
Abstract
Although neural stem cell (NSC) transplantation is widely expected to become a therapy for nervous system degenerative diseases and injuries, the low neuronal differentiation rate of NSCs transplanted into the inner ear is a major obstacle for the successful treatment of spiral ganglion neuron (SGN) degeneration. In this study, we validated whether the local microenvironment influences the neuronal differentiation of transplanted NSCs in the inner ear. Using a rat SGN degeneration model, we demonstrated that transplanted NSCs were more likely to differentiate into microtubule-associated protein 2 (MAP2)-positive neurons in SGN-degenerated cochleae than in control cochleae. Using real-time quantitative PCR and an immunofluorescence assay, we also proved that the expression of Wnt1 (a ligand of Wnt signaling) increases significantly in Schwann cells in the SGN-degenerated cochlea. We further verified that NSC cultures express receptors and signaling components for Wnts. Based on these expression patterns, we hypothesized that Schwann cell-derived Wnt1 and Wnt signaling might be involved in the regulation of the neuronal differentiation of transplanted NSCs. We verified our hypothesis in vitro using a coculture system. We transduced a lentiviral vector expressing Wnt1 into cochlear Schwann cell cultures and cocultured them with NSC cultures. The coculture with Wnt1-expressing Schwann cells resulted in a significant increase in the percentage of NSCs that differentiated into MAP2-positive neurons, whereas this differentiation-enhancing effect was prevented by Dkk1 (an inhibitor of the Wnt signaling pathway). These results suggested that Wnt1 derived from cochlear Schwann cells enhanced the neuronal differentiation of transplanted NSCs through Wnt signaling pathway activation. Alterations of the microenvironment deserve detailed investigation because they may help us to conceive effective strategies to overcome the barrier of the low differentiation rate of transplanted NSCs.
Introduction
Spiral ganglion neurons (SGNs) are primary auditory afferent neurons that deliver signals from hair cells in the cochlea, the peripheral auditory receptors, to the brain through the cochlear nerve. SGNs do not regenerate to any clinically significant extent after neural degeneration. However, the effect of cochlear implants, the main means to treat severe sensorineural hearing loss, largely depends on the degree of preservation of SGNs (17). Therefore, the repair and replacement of SGNs is one of the essential steps in any attempt to restore auditory function to this patient population.
In recent years, cell replacement therapy has emerged as a promising approach to replace damaged SGNs within the mammalian cochlea. Otologists tested this approach by transplanting stem cells into SGN-degenerated cochleae. Interestingly, data show that SGN-degenerated cochleae provide a more permissible microenvironment than normal cochleae for stem cells to survive (23,29), migrate into the injured tissue (36), and differentiate into neurons (14). However, the cellular and the molecular mechanisms by which the SGN-degenerated microenvironment regulates the behavior of neural stem cells (NSCs) remain unclear.
Schwann cells are glia cells of the peripheral nervous system (PNS). Schwann cells in the mature PNS can be further categorized into (i) myelinating, (ii) nonmyelinating, (iii) perisynaptic Schwann cells of the neuromuscular junction, and (iv) satellite cells that ensheathe the cell bodies of sensory neurons (1). Schwann cells in other peripheral nerves such as the sciatic nerve have been shown to support neurite growth and regeneration, a process that depends on secreted factors and molecules (2,8,19,44). In the cochlea of rodents, Schwann cells myelinate the peripheral axons, the cell bodies, and the initial portion of the central axons of SGNs (16,38,39). Cochlear Schwann cells are associated with growth and regrowth of spiral ganglion neurites (3,10,25,46,47), they may express neurotrophins (4,12), and they play a role in neuronal pathfinding for SGNs (18,31). Schwann cells that were genetically modified to express neurotrophins enhance SGN survival (34). Inactivation of fibroblast growth factor receptor signaling in Schwann cells has been shown to result in SGN degeneration (43). However, the effects of Schwann cells on the neuronal differentiation of transplanted NSCs in SGN-degenerated cochleae have not been well studied.
