
Abstract
To investigate the role of dopamine release in cognitive impairment and motor learning deficits after brain injury, different levels of traumatic brain injury (TBI) were made in rats by using fluid percussion at two different atmospheres (2 Psi and 6 Psi). Tonic and phasic bursting dopamine release and behavior tests followed at several time points. We used in vitro fast-scan cyclic voltammetry to survey dopamine release in the striatum and analyzed the rats’ behavior using novel object recognition (NOR) and rotarod tests. Both tonic and bursting dopamine release were greatly depressed in the severely (6 Psi) injured group, which persisted up to 8 weeks later. However, in the 2 Psi-injured group, the suppression of bursting dopamine release occurred at 1~2 weeks after injury, but there were no significant differences after 4 weeks. Tonic dopamine release was also diminished significantly at 1~2 weeks after the injury; partial recovery could then be seen 4 weeks after injury. A significant deficiency in the fixed speed rotarod test and NOR test were noted in both 2 Psi and 6 Psi groups initially; however, the changes recovered in the 2 Psi group 2 weeks after injury while persisting in the 6 Psi group. In conclusion, striatal evoked dopamine release was affected by fluid percussion injury, with behavioral deficits showing differences as a function of injury severity. The severe fluid percussion injury (6 Psi) group showed more dopamine release defects, as well as cognitive and motor deficiencies. Recovery of dopamine release and improvement in behavioral impairment were better in the mild TBI group.
Keywords
Introduction
Traumatic brain injury (TBI) is a heterogeneous and complex condition composed of acute, subacute, and chronic pathologies (27,35). Cognitive impairment, and learning and memory deficits, after head injury have been noted for years. Different degrees of head injury cause different levels of central nervous system damage and neurological symptoms. Mild TBI, often referred to as “concussive,” is known to produce symptoms like nausea, dizziness, intermittent headache, and cognitive problems in at least 15% of patients, while more severe brain injury may cause brain parenchymal loss, and some estimates suggest that 50% of these individuals show long-term injury-related disabilities (28). Animal models of brain injury, including controlled cortical impact (CCI) (12,29) and fluid percussion (FP) (13), have provided insights into the cellular and molecular mechanisms of central nervous system (CNS) dysfunction and cell death. TBI-related impairments in behavior and changes in the transcriptome of certain brain areas are known to occur in mild TBI (2,37,48). A role for dopamine (DA) in these postinjury sequela has been suggested, but how DA system function changes after injury and how such changes influence behavior have not been fully defined.
The purpose of this study was to compare the effects of different degrees of brain injury on rat cognitive and motor performance and to analyze sequential changes in striatal DA release after fluid percussion injury. The striatum was selected due to its role in procedural learning and cognition. Behavioral changes and DA release dynamics were examined at different times following injury.
We divided the animals into mild (PSI-2 Psi) and severe (PSI-6 Psi) cortical injury groups. We carried out behavioral tests including novel object recognition (NOR, for cognition) and rotarod (for motor skills) sequentially and analyzed DA system function by using in vitro electrochemical methods [high-performance liquid chromatography (HPLC) and fast-scan cyclic voltammetry (FSCV)] at different times (2 h, 24 h, 1 day, 7 days, and 8 weeks after cerebral cortical injury) to determine the relationship between DA dynamics and the behavioral symptoms of cortical injury in each group. We now report that DA release and uptake were suppressed at 1 week after injury, and the behavioral tests revealed significant differences between the sham and injured animal groups as early as 2 h after injury.
Materials and Methods
Animals
Young adult male Sprague–Dawley rats (LASCO Taiwan Co., Ltd., Taipei City, Taiwan) were used with all procedures approved by the National Defense Medical Center (NDMC) Animal Care and Use Committee. Animals received a fluid percussion injury (n = 101) or were sham controls (n = 21). Animals were provided food and water ad libitum and were housed in a 12-h light– dark cycle room.
Surgical Preparation and Fluid Percussion Model
Male SD rats (6 weeks old) weighing 200–250 g were anesthetized with chloral hydrate (4 mg/100 mg; Tokyo Chemical Industry Co., Ltd., Tokyo, Japan). With the animals in a stereotaxic frame (Mod 51600; Stoelting Co., Wood Dale, IL, USA), the scalp and temporal muscle were reflected. A craniectomy of 4.8 mm in diameter was performed over the right parietal cortex, 3.8 mm posterior to the bregma, and 2.5 mm lateral to the midline, taking care not to penetrate the dura (10). A cranial Leur adapter (HS-NY-M-2H; Dragonfly Research & Development, Inc., Ridgeley, WV, USA) of 2.5 mm in inner diameter was placed on the craniectomy site and tightly mounted to the skull using dental acrylic resin (Lang Dental Manufacturing Co., Inc., Wheeling, IL, USA). The cranial Leur adapter was filled with saline and attached to the fluid percussion device. The fluid percussion device (model HPD-1700; Dragonfly Research & Development) was used to produce TBIs in rats as described by Matsushita et al. (31). Injury was induced by striking the piston with a weighted metal pendulum released from a predetermined angle (2 Psi 16° and 6 Psi 43°; Pa means pounds per square inch). The resulting rapid injection of a small volume of saline into the closed cranial cavity caused a pulse of increased intracranial pressure that was associated with a deformation in the brain. Pressure pulses were measured extracranially with a pressure transducer, recorded on a digital real-time oscilloscope (TDS210; Sony Tektronix Corp., Osaka, Japan), analyzed by WaveStar software (Sony Tektronix Corp.) and, based on prior instrument calibration, are expressed in pounds per square inch (Psi). The fluid percussion device delivered transient pressure fluid pulses with constant wave form and duration (17–21 ms) to cause brain injuries (42). Animals were subjected to sham (control, n = 21), low (1.9 ± 0.2 atm, n = 50), or high (6.0 ± 0.5 atm, n = 51) fluid percussions.
