjing, China), MDA content and SOD activity were monitored with a microplate reader (Thermo Scientific Multiskan MK3, Kanez Laboratory Equipment Co., Shanghai, China) at 532 and 450 nm by MDA and SOD ELISA kits, respectively 19 .
ROS Production
To assess the effect of phenothiazines on oxidative injury of graft hepatocytes, levels of ROS production were determined. The method by which ROS were measured was used by our group previously 20 , and consists in the use of hydrogen peroxidase (H2O2) linked to a fluorescent compound to detect H2O2. Because it dismutates enzymatically (or rapidly in the absence of enzyme catalysis) into H2O2, superoxide is reliably quantified by this method. First, liver tissue was homogenized in buffer (mannitol 225 mM, sucrose 75 mM, EGTA 1 mM, HEPES 5 mM pH 7.4, 1 mg/ml fatty acid-free BSA, 0.1% Triton-X-100). Then, employing the BCA method for determining protein concentration, samples were diluted to 10 mg/ml, and 100 μg/ml digitonin was added. After 30 min incubation, H2O2 in liver homogenates was detected using a commercially available kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). H2O2 combined with molybdic acid to form a complex, which was measured at 405 nm, and the content of H2O2 was then calculated using a microplate reader (Thermo Scientific Multiskan MK3).
Protein Expression
Western blot analysis was used to detect protein expression in the ischemic tissue, as described by us in previous work 21,22 . Upon completion of electrophoresis, proteins were transferred to polyvinylidene fluoride membranes, which were incubated with primary antibodies (rabbit polyclonal anti-Bcl-2, rabbit polyclonal anti-BAX antibody, rabbit polyclonal anti-TN-Fα antibody, or rabbit polyclonal anti- IL-1β antibody; Abcam, Cambridge, MA, USA) for 24 h at 4°C. Next, membranes were washed with PBS three times for 6 min per wash, and then re-incubated with a secondary antibody (goat anti-rabbit IgG, Santa Cruz, Dallas, TX, USA; goat anti-mouse IgG, Abcam) for 1 h at room temperature. Finally, an enhanced chemiluminescence system was used to detect immunoreactive bands. Western blot images for each antibody, including β-actin, were analyzed using an image analysis program (Image J 1.42, NIH, Bethesda, MD, USA) to represent protein expression in terms of relative image density; mean protein expression from the control group was assigned a value of 1 to serve as a reference.
Statistical Analysis
Statistical analysis was performed with SPSS for Windows, version 17.0 (SPSS, Inc., Chicago, IL, USA). The differences among groups were assessed using one-way ANOVA with the significance level set at P < 0.05. Post-hoc comparison among groups was performed using the least significant difference (LSD) test.
Results
Levels of ALT, AST, and the AST/ALT Ratio
Preservation solutions were collected from all groups at 3, 12, and 24 h after incubation. Compared with controls, 2.4 mg phenothiazine did not yield any effect on ALT levels (165.8 vs 165.3 unit/L at 3 h, 417 vs 315 unit/L at 12 h, and 419 vs 356.5 unit/L at 24 h). In the 3.6 mg group, a significant, but relatively small, decrease was found at 3 (108.5 vs 165.3 unit/L, # P < 0.05) and 24 h (310.5 vs 356.5 unit/L, # P < 0.05). In the 4.8 mg group, however, ALT levels in the preservation solution were significantly and markedly decreased at all three time points: 3 (54.3 vs 165.3 unit/L, ## P < 0.01), 12 (141.8 vs 315 unit/L, ## P < 0.01), and 24 h (154.7 vs 356.5 unit/L, ## P < 0.01) after incubation, as compared with the control group (Fig 1A). Similar reductions in AST levels were observed in C+P-treated groups (Fig 1B). Furthermore, the AST/ALT ratio yielded by C+P treatment was significantly decreased in the 3.6 (1.1 vs 1.6 at 12 h, # P < 0.05; 1.2 vs 1.7 at 24 h, # P < 0.05) and 4.8 mg groups (1.1 vs 1.6 at 12 h, # P < 0.05; 1.1 vs 1.7 at 24 h, # P < 0.05) (Fig 1C). These results indicate that phenothiazines possess a dose-dependent potential to reduce liver damage during pre-engraftment storage.