Wingless-type mouse mammary tumor virus (MMTV) integration site family proteins (Wnts) are secreted factors that regulate various cell fate decisions (6,40). Wnts signal through binding on a receptor complex, and they activate several intracellular signaling pathways including the canonical Wnt pathway (known as the Wnt/ β-catenin pathway) (5,48) and noncanonical pathways, including those mediated by Ca2+ or Jun N-terminal kinase (Jnk) (11,15,21). The Wnt signaling pathway is implicated in regulating the self-renewal capacity and the neuronal differentiation of NSCs during neural development (9,13,20,26,27). Hirabayashi et al. (13) compared previous studies and revealed the stage-specific roles of Wnt signaling during neural development: Wnt signaling activation increases the neuronal differentiation of NSCs in the late stage of neural development (28) but results in the expansion of NSCs and suppression of neuronal differentiation in the early stage of neural development (9,30,42). In adult mouse ear, Wnt receptors were found expressed in SGNs (37). While the functional significance of Wnt ligands and their receptors for neuronal differentiation during neural development is well established, the effects of these ligands and receptors during neural injury and repair in the adult ear are largely unclear.
The purpose of this study was to examine the effects of the microenvironment on transplanted NSCs. In this study, we transplanted NSCs into control or SGN-degenerated cochleae to show that the microenvironment in SGN-degenerated cochleae enhances the neuronal differentiation of transplanted NSCs. We also showed that the expression of Wnt1 increased in the satellite cells in SGN-degenerated cochlea. In vitro experiments demonstrated that Wnt1-expressing Schwann cells induced the neuronal differentiation of NSCs through Wnt signaling pathway activation. Therefore, we hypothesized that after SGN degeneration, the satellite cells in the spiral ganglion released Wnt1 and promoted the neuronal differentiation of transplanted NSCs by activating their intracellular Wnt signaling pathway.
Materials and Methods
Animals and Treatment Groups
Embryos of C57BL/6-green fluorescent protein (GFP) mice [gestational day (E) 14.5 (E14.5), n = 8] were used for initiating NSC cultures. Male and female newborn rats [postnatal day 1 (P1), n = 20] were used for initiating primary cochlear Schwann cell cultures. Adult female Sprague-Dawley (SD) rats (250–300 g) were used for cell transplantation experiments (n = 24) or animal model identification (n = 10).
The 24 SD rats used for cell transplantation experiments were divided equally into two groups. The animals in the first group were normal-hearing controls and received NSC transplantation into their inner ear. The second group first received local ouabain administration to cause SGN degeneration and then received NSC transplantation. In each animal, the left ear was the surgery ear, and the right ear served as an untreated control.
The 10 SD rats used for animal model identification received only the surgery to establish the SGN degeneration model; they did not receive NSC transplantation. In each animal, the left ear was treated with ouabain application to the round window niche (Fig. 1A), while the right ear served as a control and received the same surgical approach with injected physiological saline instead of ouabain.
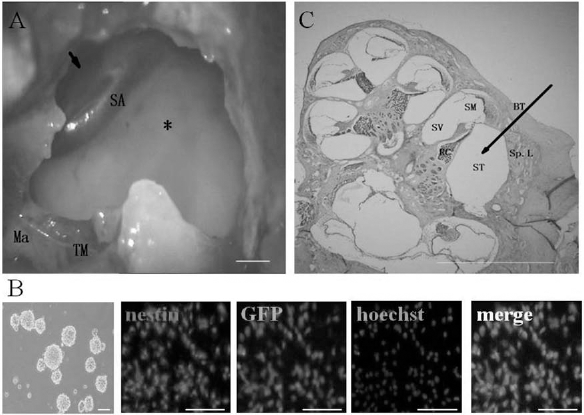
Approaches used for ouabain application and neural stem cell (NSC) transplantation (A, C) and the identification of cultured NSCs (B). (A) Surgical approach to expose the round window niche (arrow) for ouabain application and the basal turn of the cochlea (asterisk) for NSC transplantation. Scale bar: 500 μm. (B) The NSCs at passage 3 formed cell spheres. The cultured green fluorescent protein (GFP)-positive NSCs stained for nestin. Nuclei were counterstained with Hoechst 33342. Scale bar: 100 μm. (C) A radial frozen section of the cochlea from a normal adult rat. The arrow indicates the delivery routes of NSCs into the scala tympani (ST). TM, tympanic membrane; Ma, malleus; SA, stapedial artery; Sp.L, spiral ligament; S V, scala vestibule; SM, scala media; BT, basal turn of cochlea; RC, Rosenthal's canal. Scale bar: 500 μm.