FSCV Recordings
Carbon fibers (7 μm diameter; Goodfellow Corp., Oakdale, PA, USA) were prepared as described previously (7,18). Briefly, micropipettes containing the carbon fibers were backfilled with a 4 M potassium acetate/150 mM KCl solution and connected to a standard patch pipette holder/ headstage assembly. CHEM-CLAMP (cornerstone series; Minneapolis, MN, USA). A voltammeter– amperometer (Dagan Corporation, Minneapolis, MN, USA) was used to change the carbon fiber electrode potential and to measure current. Voltammetric scans and electrical stimulus timing protocols were performed using PCI-based analog to digital boards (National Instruments, Austin, TX, USA) and LabView-based software (courtesy of Dr. Mark Wightman, University of North Carolina, Chapel Hill, NC, USA). During electrochemical detection, the potential of the carbon fiber was driven from –0.4 to 1.0 V and back to –0.4 V, using a triangular waveform (400 V/s, 7-ms duration) applied every 100 ms. A 5-s (50-scan) control period was used to obtain a stable background current that was digitally subtracted from that obtained during the peak of the response following electrical stimulation of tissue. Peak oxidation currents were converted to dopamine (DA) concentrations using a calibration performed for each electrode with a 1-μM DA standard solution. All signals used in the statistical analyses matched the expected voltammetric profile for DA (25).
Electrically Evoked DA Signals in Brain Slices
Under stereoscopic magnification, carbon fibers were lowered to a depth of 100 μm into the dorsolateral striatum. A bipolar stimulating electrode was positioned 75–100 μm from the carbon fiber, and constant voltage pulses (1–20 Volt, 1-ms duration) were delivered between voltammetric scans to elicit DA release. Responses were obtained every 2 min, and all those used for analysis were stable throughout the duration of the recordings. For single-pulse experiments, DA uptake was assessed by fitting a single exponential function to the signal decay using a least-squares minimization algorithm (WinWCP; Dr. John Dempster, Strathclyde Institute for Biomedical Sciences, Glasgow, UK; http://spider.science.strath.ac.uk). A tau (t) value was obtained for each recording site by averaging all time constants obtained from each DA signal generated during input/output curves (stimulus intensity vs. DA signal). The first-order rate constant (k or 1/τ) obtained using this approach provides an index of the efficiency (Vmax/Km) of DA clearance mediated by the dopamine transporter (DAT) (7,39).
To assess the capacity of axon terminals to release DA during burst stimulation, three voltammetric signals were obtained at each recording site using a single pulse and 2, 5, and 10 pulses delivered at 25 Hz. After three signals at each site were averaged, the difference between the peak DA signal obtained immediately after burst stimulation or single pulses was determined as DAnp – DA1p, where DAnp is the amplitude of the voltammetric signal for n pulses, and DA1p is the amplitude of the voltammetric signal obtained following a single electrical pulse. Data were obtained for each slice before and during drug treatment. The DAT inhibitor nomifensine was used to determine the role of uptake in the regulation of frequency-dependent DA release in control and 6 Psi-injured rats. The data were fit to a linear regression model (y = mx + b; Prism 5.02; GraphPad, San Diego, CA, USA), where the slope m represents the relative change in DA concentration per pulse (18).
High-Pressure Liquid Chromatography (HPLC)/ Electrochemical Detection
Rats from different groups were sacrificed at fixed times, and the brains were then rapidly removed. The striatum were dissected out and frozen immediately on dry ice. On the day of assay, the dissected tissue was homogenized in 0.1 mM oxalic acid (Nacalal Tesque, Inc., Osaka, Japan). The homogenates were centrifuged at 15,000 × g for 40 min at 4°C. The resulting supernatants were filtered through a 0.22-μm syringe filter (Millipore, Bedford, MA, USA) followed by HPLC to determine the concentrations of DA, 3,4-dihydroxyphe-nylacetic acid (DOPAC), and homovanillic acid (HVA). The HPLC system was composed of a reverse-phase C-18 column (MD-150, RP-C-18, 3 mm, length: 15 cm; ESA Biosciences, Chelmsford, MA, USA) and a high-pressure pump (LC-10AD; Shimadzu, Kyoto, Japan); these were connected to an electrochemical detector (ECD) coupled with three electrodes (Coulochem II; ESA Biosciences). The electrode of the guard cell was set at 40 mV, and electrodes 1 and 2 (for detection) were set at 250 and 350 mV, respectively. Under an isocratic condition, the mobile phase solvent (MD-TM; ESA Biosciences) was circulated at a flow rate of 0.5 ml/min in the system. To quantify the sample peaks, each chemical species (DA, DOPAC, and HVA) was compared to the external standards, which were freshly prepared and injected every five-sample run as we described previously (23). The turnover rates of DA [(DOPAC + HVA)/DA] were calculated to represent the activity of DA neurons.
Behavioral Tests
Rotarod Test
Rats were trained at three times/trial weekly using a rotarod (Acceler rota-rod for rats 7750; Ugo Basile, Comerio-Varese, Italy) with a fixed rate of 4 rpm before the injury. We tested motor function five times/trial at 24 h before and after the injury, and subsequent tests were performed every week after injury for 8 weeks.