ALT and AST levels were used to assess mouse liver functional outcomes after progressive cold storage periods with or without phenothiazines, as an index of the state of preservation-related injury. (A) In the 3.6 mg group, a significant but relatively smaller decrease in ALT levels was found at 3 (# P < 0.05) and 24 h (# P < 0.05). ALT levels in the preservation solution were markedly and significantly (## P < 0.01) decreased in the 4.8 mg group at 3, 12, and 24 h after incubation as compared with the control group. (B) Reduced AST levels were also observed following phenothiazine treatment. (C) The AST/ALT ratio was significantly (# P < 0.05) decreased in the 3.6 and 4.8 mg C+P groups at 12 and 24 h.
Morphological Cell Damage
Phenothiazine treatment (4.8 mg) mitigated hepatocellular injury at all time points (3, 12, and 24 h after incubation), maintaining tissue architecture and reducing edema compared with the control group (Fig 2A). The damaged hepatocyte ratio, which was determined, as described above, by the presence of severe swelling and vacuolar degeneration, was therefore also reduced at all time points: 3 (7.8 vs 11.8, # P < 0.05), 12 (15.1 vs 20.0, # P < 0.05), and 24 h (23.7 vs 38.7, # P < 0.05) (Fig 2B).
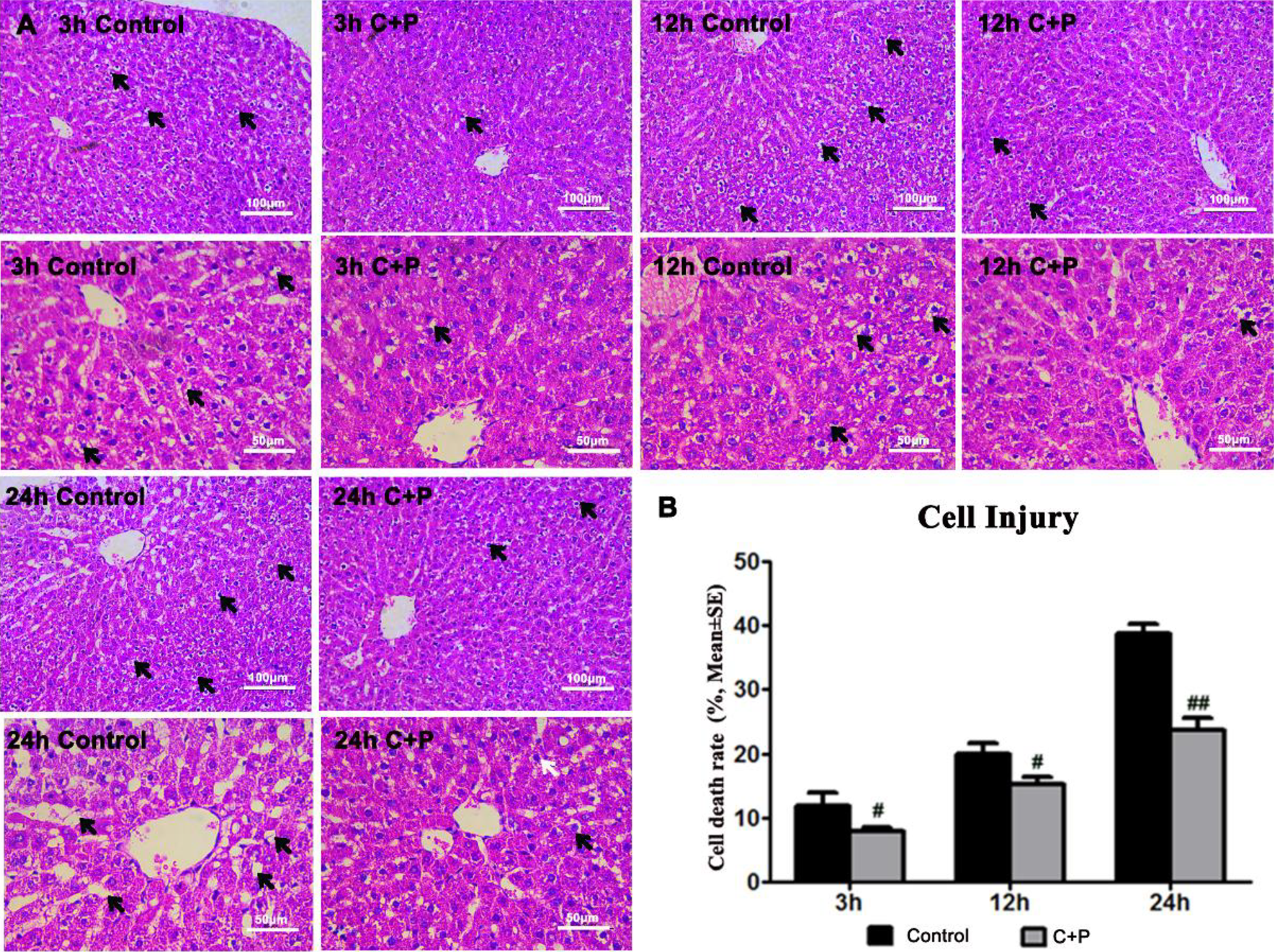
(A) Phenothiazine treatment maintained the tissue architecture and reduced edema compared with the control group when assessed at 3, 12, and 24 h after incubation. (B) The hepatocyte death ratio was significantly reduced by use of phenothiazines at 3 (# P < 0.05), 12 (# P < 0.05), and 24 h (## P < 0.01).