The animals were raised in a low-noise environment [median sound level was 40 decibel (dB) sound pressure level (SPL)] and were kept on a normal day–night cycle (12/12) with free access to food and water. The SD rats used in the experiments were provided by the animal center of the Fourth Military Medical University (Xi'an, China). C57BL/6-GFP transgenic mice were gifted from the Institute of Neuroscience of the Fourth Military Medical University. The animal procedures reported in this study were approved by the Animal Care and Use Committee at the Fourth Military Medical University.
Establishment of the SGN Degeneration Animal Model
A brief application of ouabain to the round window niche destroys most cochlear SGNs while leaving hair cells intact (24,35). For surgical procedures, rats were anesthetized with 2% pentobarbital sodium (2 ml/kg, intraperitoneal; Northeast Pharm, Liaoning, China). Additional doses were given at half the starting dose when needed. Atropine (0.2 mg/kg, intramuscular; Northeast Pharm) was given to reduce secretions. Animals were placed on a homeothermic pad at 39°C. A posteroinferior skin incision was made in the retroauricular area of the left ear. The underlying muscles and the facial nerve were separated by blunt dissection to expose the middle compartment of the bulla. A 1-mm diamond paste burr (Bien-Air, Bienne, Switzerland) was used to make a small opening in the bulla, and the opening was expanded with hemostatic forceps to expose the cochlea and the round window niche (the arrow in Fig. 1A). A total volume of 10 μl of ouabain (10 mM, Sigma-Aldrich, St. Louis, MO, USA) was slowly administered to fill the round window niche. Ouabain was washed off after 60 min. The bulla was covered with the underlying muscle and fascia, and the incision was closed with a nonabsorbable suture (Jinhuan, Shanghai, China).
Cochlear Function Tests
To assess the hearing loss levels of the SGN-degenerated and the control ears, auditory brain stem response (ABR) audiometry (Bio-logic Systems, San Carlos, CA, USA) was performed. ABRs were measured at the three following times: before the ouabain application, 2 days after ouabain application, and 7 days after ouabain application. The rats were anesthetized, and needle electrodes were placed subcutaneously. The active electrode was inserted beneath the skin on the top of the head, the reference electrode was inserted beneath the pinna of the measured ear, and the ground electrode was inserted beneath the skin of the tail. Stimulus signals were generated through Intelligent Hearing Systems (Bio-logic Systems) and were delivered through an earphone. The stimulus signals were presented at a repetition rate of 10/s. Responses of 1,024 sweeps were averaged at each intensity level, which varied at intervals of 5 dB SPL. The threshold was determined by visual inspection of the evoked trace and defined as the lowest intensity level at which a clear waveform was visible.