NOR Test
Methods for NOR tests were modified from those originally reported by Ennaceur and Delacour (15). An acrylic box (50 × 50 × 50 cm) consisting of four black walls and one white floor was used. The chamber was equipped with an overhead camera for recording. The tests were performed in an isolated room free from noise. The NOR tests were carried out on 1 day preinjury and 1, 2, 4, 6, and 8 weeks postinjury. During the first NOR test, familiar objects (F) used in the first and second trial were gray plastic egg-shaped colors, whereas the novel object (N) in the second trial was a white glass cube-shaped color. All the objects for the rats to explore were within a certain weight, which controlled for movements caused by rats. The time that the animal spent on exploring the familiar and novel objects within 5 min was recorded and converted to the percentage of the total exploration time. The exploration time of (novel minus familiar)/total time was also calculated. To consider the preference of the rats between the novel and familiar objects, the discrimination index (DI) defined as DI = (novel object exploration time/total exploration time) - (familiar object exploration time/total exploration time) × 100 was used.
Western Blot Analysis of Tyrosine Hydroxylase (TH) Phosphorylation
After anesthesia, animals were decapitated and the brains quickly removed and placed on ice. The striatum was excised and frozen in liquid nitrogen. Tissue was stored at -80°C until used for analysis. Striatal tissue was homogenized in 300 μl of RIPA lysis buffer containing phosphatase inhibitor cocktail (Cell Signaling, Danvers, MA, USA), and protease inhibitor cocktail (Bionovas Biotechnology Co., Ltd., Toronto, Ontario, Canada) was added to samples used for Western blot. Protein concentrations were determined using a BCA protein Assay Kit (Biotools Corporation, Taipei City, Taiwan). Samples containing 10 μg of protein were separated by 10% SDS-PAGE (Biotools Corporation) and were subsequently electrophoretically transferred onto polyvinylidene difluoride blotting membranes (PVDF; Pall Corporation, Port Washington, NY, USA). PVDF were blocked with 5% bovine serum albumin (Sigma-Aldrich, St. Louis, MO, USA) in tris-buffered (Bioman Corporation, Tai Pei, Taiwan) saline with 0.05% tween 20 (Biokit Biotechnology Incorporation, Hsinchu, Taiwan) (TBST). Following blockade, membranes were immunolabeled with antibodies [1:1,000 p-TH ser40 (Cell Signaling); 1:1,000 dopamine transporter, 1:1,500 TH, 1:20,000 β-actin (Novus Biological LLC, Littleton, CO, USA); 1:1,000 p-TH ser19 (Millipore)] in TBST for 16–20 h at 4°C. Following primary antibody incubation, membranes were washed 6 × 5 min in TBST and incubated for 1 h in goat anti-rabbit immunoglobulin G conjugated to peroxidase (1:5,000; Epitomics) at room temperature. Finally, the membrane was washed 6 × 5 min in TBST. Proteins were visualized with a chemiluminescence (UVP Upland, CA, USA) detection system, and protein bands were quantitated with ImageJ software (NIH, Bethesda, MD, USA).
Statistical Analyses
Data in the text and figures are expressed as means ± SEM. Statistical analyses of data for the dopamine release input/ output curves and behavioral tests were performed using a two-way analysis of variance (ANOVA) followed by a Bonferroni post hoc test for multiple comparisons. A one-way ANOVA and a Bonferroni post hoc test were used to determine changes in t values and HPLC data for the injured and control groups. For DA-releasing probability experiments, we analyzed the group differences by using analysis of covariance (ANCOVA) testing followed by Student– Newman–Keuls (SNK) testing for multiple comparisons. Mixed-effects regression analysis for repeated measures was used to evaluate group differences for evoked DA release in the striatum. All statistical tests were two tailed and were performed using GraphPad Prism 5.02 (GraphPad Scientific, San Diego, CA, USA). A value of p < 0.05 was considered significant for all analyses. Group differences in protein expression were evaluated using two-tailed independent t-tests, with band density of injured groups reported as a percent of control.
Results
The Extent of Suppression of DA Release in the Striatum Varied with Severity of Fluid Percussion Injury
To investigate the sequential effect of cerebral cortical injury on the striatal DA release and the relationship between the severity of injury and the duration on the effect of DA release, we compared the evoked DA signals of dorsal striatum after 2 Psi injury (Fig. 1) and 6 Psi injury group (Fig. 2) by using FSCV at different time points (2 h, 24 h, 1, 2, 4, 6, and 8 weeks postinjury).