Detection of Apoptosis and Apoptotic Protein Expression
Compared with control groups, cell death was significantly decreased in all groups that underwent C+P incubation (2.5 vs 3.0 at 3 h, # P < 0.05; 2.5 vs 3.1 at 12 h; 2.5 vs 3.5 at 24 h, ## P < 0.01) (Fig 3A). Additionally, Western blot was used to measure protein expression changes attributable to C+P. The expression of pro-apoptotic (BAX) and anti-apoptotic proteins (Bcl-2) were measured at 3, 12, and 24 h after C+P incubation. In the 4.8 mg C+P treatment group, a significant decrease (# P < 0.05) in BAX expression was seen at 3 (1.0 vs 0.6), 12 (1.4 vs 1.2), and 24 h (1.6 vs 1.3) (Fig 3B); similarly, a significant (# P < 0.05) increase in Bcl-2 expression was seen in the 4.8 mg C+P treatment group at 3 (0.6 vs 0.8) and 24 h (1.0 vs 1.2), while an increase that did not reach statistical significance was seen at 12 h (0.8 vs 1.0) (Fig 3C). These findings indicate that C+P may both inhibit pro-apoptotic protein expression, and enhance expression of anti-apoptotic proteins.
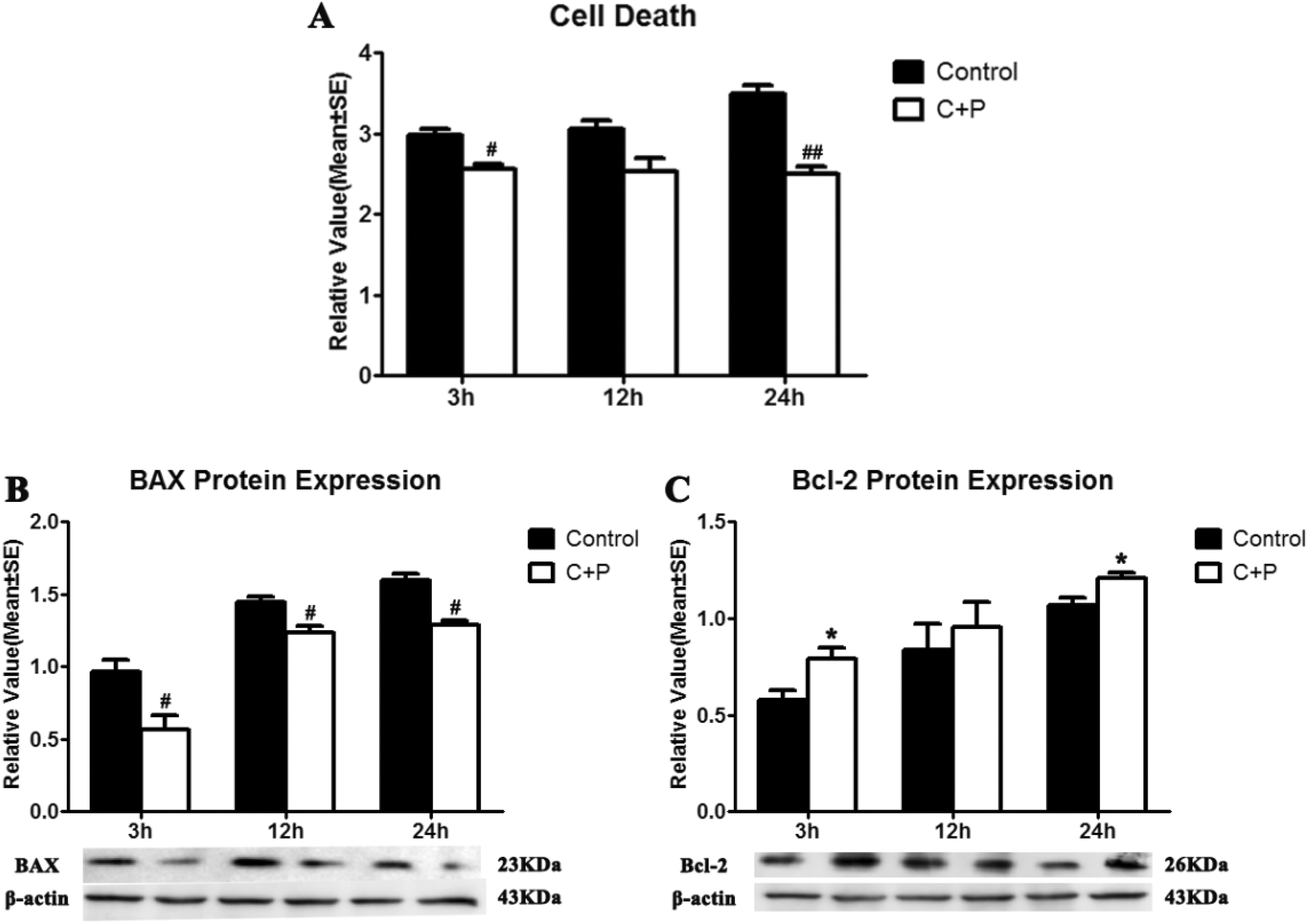
(A) Apoptotic cell death was detected using ELISA. Compared with controls, cell death was decreased at 3 (markedly, # P < 0.05), 12 (slightly), and 24 h (markedly,## P < 0.01). (B) BAX protein expression at 3, 12, and 24 h after C+P