Isolation and Culture of NSCs
NSCs were isolated and cultured by a previously described procedure (7,41). GFP-positive NSCs were collected from the embryonic olfactory bulbs of C57BL/6-GFP mice on E14.5 (the day on which a vaginal plug was found was considered E0.5). The timed-pregnant mother was anesthetized by injection of 2% pentobarbital sodium (2 ml/kg, intraperitoneal) and was sacrificed by cervical dislocation. The embryos were removed from their sacs. The embryonic brains were removed, and the olfactory bulbs were dissected. Cells were obtained by mechanical dissociation of dissected and pooled olfactory bulbs, followed by enzymatic digestion with 0.125% trypsin (Sigma-Aldrich) for 15 min at 37°C. After trypsin inhibition with heat-inactivated 10% fetal bovine serum (FBS; Hyclone, Logan, UT, USA), the cells were resuspended in Dulbecco's modified Eagle medium and Nutrient Mixture F-12 (DMEM/F12; HyClone) supplemented with B27 (Invitrogen, Carlsbad, CA, USA), epidermal growth factor (EGF; 20 ng/ml, PeproTech, Rocky Hill, NJ, USA), fibroblast growth factor-2 (FGF-2; 20 ng/ml, PeproTech), and antibiotics (100 IU/ml penicillin and 100 μg/ml streptomycin; Sigma-Aldrich) and were incubated at 37°C in a 5% CO2 atmosphere. Cells growing as floating spheres or aggregates were passaged every 4 to 7 days by mechanical procedures and plated at 5,000 cells/cm2 at each passage. NSCs were identified based on the expression of nestin (an NSC marker; rabbit anti-nestin; 1:200; Millipore-Chemicon, Billerica, MA, USA) (Fig. 1B). NSCs within passages 3–5 were used for further experiments.
NSC Transplantation
A second surgery to the inner ear was performed 2 days after ouabain application, and NSCs were transplanted into the scala tympani of the cochlea (Fig. 1A, C). Animals were anesthetized, an incision was made, and cochleae were exposed using the same protocol as used for the application of ouabain. A microdissecting needle (Yiman, Shenzhen, China) was used to drill a small hole (approximately 0.1 mm) in the bony wall of the basal turn of the cochlea (asterisk in Fig. 1A), allowing access to the scala tympani. A suspension of cultured NSCs (1 × 106 cells/ml) was then aspirated into a microliter syringe (Gaoge, Shanghai, China). The microliter syringe was coupled to a micropipe (homemade from a 200-μl Eppendorf pipette tip by pulling it after heating; external and internal diameters of tip: 100 and 40 μm, respectively). The tip of the micropipe was inserted into the hole in the bony wall of the basal turn of the cochlea. Then, 1 μl of NSC suspension was infused into the scala tympani over a period of 5 min (the arrow in Fig. 1C illustrates the delivery routes of NSCs into the scala tympani). When the micropipe was in place, the transplantation site was covered with a small piece of tissue to prevent spillage of the transplanted cells.
The bulla was covered with the underlying muscle and fascia, and the incision was closed with a nonabsorbable suture. The animal was placed on a homeothermic pad at 39°C for the recovery period. To prevent cell rejection, cyclosporine A (10 mg/kg, subcutaneous; Novartis, Basel, Switzerland) was given daily from the day of transplantation to the end of the experiment.
Immunofluorescence Staining
For immunohistochemistry, animals were deeply anesthetized and perfused with 100 ml of phosphate-buffered saline (PBS; O'BioLab, Beijing, China) followed by 300 ml of 4% paraformaldehyde (Fuyu, Tianjin, China). Both cochleae were removed and immersed in the same fixative at 4°C overnight. Samples were then decalcified in 10% ethylenediaminetetraacetic acid (EDTA; Amresco, Solon, OH, USA) for 10–14 days and cryoprotected in 30% sucrose (Fuyu) for 24 h. Serial 10-μm frozen sections of each cochlea embedded in OCT compound (Electron Microscopy Sciences, Hatfield, PA, USA) were prepared. The sections were mounted on glass slides (Fuyu), dried in room temperature air for 30 min, and washed in PBS three times. Some of these histological sections were stained with hematoxylin and eosin (H&E; Beyotime, Jiangsu, China) according to standard procedures. The remaining sections were permeabilized with 0.3% Triton X-100 (Sigma-Aldrich) in PBS for 30 min, blocked with 1% normal goat serum (Boster, Wuhan, China) for 60 min at 37°C, and then incubated at 4°C overnight with the following primary antibodies: mouse anti-neuron-specific enolase (NSE; 1:200; Boster), rabbit anti-microtubule-associated protein 2 (MAP2; 1:1000; Millipore-Chemicon), rabbit anti-glial fibrillary acidic protein (GFAP; 1:500; Millipore-Chemicon), mouse anti-Wnt1 (1:200; Abcam, Cambridge, UK), rabbit anti-S100β (1:500; Epitomics, Burlingame, CA, USA), and rabbit antinestin (1:200; Millipore-Chemicon). The sections were washed with PBS and then incubated with the appropriate secondary antibodies. For nuclear staining, the slides were counterstained in Hoechst 33342 (1:1000, Sigma-Aldrich) at room temperature for 15 min. The sections were rinsed, dehydrated, and coverslipped for microscopic evaluation under a fluorescence microscope system with appropriate filters (Olympus, Tokyo, Japan). Photographs were taken with a digital camera (Olympus).