The suppression of DA release after 2 Psi FPI. The representative voltammetric signals evoked by 5 V stimulation intensity, obtained from control and injured animals at sequential time points, are shown in (A1–H1; IT curve), while (A2–H2) show that cyclic voltammograms obtained at the peak current are consistent with DA in each case (CV curve). Evoked DA release by single pulse stimulation that mimics tonic DA release (1 pulse/25 Hz, arrow) at each of the following time points after injury (A: Control, B: 2 h, C: 24 h, D: 1 week, E: 2 weeks, F: 4 weeks, G: 6 weeks, and H: 8 weeks after 2 Psi FPI). The DA release signal by 10 pulses at 25 Hz that mimic phasic bursting release are shown in (J–Q), at the same sequential times as tonic release (J1–Q1: IT curve; J2–Q2: CV curve). A decrease in signal occurs initially after injury and declines further at subacute stages (postinjury 1 week and 2 weeks). Input/output (I/O) curves for single pulse (I) and 10 pulses (R) 25 Hz stimulation/elicitation in brain slices obtained from control and injured animals at different stimulation intensities (from 1 V to 10 V), which indicate that DA release recovers by 4 weeks (open triangles) until 8 weeks (open circles) after 2 Psi FPI. The I/O curve of both tonic (evoked by 1 p/25 Hz stimulation) and phasic (evoked by 10 p/25 Hz stimulation) DA release decreased in the injured animals at 2 h, 1, 2, 4, and 6 weeks from 5 to 10 stimulus intensity (V). However, p < 0.05 in control versus 8 weeks postinjury under 7-, 9-, and 10-V stimulation. (S) The peak value of DA tonic (1 P) and bursting (10 P) release evoked by 5 V/25 Hz stimulation at each time point after 2 Psi FPI also indicates that reduction of the signal occurred initially and then recovered by 4 weeks after injury. *p < 0.05, **p < 0.01, ***p < 0.001.

A greater suppression of DA release is found in the 6 Psi FPI animal. The evoked DA signal obtained from striatal slices in control and injured animals at different time points after a single pulse/25 Hz (1 p, A1–H1: representative voltammetric signals and A2–H2: representative cyclic voltammograms at 5 V stimulation intensity). The signals evoked by 10 pulses/25 Hz (10 p) are shown in (J1–Q1: IT curve) and (J1–Q2: CV curve). A summary of evoked DA release with varying stimulation intensities at different postinjury times (input/output; I/O curve) are shown in (I) and (R) at 1 and 10 P, respectively, which indicate that marked suppression occurs in the severe (6 Psi FPI) injury group. This release suppression persisted until 8 weeks (open circles). (S) The peak values of DA tonic (1 P) and bursting release (10 P) signals evoked by 5 V/25 Hz stimulation at each time point after 6 Psi FPI are shown; the signal is still reduced markedly after 8 weeks. *p < 0.001.
The signals for evoked DA release after a single pulse, which mimics tonic DA release, and 10 pulses, which mimics phasic bursting release in striatum were recorded. The representative voltammetric signals evoked by 5 V stimulation intensity, obtained from control and injured animals at different time points are shown in Figure 1A1–H1 (IT curve), while Figure 1A2–H2 shows cyclic voltammograms obtained at the peak of the current consistent with DA in each case (CV curve). The evoked DA release after injury decreased initially and further deteriorated at a subacute point (postinjury 1 week and 2 weeks) and then increased after 4 weeks in the 2 Psi injury group. Input/output curves for single-pulse (Fig. 1I) and 10-pulse (Fig. 1R) at 25 Hz stimulation/elicitation in brain slices obtained from control and injured animals revealed that tonic DA release (evoked by 1 p/25 Hz stimulation) decreased in the injured animals [F(63, 499) = 1.96, p < 0.001] according to a two-way ANOVA followed by Bonferroni posttests [all p < 0.001 in control vs. postinjury groups at 2 h, 1, 2, 4, and 6 weeks from 5 to 10 stimulus intensity (V); however, p < 0.05 in control vs. 8 weeks postinjury under 7-, 9-, and 10-V stimulation]. Bursting release (evoked by 10 p/25 Hz stimulation) deficits tend to return to near-normal range 4 weeks postinjury in the 2 Psi group [F(63, 607) = 2.77, p < 0.001] according to two-way ANOVA followed by Bonferroni posttests [all p < 0.01 in control vs. postinjury groups at 2 h, 1, 2 weeks postinjury from 5 to 10 stimulus intensity (V), p < 0.05 in control vs. 4 and 8 weeks postinjury only at high stimulation intensities (8~10 V), but all p > 0.05 in control vs. 6 weeks postinjury]. However, in the 6 Psi-injured group, the low DA release persisted until 8 weeks postinjury with both tonic dopamine release evoked by 10 p/25 Hz stimulation [F(63, 639) = 7.315, p < 0.001] according to two-way ANOVA followed by Bonferroni posttests [all p < 0.001 in control vs. postinjury groups at 2 h, 1, 2, 4, 6, and 8 weeks from 3 to 10 stimulus intensity (V)] (Fig. 2A–H), and phasic bursting (10 p/25 Hz stimulation) shown in Figure 2J–Q [F(63, 692) = 6.224, p < 0.001] according to two-way ANOVA followed by Bonferroni posttests [all p < 0.001 in control vs. postinjury groups at 2 h, 1, 2, 4, 6, and 8 weeks from 4 to 10 stimulus intensity (V)].
The DA signals with maximal stimulation intensity (10 V) at different times after injury are summarized graphically in Figure 3. The dopamine signals in the 2 Psi group were decreased for 2 weeks after single and multiple pulse stimulation but returned to normal levels by 4 weeks (Figs. 1 and 3A). The DA signals for both single and 10-pulse stimulation were also markedly reduced in the 6 Psi group. However, in contrast to the 2 Psi, the reduction persisted for at least 8 weeks (Figs. 2 and 3B). Thus, our data indicated that the striatal dopamine release was affected by fluid percussion injury to the cortex, and the extent of release suppression was related to the severity of the injury.