incubation was significantly (*P < 0.05) decreased compared with the control group. (C) A significant increase in Bcl-2 protein expression at 3 and 24 h after C+P incubation (*P < 0.05) was also observed, while a slight increase was seen at 12 h. Representative immunoblots are presented.
GLU and LDH Levels
At 2.4 mg, phenothiazine augmentation did not produce any effect on GLU levels (Fig 4A). In the 3.6 mg group, however, a significant decrease was found at 3 (0.9 vs 1.6 unit/L, # P < 0.05), 12 (2.6 vs 4.2 unit/L, # P < 0.05), and 24 h (3.0 vs 5.2 unit/L, # P < 0.05); GLU levels were also significantly decreased in the 4.8 mg group at 3 (0.9 vs 1.6 unit/L, ## P < 0.01), 12 (2.5 vs 4.2 unit/L, # P < 0.05), and 24 h (3.0 vs 5.2 unit/L, # P < 0.05) after incubation. Similar results were observed for LDH (Fig 4B). These results suggest that phenothiazines exert a dose-dependent suppressive effect on brain metabolism.

(A) A significant (# P < 0.05) decrease was found in GLU levels at 3, 12, and 24 h compared with the control group following treatment with 3.6 mg C+P. GLU levels were also significantly decreased by 4.8 mg C+P at 3 (## P < 0.01), 12 (# P < 0.05), and 24 h (# P < 0.05). (B) Similar results were observed with LDH levels, on which C+P was found to have dose-dependent suppressive effects.