RT-PCR and Real-Time RT-PCR
For quantitative real-time RT-PCR, animals were decapitated under deep anesthesia 2 days after ouabain application. The cochleae were immediately removed. The bony capsule was removed, and the spiral ligament, the stria vascularis, and the basilar membrane were separated from the central modiolus under a dissecting microscope. The soft tissue was carefully dissected out of the osseous modiolus. Total RNA was isolated from the spiral ganglion samples collected from ouabain-treated and control rats with Trizol reagent (Invitrogen) following the manufacturer's protocol, and cDNA was synthesized. Real-time PCR was performed with a kit (SYBR Premix EX Taq, Takara, Kyoto, Japan) and the ABI PRISM 7300 real-time PCR system. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) served as the reference control gene.
For RT-PCR, total RNA of NSC cultures was isolated using the same protocol. PCR products were separated on a 5% polyacrylamide gel (BioDev-Tech, Beijing, China) and detected by autoradiography. Sizes were confirmed with a DL2000 DNA ladder marker (Takara, Kyoto, Japan). Primers are summarized in Table 1.
Primer Sequences for RT-PCR and Real-Time RT-PCR

Wnt1, Wingless-type mouse mammary tumor virus (MMTV) integration site family protein 1; fzd3, frizzled family receptor 3; Ctnnb1, catenin (cadher-in-associated protein), b1, 88 kDa; Dvl1, dishevelled segment polarity protein 1; Lef1, lymphoid enhancer-binding factor 1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Isolation and Purification of Cochlear Schwann Cells
Cochlear Schwann cells were isolated and purified by a previously described procedure (45). Briefly, newborn rats (P1) were used. Dissociated modiolus tissue was plated in a six-well plastic plate (Corning, Corning, NY, USA), previously coated with poly-l-lysine (0.1 mg/ml, Sigma-Aldrich), and was cultured in DMEM/F12 complete medium (containing 10% FBS and antibiotics) for 4 days with changes of medium every other day.
Cells were purified using an immunomagnetic purification procedure. A microcolumn designed to handle up to 1 × 107 magnetically labeled cells from a total of 2 × 108 total cells was used (Miltenyi Biotec, Gladbach, Germany). The procedure followed the instructions provided by the manufacturer of the microbeads (Miltenyi Biotec). Rabbit anti-75-kDa low-affinity neurotrophin receptor (P75; Millipore-Chemicon) and MACS micro-beads coupled to goat-anti-rabbit IgG antibody (Miltenyi Biotec) was used in this procedure. After immunomagnetic purification, the cells were counted, added to a six-well dish previously coated with poly-l-lysine, and cultured in DMEM/F12 complete medium (containing 10% FBS and antibiotics). Cells were maintained at 37°C in 5% CO2/air with changes in medium every other day.
Lentiviral Vector Construction
Rat Wnt1 DNA was cloned into a lentiviral expression vector (Tronolab, Lausanne, Switzerland), and the final vector with red fluorescent protein (RFP), SKF-RFP-Wnt1 was confirmed by sequencing. The target gene was transfected into human embryonic kidney 293T (HEK293T) cells (Cell Bank of the Chinese Academy of Sciences, Shanghai, China) together with a lentiviral packaging mix (Tronolab, Lausanne, Switzerland) using Lipofectamine 2000 (Invitrogen). After 72 h, the virus was harvested in the supernatant. The viral supernatant was concentrated by centrifugation at 4,000 × g for 10 min at 4°C, filtered through a 0.45-μm filter (Corning), and stored at −80°C.