The DA signals with maximal stimulation intensity (10 V) at times after injury are shown in (A; 2 Psi group) and (B; 6 Psi group). In the 2 Psi group, the DA signal evoked by single pulses decreased at 2 h after injury, and the signal evoked by 10 pulses decreased at 1 week after injury (A). A significant decrease could be found in DA signals evoked either by single pulses or 10 pulses in the 6 Psi-injured group (B). The signal in the chronic state (4 weeks after injury) increased to almost control levels in the 2 Psi group, but this could not be found in the 6 Psi group. (Significance is given as *p < 0.05; **p < 0.01; ***p < 0.001 compared with control.) The evoked DA release with 5 V (C) and 10 V (D) stimulation after injury in each group is shown, which revealed more significant suppression in the 6 Psi group, especially with phasic release (p < 0.05).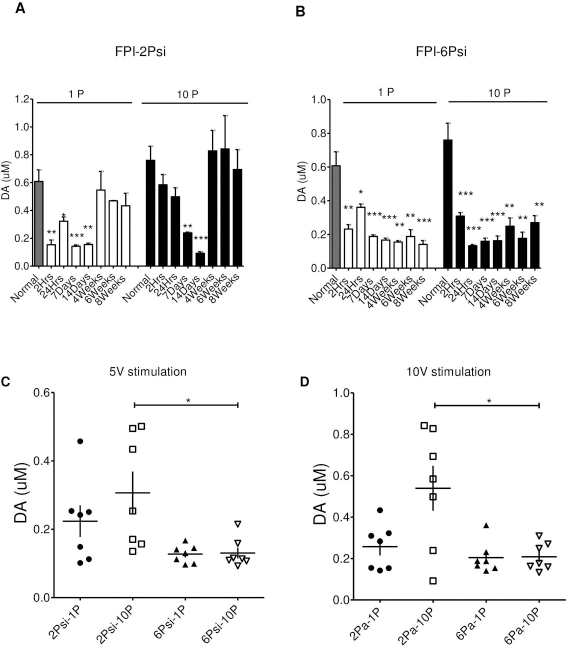
The Reuptake System of Dopamine Was Affected by Head Injury; t Value Was Prolonged by Head Injury Especially at Subacute Stages in Either the Mild (2 Psi) or Severe (6 Psi) Group
We analyzed the clearance rate for dopamine in the striatum by comparing decay time constants (t) after cortical injury. Figure 4 shows decay time constants (t) after injury in different injury groups. At value was obtained for each recording site by averaging all time constants obtained from each DA signal generated during input/output curves (stimulus intensity vs. DA signal). The first-order rate constant (k or 1/t) obtained using this approach provides an index of the efficiency (Vmax/Km) of DA clearance mediated by DAT. In noninjured animals, there was no effect of single or multiple pulses on clearance (Fig. 4A). The clearance rate of DA was prolonged (increased) at 1 week postinjury in both 2 Psi (Fig. 4B) and 6 Psi groups (Fig. 4C). In the 2 Psi group, the time constant increased significantly at 1 week and decreased to normal level by 4 weeks. However, in the 6 Psi group, the time constant was prolonged at 1 and 2 weeks after injury, but reversed to shorter than control values 8 weeks later (p < 0.05). This may indicate that the dopamine clearance rate in striatum is increased at this stage.

Summary of decay time constants (t). At value was obtained for each recording site by averaging all time constants obtained from each DA signal generated during I/O curves (stimulus intensity vs. DA signal). The first-order rate constant (k or 1/t) obtained using this approach provides an index of the efficiency (Vmax/Km) of DA clearance mediated by DAT. The t value was not affected by the stimulation pulse in control animals (A; one way ANOVA, p = 0.9436). A one-way ANOVA revealed significant differences (p < 0.01) comparing each time point with control groups. In the 2 Psi group (B), the time constants tend to increase (significant at 2 weeks after injury with single pulse stimulation, p < 0.05) by 1 day until 2 weeks later, and then return to normal by 4 weeks after injury. However, in the 6 Psi group, the time constants were prolonged significantly at 1 and 2 weeks (C) after injury, and reversed to shorter than control values 8 weeks later. (Significance is given as *p < 0.05, compared with control.) (D) HPLC analysis after injury in acute (2 h to 24 h), subacute (1 and 2 weeks after injury), and chronic (8 weeks after the injury) time points. The elevated metabolite levels were observed at 8 weeks after 6 Psi injury. (E) The absolute concentration of striatal dopamine. (F) Summary of the relationship between the observed changes in DA concentration per pulse in slices from control and 8 weeks after 6 Psi injury (n = 3 slices of three rats each group). Slopes of the regression lines from slices of 8 weeks after 6 Psi injury (black squares) were significantly reduced, relative to controls (open circles) (p < 0.05; one-way ANOVA, Tukey's post hoc). (G) In the presence of nomifensine, the slope of the regression line increased significantly in slices from the 6 Psi injury group. *p < 0.05, ***p < 0.001.
The Effect of Metabolism and Releasing Probability of the Dopamine in Striatum Caused by Fluid Percussion Injury
To measure the metabolism of DA and the activity of DA neurons, we performed HPLC analysis. A decrease of the turnover rate was found initially in 6 Psi injured animals, but this was not significant. Then a significantly increased turnover rate was seen at 8 weeks in the 6 Psi group (Fig. 4D). There were no significant changes of turnover rate of dopamine in either acute or chronic stages in 2 Psi-injured animals (Fig. 4D). We also measured the absolute concentration of dopamine in the striatum ipsilateral to the injury, and no significant difference was found while comparing controls to 2 Psi- or 6 Psi-injured groups (Fig. 4E, ANOVA, p > 0.05).