ROS, SOD and MDA Levels
In tissue preserved in UW solution augmented with phenothiazines, ROS levels were significantly decreased at 3 (20.1 vs 33.2 mmol/g, # P < 0.05), 12 (81.5 vs 96.1 mmol/g, # P < 0.05), and 24 h (69.6 vs 102.8 mmol/g, ## P < 0.01) compared with UW solution alone (Fig 5A). The activity of SOD in liver tissue treated with phenothiazines (4.8 mg), in contrast, was significantly increased at 3 (136.3 vs 128.5 unit/mg, *P < 0.05), 12 (101.9 vs 93.3 unit/mg, *P < 0.05), and 24 h (106.5 vs 84.9 unit/mg, *P < .05) (Fig 5B). MDA concentration in liver tissue treated with 4.8 mg C+P (6.2 vs 7.9 unit/mg at 3 h, # P < 0.05; 4.3 vs 6.4 unit/mg at 12 h, # P < 0.05; 13.6 vs 31.8 unit/mg at 24 h, ## P < 0.01) was, like ROS levels, significantly reduced (Fig 5C). These results suggest that the augmentation phenothiazines provide to the protective effects of cold liver preservation occurs, at least in part, through inhibition of oxidative stress.
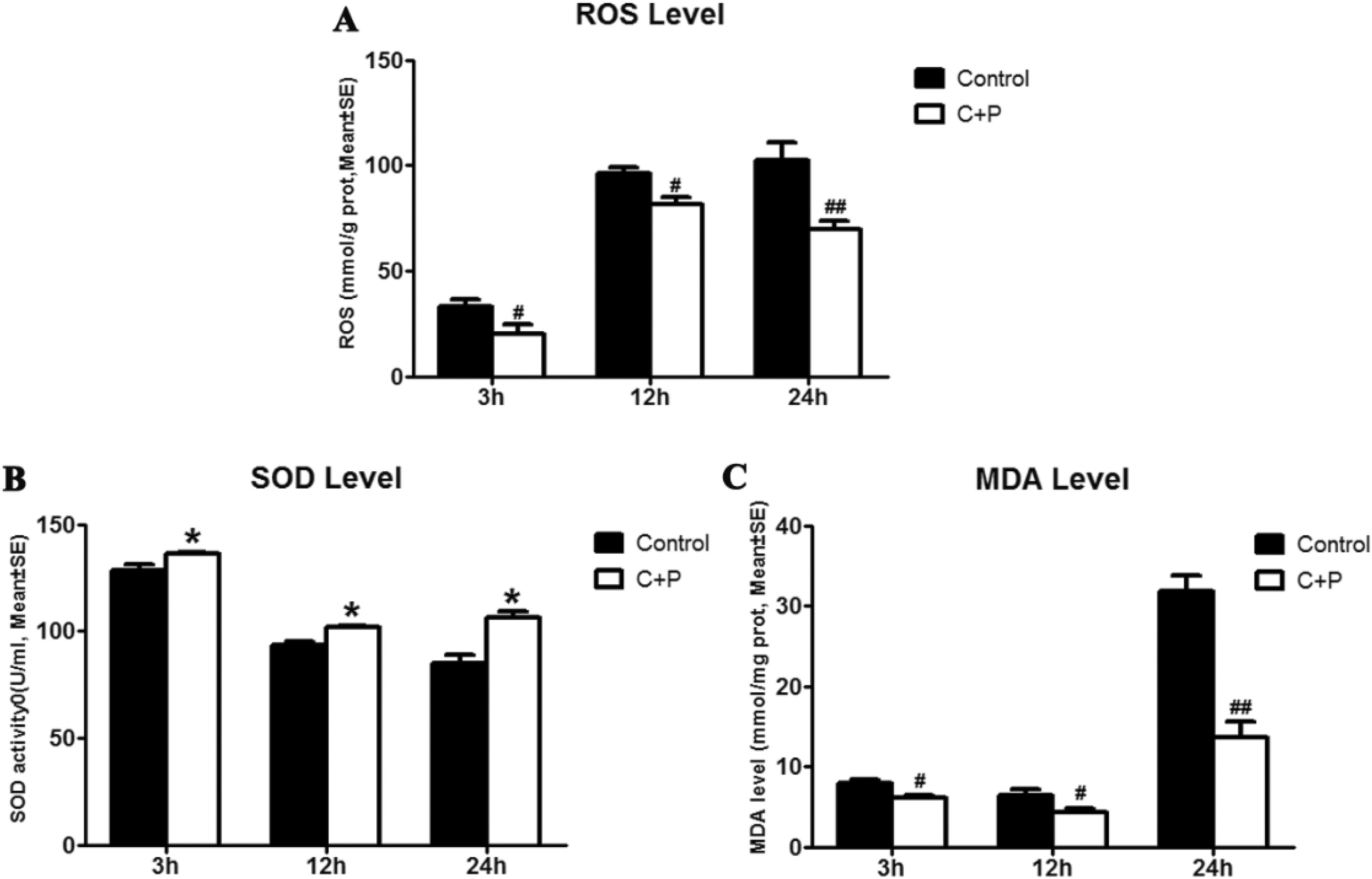
(A) ROS levels were significantly deceased in the phenothiazine treatment group at 3 (# P < 0.05), 12 (# P < 0.05), and 24 h (## P < 0.01), as compared with the control group. (B) SOD activity was compared between the treatment and control group. Phenothiazines significantly (*P < 0.05) elevated SOD activity as compared with the control group at 3, 12, and 24 h after incubation. (C) Phenothiazine treatment significantly decreased MDA levels at 3 (# P < 0.05), 12 (# P < 0.05), and 24 h (## P < 0.01).