Lentiviral Vector Infection
For infection experiments, cochlear Schwann cells were seeded in a 24-well plate (Corning) at 50,000 cells/ well. The next day, Schwann cells (50,000 cells) were incubated for 24 h with 5 × 106 viral particles in DMEM/ F12 complete medium (containing 5% FBS). After 24 h of incubation, the cells were washed, and the medium was replaced with DMEM/F12 complete medium (containing 10% FBS). The expression of RFP was detected 96 h after infection.
Wnt1 Western Blotting
At 96 h after infection, protein extracts were prepared from cochlear Schwann cells infected with lenti-virus encoding either RFP alone (control) or both RFP and Wnt1. After removal of the medium and washing with PBS, lysis buffer was added. Each protein sample was electrophoresed onto 12% sodium dodecyl sulfate (SDS)–polyacrylamide gels (Amersham Biosciences, Piscataway, NJ, USA) and transferred onto nitrocellulose membranes (Amersham Biosciences). The membranes were blocked with 5% fat-free milk powder (Yinqiao, Xi'an, China) for 2 h at room temperature and incubated with mouse anti-Wnt1 (1:500; Abcam) at 4°C overnight. Then, the membranes were incubated with a secondary antibody conjugated to horseradish peroxidase (Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 2 h at room temperature. β-Actin (Abcam) was used as an internal control. The reaction product was visualized with enhanced chemiluminescence (Amersham Biosciences).
Transwell Coculture
GFP-positive NSCs were cocultured for 4 days with cochlear Schwann cells infected with a lentivirus encoding either RFP alone (control) or both RFP and Wnt1. NSCs (5 × 104/well) were cultured in 24-well plates covered with poly-l-lysine-treated coverslips and were cultured in DMEM/F12 containing 10% FBS supplemented with antibiotics. The Schwann cells (5 × 105/well) were plated in transwell chambers (polyester membranes with 0.4-μm pores; Corning), and the chambers were placed in the wells above the NSCs. The transwell allows exchange of the media; however, cell-to-cell contact between NSCs and Schwann cells is inhibited by the polyester membranes. After 4 days of coculture, neuronal differentiation was evaluated by immunofluorescence to quantify the percentage of NSCs expressing the mature neuronal marker MAP2.
For experiments involving the antagonism of the Wnt pathway, recombinant Dickkopf 1 (Dkk1, a canonical Wnt pathway inhibitor; 100 ng/ml; R&D Systems, Minneapolis, MN USA) was added to the media of the transwell coculture system.
Statistical Analysis
Repeated-measures ANOVA analysis (with Bonferroni confidence interval adjustment) and LSD t tests were used for ABR threshold data. The difference between the two groups was analyzed using Student's t test. The difference among three or more groups was analyzed with one-way ANOVA followed by the least significant difference test. Values of p < 0.05 were considered statistically significant. Data are presented as the mean ± SD. All statistical analyses were performed using SPSS version 16.0 software (SPSS Inc., Chicago, IL, USA).
Results
Identification of an SGN Degeneration Animal Model
To establish an SGN degeneration animal model, we locally administered 10 mM ouabain solution into the round window niche of rats. Two days after ouabain treatment, animals were sacrificed, and immunohistochemistry staining was used to identify the pathology of the SGNs (Fig. 2). H&E-stained sections showed an empty space in the area normally occupied by the SGNs: only few SGNs survived (Fig. 2A, A′). A similar result was obtained by immunofluorescence staining, in which the SGN cell bodies and dendrites were stained with NSE (a neuron-specific marker) (Fig. 2B, B′). Two days after ouabain treatment, most of the NSE-positive SGNs disappeared; only 5% of them survived in the ouabain-treated group (Fig. 2B) compared with the control group (Fig. 2B′), which received the same surgical approach but was injected with physiological saline instead of ouabain. These results confirmed that the local application of ouabain destroyed most of the SGNs in the cochlea.