The reduction in DA release observed with single-pulse stimulation and FSCV is consistent with the lower levels of evoked DA release in injured animals described above. However, DA release dynamics differ according to tonic (4 Hz) or phasic (25 Hz) firing patterns of midbrain DA neurons, and these patterns of activity are related to motivated behavior (20,40). Therefore, we also evaluated DA release by comparing peak concentrations elicited by single and multiple stimuli, delivered at 25 Hz (see Materials and Methods). There was a linear increase in DA concentration as a function of pulse number (Fig. 4F, G). The 6 Psi-injured rat striatal slices demonstrated a significant reduction in DA concentration per pulse, relative to control animals (Fig. 4F, injured rat slope: 19.2 ± 6.3 nM/pulse vs. control rat slope: 42.9 ± 5.3 nM/pulse, n = 3; p < 0.05, Tukey's post hoc). To determine the role of uptake in the regulation of frequency-dependent DA release in control and 6 Psi-injured rats, the DAT inhibitor nomifensine was used. Nomifensine (5 μM) greatly increased the frequency-dependent DA signal in striata of control rats (Fig. 4G, injured rat slope: 169.2 ± 2.3 nM/pulse vs. control rat slope: 117.4 ± 2.2 nM/pulse). Thus, our data indicate that dopamine reuptake in striatum was affected by the FPI with both mild (2 Psi) injury and severe (6 Psi) injury at acute (within 1 week after injury) and subacute (1 to 2 weeks after injury) stages. Subsequently, clearance rates return to normal after 4 weeks in the mild injured group. The shortening of t values in the 6 Psi group at 8 weeks suggests that some recovery of the dopaminergic terminals in striatum at this chronic stage occurs even with severe injury. Taken together, these data suggest that DAT greatly limits the size of the electrically released DA signal in striatal slices from control rats and underscores the extent of the decrease in DA release in post-6 Psi-injured rats at 8 weeks.
Cognition and Motor Function Were Affected by Severe Head Injury in Contrast to Minor Injury
The fixed speed rotarod test data showed that impairment of this motor behavior only occurred in the 6 Psi-injured group [Fig. 5A, two-way ANOVA test, F(18, 238) = 1.91, followed by Bonferroni posttests, with p < 0.05 in sham vs. 6 Psi-injured groups, at 1 day, 1, 3, 4, 5, 6, 7, and 8 weeks postinjury], but there was no significant difference between sham and 2 Psi groups (squares) [Fig. 5A, two-way ANOVA test, F(18, 238) = 1.91, followed by Bonferroni posttests, with all p > 0.05 in sham vs. 2 Psi-injured group, at 1 day, and 1–8 weeks postinjury], which indicates that impairment in motor skill persists in the 6 Psi-injured group within our observation time.

Behavioral tests revealed impairment in cognition (NOR) and motor skills (fixed speed rotarod test) in injured animals. (A) In the fixed speed rotarod test, impairment only occurred initially (1 day after injury), and afterward there were no significant differences between the 2 Psi (squares) and sham group. However, there was persistent reduction in motor skills in the 6 Psi group (triangles). (B) NOR tests show impairment in cognition in injured animals by 1 week after injury in both 2 Psi and 6 Psi injury groups. Recovery of cognition was found in the 2 Psi group (squares), but not in the 6 Psi group (triangles). Analysis of the relationship between the maximum evoked DA levels with the behavioral impairments using linear regression. Significant relationships between DA levels and NOR impairment are shown for both mild (E; 2 Psi-injured group) and severe groups (F; 6 Psi-injured group). In the 6 Psi-injured group, both rotarod and NOR test performance was related to the DA levels significantly (D and F). In the 2 Psi-injured group, a significant relationship only existed in the NOR test (E), but not in the rotarod test (C). *p < 0.05, **p < 0.01, ***p < 0.001, #p < 0.05, #p < 0.001.
The cognitive function in rats was evaluated by using the NOR test. In the NOR test, the time that the animal spent exploring familiar and novel objects was recorded and converted to the percentage of the total exploration time. The DI is defined as exploration time of (novel minus familiar)/total time. A significant difference was found comparing the sham and 6 Psi-injured group [Fig. 5B, triangles, two-way ANOVA with F(10, 150) = 5.81 followed by Bonferroni posttest, with all p < 0.001 in sham vs. 6 Psi-injured group at 1, 2, 4, 6, and 8 weeks postinjury], but a significant difference was found only at 1 and 4 weeks postinjury comparing the sham and 2 Psi group [Fig. 5B, squares, two-way ANOVA with F(10, 150) = 5.81 followed by Bonferroni posttest, with p < 0.001 in sham vs. 2 Psi-injured group at 1 week postinjury, p < 0.05 at 4 weeks postinjury, and p > 0.05 at 2, 6, and 8 weeks postinjury]. This indicates that an impairment in cognition in injured animals is evident since the DI was very low at 1 week after injury in both 2 Psi and 6 Psi injury groups. Subsequently, DI deteriorated until 8 weeks postinjury in the 6 Psi group, but an increment in the index occurred in the 2 Psi group and was close to the basal value by 6 weeks.
We analyzed the relationship between maximum evoked DA levels and behavioral impairments by linear regression. When we correlated the behavioral data with FSCV data at each time point, significant relationships between DA dynamics and NOR impairment were found for both the mild (Fig. 5E; 2 Psi group) and severe group (Fig. 5F; 6 Psi group). Moreover, in the 6 Psi group, both the rotarod and NOR test performances were significantly related to the DA level (Fig. 5D, F). In the 2 Psi group, there was a significant relationship in the NOR test (Fig. 5E), but not in the rotarod test (Fig. 5C).