Inflammatory Protein Expression
To investigate whether an anti-inflammatory action also contributes to the augmentation exerted by phenothiazines, we compared expression of the pro-inflammatory cytokines TNF-α and IL-1β in tissue preserved with phenothiazines plus UW solution with expression in the presence of UW solution alone. In the 4.8 mg C+P treatment group, a significant decrease (# P < 0.05) in IL-1β expression was seen at 24 h (1.3 vs 0.8), while slight decreases were seen at 3 (0.6 vs 0.5) and 12 h (1.3 vs 1.1) (Fig 6A). Similarly, a significant (# P < 0.05) decrease in TNF-α expression was seen in the 4.8 mg C+P treatment group at 24 h (1.3 vs 1.1), while decreases that did not reach statistical significance were seen at 3 (0.7 vs 0.6) and 12 h (0.9 vs 0.6) (Fig 6B). These findings indicate that C+P may exert an inhibitory effect on the expression of pro-inflammatory cytokines.

(A) IL-1β protein expression was significantly (# P < 0.05) decreased compared with the control group at 24 h after C+P incubation, while slight decreases were seen at 3 and 12 h. (B) TNF-α protein expression was significantly (# P < 0.05) decreased compared with the control group at 24 h after C+P incubation, while slight decreases were seen at 3 and 12 h. Representative immunoblots are presented.
Discussion
In this study, we demonstrated that phenothiazine augmentation enhanced liver graft preservation by UW solution, as demonstrated by reductions in histologically determined tissue damage, hepatic enzyme leakage, glucose metabolism, and oxidative injury. To our knowledge, this is the first study to demonstrate these effects; it is also the first study to describe their underlying anti-inflammatory and antioxidant mechanisms.
The phenothiazines with which these effects were achieved, chlorpromazine and promethazine, have been widely used, and often in combination, for their antipsychotic and sedative effects 5,6 . Chlorpromazine is a low-potency typical antipsychotic with primarily anticholinergic and antiadrenergic side effects 23 . Promethazine is a first-generation antihistamine clinically used for its strong sedative, antiemetic, and weak antipsychotic effects 24 . In addition to these more widely-known effects, however, the combination of these two drugs has been demonstrated to induce a hypometabolic state that has been referred to as artificial hibernation 25 , in the setting of which our group observed a dose-dependent neuroprotective effect following both transient and permanent ischemia 7,9,26 . Thus, having found doses in our previous work that produced the optimal balance of safety and cytoprotective efficacy 9 , we set out to demonstrate that the techniques that produced neuroprotection from ischemic stroke could translate into improved hepatocellular preservation from transplant-related ischemia.
In clinical liver transplantation, prolonged durations of cold ischemic preservation are associated with primary dysfunction and loss of grafts. Although the cause of this dysfunction is likely multifactorial (other potential causes include steatosis, prolonged donor warm ischemia, and poor organ flushing) 27 , any achievable reduction in injury incurred during preservation should limit the risk of dysfunction, and thus make better use of available livers. Unfortunately, in the three decades following the advent of the UW organ flush solution in 1987, there has been no further significant progress in minimizing the incidence of hypothermic liver preservation injury 28 . UW solution protects liver cells by decreasing energy depletion and lipid peroxidation 29 , and allows preservation of donor livers for up to 12–18 h in clinical settings 30 . In this study, augmentation of UW solution by phenothiazines was shown to constitute the advance in graft preservation efficacy that has been elusive in recent history. Phenothiazine augmentation reduced cell damage by suppressing oxidative stress, inflammation, and brain metabolism, as determined by measurements of cell morphology, apoptosis, BAX, Bcl-2, ROS, SOD, MDA, ALT, AST, GLU, LDH, IL-1β, and TNF-α.