Striatal DA Synthesis System After TBI
To characterize the changes in DA synthesis and metabolism, we measured TH and monoamine oxidase B (MAO-B) protein expression at several time points after injury. Neither TH nor MAO-B was affected significantly in our TBI model (data not shown). Increased activity of TH has previously been associated with phosphorylation of TH at serine 40 (pser40TH), but not serine 19 (pser19TH) (30,41). Thus, we monitored pser40TH and pser19TH levels by Western blot at 2 h, 1 day, 1 week, 2 weeks, and 8 weeks after injury. For pser19TH levels, only at 2 h was there a statistically significant difference (Fig. 6A, B). For pser40TH levels, the 2 Psi-injured groups showed significant decrease at 6 weeks after injury. A reduction in pser40TH levels could be found at 2 h in 6 Psi-injured groups, but this reduction returned to normal by 24 h (Fig. 6C, D).

The phosphorylation of TH after brain injury. The phosphorylations of TH at serine 40 site (pser40TH) and serine 19 site (pser19TH) were analyzed by Western blot at 2 h, 1 day, 1 week, 2 weeks, and 8 weeks after injury. There was a statistically significant difference in the 6 Psi group at 2 h for pser19TH levels (B, **p < 0.01). For pser40TH levels in 2 Psi-injured groups there was a significant decrease at 6 weeks after injury (C, *p < 0.05). A significant depression in pser40TH level could be found at 2 h in the 6 Psi-injured group, and this returned to normal by 24 hours (D, *p < 0.05). The representative Western blots for pser19TH and pser40TH are shown in (E).
Discussion
Our data indicate that a change in dopamine release is seen after TBI, and the magnitude and duration of changes vary with injury severity (Fig. 3). The changes in DA release returned to near-normal ranges after 4 weeks in the mild (2 Psi) injury (Fig. 1), but decreased release persisted more than 8 weeks after severe (6 Psi) injury (Fig. 2). In our study, the greatest changes in the DA system after TBI were found between acute (1 day) and subacute stages (1 week after the injury). These changes include DA tonic and phasic release that were detected by FSCV, reuptake, and clearance shown by t values, as well as by changes in DA metabolism using HPLC.
TBI has been difficult to ascribe to a single disruption in neural circuitry that could explain such an array of clinical events. Indeed, persistent deficits experienced by TBI patients are probably due to a wide spectrum of different neural system dysfunctions. However, that does not preclude the possibility of utilizing a targeted therapeutic strategy to enhance cognitive recovery after TBI. DA is a particularly important system to study in this context because it is known to have a critical role in physiological events relevant to cognition, emotion, and in numerous brain systems affected by TBI (8,21).
In this study, we chose the striatum as a target to evaluate DA release and cognition/motor impairment after TBI. Dysfunctional nigrostriatal signaling has implications for cognitive functions, including memory, executive function, and attention, which has been epitomized by investigations in Parkinson's disease (1,38). Studies have also demonstrated that both the striatum and dorsolateral prefrontal cortex, another DA target, are important for executive function and working memory (9).
DA release input/output curves (Figs. 1 and 2) and maximum stimulation evoked DA signals (Fig. 3) revealed reductions were found at 1 to 2 weeks after injury, and recovery could be found in the 2 Psi group by 4 weeks after the injury. These data are in agreement with a previous study indicating that suppression of dopamine release in injured animals may recover (43). Furthermore, we showed these influences on the dopamine system after TBI results from changes in both tonic/phasic release and clearance/reuptake.
Dopamine Release Pattern and Animal Behavior After Fluid Percussion Injury
Activating DA cell activity could increase dopaminergic transmission, and this frequency-dependent release of DA has been termed the phasic DA response. In contrast, DA may also accumulate in the extracellular space in concentrations too low to stimulate postsynaptic receptors, but of sufficient magnitude to activate DA release-inhibiting autoreceptors. This steady-state level of extrasynaptic DA has been termed the tonic DA response (19).
DA neurons can operate in distinct temporal modes; they display responses across varying time scales, including fast phasic changes of DA release on a seconds time scale and slower phasic changes in a minute-to-hour range. Furthermore, tonic levels of DA provide a DA receptor “tone.” Temporally distinct responses of DA neurons in the prefrontal cortex and striatum have been related to different kinds of information processing and may underlie distinct behavioral functions (22).
The contrast between phasic and tonic DA signals is a key feature that varies within the striatum and may be endogenously and exogenously modulated by a number of factors. DA signaling may be therapeutically manipulated to aid different patient populations depending on their DA signaling needs (41,43,47). Furthermore, experimental work suggests that phasic DA release is central to reinforcement learning, whereas tonic DA release may modulate performance without altering learning per se (3). In our study, maximum evoked DA levels and behavioral change in the injured groups were analyzed by linear regression, and a significant relationship between DA levels and NOR impairment was shown in both the 2 Psi (Fig. 5E) and 6 Psi group (Fig. 5F). Moreover, in the 6 Psi group, both rotarod or NOR test performance deficits were related to the suppression of DA release (Fig. 5D, F). However, in the 2 Psi group, the relationship only occurred in the NOR test (Fig. 5E) and not in the rotarod test (Fig. 5C).
Factors Involving the FPI-Induced Suppression of Evoked Striatal DA Signal: Impairments in Reuptake, Metabolism, Releasing Probability, or Synthesis of DA
Effective neuronal communication depends on the rapid and efficient clearance of neurotransmitter from the synaptic cleft. The DA transporter is an important regulator of DA neurotransmission, and the striatum expresses the highest DA transporter levels in the central nervous system (33).
It is known that the striatum is vulnerable to damage from TBI. Even with the impact directed toward the hippocampal region, damage to the striatum can occur from experimental TBI. The known effects of experimental TBI on the rat striatum include axonal degeneration (11), neuronal cell loss (14), and ischemia (10). One of the most important systems affected by brain injury is the DA transporter within striatum. A significant decrease in DA transporter protein following controlled cortical impact at 7 days on the injured side and at 28 days bilaterally has been shown previously (44,46). The function of the DA transporter was altered in the striatum at 2 weeks and 4 weeks following FPI in the present study. In our study, we showed a very significant prolongation of t values, which indicates that the DA clearance rate is delayed by the injury. Although the clearance rate was prolonged significantly at subacute states (1 week after injury) in both 6 Psi and 2 Psi groups, the clearance rate improved by 4 weeks after injury in the 2 Psi group, but not in the 6 Psi group. In a rat model of cerebral concussion, DA utilization was decreased in the striatum, and DA levels were decreased in the whole brain at 6 h after injury (26). Regional changes in DA levels, measured by HPLC, were found after FPI, including a persistent decrease in the cortex. After controlled cortical impact in rats, TH, but not dopamine β-hydroxylase, showed a delayed increase (28 days) in the frontal cortex (45).
On the other hand, increasing reuptake was found here at 8 weeks postinjury, which may indicate that the regrowth or collateral sprouting of catecholaminergic axons may occur in the chronic stage in severely injured animals. These phenomena have also been demonstrated in experimentally induced lesions in adult CNS neurons (16,24). The occurrence of spontaneous regrowth of DAergic fibers after partial nigrostriatal denervation has been suggested, (34) and a long-term increase in the amount of striatal TH has been observed after 6-hydroxydopamine injection in the SN pars compacta (6,36). Although absolute striatal DA concentration was not affected by the injury in our study, our HPLC data showed increased turnover at 8 weeks after injury. Thus, further studies evaluating DA turnover, TH activity, DOPAC/DA, and DA in the nigrostriatal system after TBI are needed to confirm whether there is such a compensatory mechanism after TBI.
The Role of Dopamine in Behavioral Deficits in TBI Animals
To determine whether different levels of injury induced different deficits in behavior, we analyzed motor skills of rats by using the rotarod test, while cognition was evaluated by NOR in 2 Psi and 6 Psi groups. The rotarod tests rodent coordination, which involves the nigrostriatal dopaminergic pathway. Cognitive deficits can be categorized into one of three general domains: attention and processing speed, memory, and executive function (4,32). Of these, memory difficulties are the most commonly reported and most difficult for TBI patients and caregivers (5).
FPI affected cognition after injury in the NOR test, and the impairments recovered by 1 week after 2 Psi injury, but persisted up to 8 weeks after 6 Psi injury. In the rotarod test, the impairment persisted up to 8 weeks in 6 Psi-injured animals. In the 2 Psi-injured group, a deficit occurred initially in the acute stage (2 h and 1 day after the injury), but then recovered by 1 week.
Using linear regression analysis, the NOR and rotarod test data in 6 Psi-injured group were significantly related to DA release (Fig. 5D, F). The locomotor abnormalities correlated well with striatal DA release in the severely injured group, but not significantly in mildly injured animals (Fig. 5C, E). Given these observations, we postulate that these behavioral abnormalities after FPI are closely related to the dopaminergic system dysfunction.
We thus hypothesize that DA plays an important role in TBI-related cognition and motor deficits based on the following reasons: first, the role of DA in cognition and its functional anatomy relevant to TBI; second, observed changes in DA signaling and anatomy in experimental models, and the importance that these alterations have on cognitive and behavioral deficits. Furthermore, clinical research has demonstrated potential efficacy of dopaminergic medications in the treatment of TBI (17,28). Our data provide additional insights into potential mechanisms and therapeutic targets for chronic cognitive dysfunction. Pharmacotherapies targeting DA systems have beneficial effects after TBI, but it is not known which brain regions are most affected by the treatments or whether improvements in striatal function are responsible for any behavioral effects. In our study, although the striatum is relatively distal to the site of impact, it seems that the injury is diffuse enough that many brain regions are affected either by the initial impact or by secondary injury cascades. The results of this research and future studies should help in verifying the correlation between neuronal circuits and behavioral dysfunction and determining best course of treatment for patients in whom striatal function is disrupted by TBI. It should be noted that correlation does not prove causation. However, it does provide insights into the “null hypothesis”; if there were no correlation between DA synaptic dynamics and the behavioral changes after TBI, it would suggest that DA is not involved.
Conclusion
In this study, we showed that both tonic and phasic release of DA are suppressed by FPI, and the severity of the brain injury affects striatal DA release differentially. Then the behavioral deficits of animal are altered in different severity injured groups. In addition, these variations of DA releasing after brain injury were also found related to the behavior performance. Moreover, the recovery of DA release, turnover rate, and behavioral deficits could be found in 2 Psi FPI group in chronic stage, but the deficits in DA release and behavior persisted through our observation period in the 6 Psi FPI group.
Footnotes
Acknowledgments
This work was supported by the National Science Council of Taiwan under grant NSC 101-2314-B-016-008, USPHS grant NS070825, and by Medical Research Project grants TSGH-C101-084, and TSGH-C100-033 from the Tri-Service General Hospital of Taiwan. The authors declare no conflicts of interest.