The results of these measurements accord with previous research regarding the metabolic effects of phenothiazines. Previous studies have demonstrated that mitigation of ischemic injury is one of the mechanisms whereby phenothiazines exert their protective effect 7 . Ischemic injury, through hypoxic inhibition of oxidative phosphorylation, generates an overproduction of ROS 31 ; ROS-induced oxidative damage has been shown to contribute to graft injury 32 . In the present study, elevated SOD and decreased MDA suggest that phenothiazine-induced reductions in ischemic injury exert an enhanced antioxidant effect with positive ramifications on graft quality. ROS levels are dependent on the ratio of their production to their clearance, which is regulated by various cellular antioxidant compounds, including SOD 33,34 . In basic and clinical research, SOD levels are thus used as an indicator of free radical scavenging capacity 35 . MDA, an indicator of lipid peroxidation, is measured for the converse purpose of demonstrating the extent of free radical-mediated cellular damage 36,37 . In addition to the oxidative mechanism, ischemic hepatocellular injury has also been shown to be associated with inflammation, and in particular with elevated levels of the pro-inflammatory cytokines TNF-α and IL-1β 38 . In previous work, our group has demonstrated the capacity of phenothiazines to reduce post-stroke inflammation; likewise, the reductions in TNF-α and IL-1β observed in this study further suggest an anti-inflammatory mechanism for phenothiazine-mediated cytoprotection.
Further work conducted by our group and others suggests a role for the endoplasmic reticulum (ER) in mediating both the inflammatory and oxidative mechanisms of cellular injury observed in this study. Specifically, the protein constituents of the ER stress (ERS) signaling pathway have been found to be upregulated in traumatic brain 39 and liver 40,41 injury models, while expression of TNF-α and ROS has been found to be decreased when ERS was inhibited 42 . A growing body of evidence suggests that there is extensive cross-talk between the inflammatory response and ERS 43 : several stimuli (lipids, cytokines, or injury) appear to activate ERS, which then further disrupts metabolic functions, causing more inflammation 44 . Mechanistically, the relationship between inflammation and ERS may include the release of calcium from the ER 45 , activation of the transcription factor nuclear factor-κB (NF-κB) 46 , mitogen-activated protein kinase (MAPK) 47 , induction of the acute phase response, and the production of ROS 48 . Studies have also linked ERS to the generation and accumulation of ROS, suggesting concomitant involvement with oxidative stress 48,49 . Because of the importance of the inflammatory and oxidative mechanisms suggested by the results of the present study, we intend in future work to investigate whether ERS attenuation contributes to C+P-mediated augmentation of liver graft preservation.
Conclusion
Despite improvements made by the use of UW solution, considerable temporal limitations continue to curtail the success of liver transplantation in both basic scientific and clinical settings. It is this circumstance that foregrounds the utility of the results obtained in this study, in which we demonstrated in vitro that augmentation of UW solution with phenothiazines provided improved protection of hepatocytes against ischemia-induced oxidative stress. In order to see whether these findings can translate into improvements in clinical efficacy, further studies in vivo that assess the effects of phenothiazine supplementation on organ preservation and transplant survival in whole animals are warranted.
Footnotes
Ethical Approval
The clinical study was approved by the Institutional Animal Investigation Committee of the Capital Medical University.
Statement of Human and Animal Rights
All of the experimental procedures involving animals were conducted in accordance with the Institutional Animal Investigation Committee of the Capital Medical University.
Statement of Informed Consent
Statement of Informed Consent is not applicable for this article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was partially supported by the National Nature Science Foundation of China (81871838 and 81802231), the China-Japan Friendship Hospital Medical Development Research Foundation Project (2013-RC-2), the Beijing Tongzhou District Financial Fund, and the Science and Technology Plan of Beijing, Tongzhou District (KJ2018CX006-02).