Ouabain-induced spiral ganglion neuropathy. (A–E) Spiral ganglion sections from ouabain-treated ears. (A′–E′) Spiral ganglion sections from control ears. (A, A′) Sections were stained with hematoxylin and eosin (H&E). (B)–(E) and (B′)–(E′) were stained for immunofluorescence. (B, B′) Neuron-specific enolase (NSE; a neuron-specific marker) was used to mark SGNs. (C, C′) S100 (a Schwann cell marker) was used to mark satellite cells. (D, D′) Nuclei were counterstained with Hoechst 33342. Spiral ganglia neurons (SGNs) were plentiful in the Rosenthal's canal in the control ear (arrow in B′) but were almost completely absent in the treated ear; few SGNs survived (arrow in B). (C′, E′) In normal spiral ganglion, Schwann cells enclosed the cell bodies of SGNs. (C, E) In ouabaintreated spiral ganglion, the shape of Schwann cells markedly changed. Most of them lost their ensheathing structure. In the places where few neurons survived, Schwann cells still enclosed the cell bodies of the neurons (arrow in C, E). Scale bar: 100 μm. (F) ABR thresholds were significantly elevated 2 days and 7 days after ouabain treatment, consistent with a massive loss of SGNs. *p < 0.05 (repeated-measures ANOVA analyses with Bonferroni confidence interval adjustment). dB SPL, decibel sound pressure level.
To confirm the neuronal effects of the ouabain treatment, hearing loss levels were assessed by measuring ABR (Fig. 2F). Ouabain-treated ears experienced a significant elevation in hearing threshold, from 22.50 ± 5.24 dB SPL prior to ouabain application to 88.33 ± 2.58 dB SPL 2 days after ouabain application (p < 0.05) and 87.50 ± 4.18 dB SPL 7 days after ouabain application (p < 0.05). The observed auditory threshold elevations were consistent with a massive loss of SGNs. In contrast, control ears, which received the same surgical approach but were injected with physiological saline instead of ouabain, did not show a significant change in hearing threshold after a 7-day recovery, indicating there was no significant influence of surgery on hearing function.
In summary, we established an SGN degeneration animal model using local ouabain application.
The Shape of Satellite Cells Changes After SGN Degeneration
Satellite cells are one subcategory of Schwann cells, which ensheathe the cell bodies of sensory neurons in the PNS. In the spiral ganglion, each neuronal cell body is completely surrounded by several satellite cells to form a discrete anatomical unit. The interdependence between the neuron and the satellite cell underpins the functioning of the entire PNS, and the fates of these two cell types are inextricably entwined (1).
To observe the changes in satellite cells after SGN degeneration, we stained satellite cells with S100β (a Schwann cell marker) and examined the cells by immunofluorescence (Fig. 2C, C′). In the normal spiral ganglion, satellite cells enclose the cell bodies of SGNs, as shown in the control group (Fig. 2C′, E′). However, in the ouabain-treated group, the shape of the satellite cells markedly changed (Fig. 2C, E). Most of them no longer enclosed the cell bodies of SGNs; instead, there were only a few surviving neurons, and satellite cells still enclosed the cell bodies of the neurons (arrow in Fig. 2C, E). These results indicated that because of the cutoff of the intimate physical contact with the neurons, the shape of the satellite cells changed significantly.
SGN Degeneration Enhances the Neuronal Differentiation of Transplanted NSCs
To test whether SGN degeneration promotes the neuronal differentiation of transplanted NSCs, we transplanted GFP-positive NSCs into control or SGN-degenerated cochleae 2 days after ouabain application. Cochleae were fixed 7 days after NSC transplantation and subjected to immunostaining to examine the neuronal differentiation of the transplanted NSCs (Fig. 3). GFP-positive cells were found at several positions in the cochleae, including the scala tympani and the scala vestibule of the basal turn, the middle turn, and the apex turn. We only observed and quantified the neuronal differentiation of the transplanted NSCs located within or close to the Rosenthal's canal (where the cell bodies of SGNs are located). In control cochleae, all of the GFP-positive cells were found outside the Rosenthal's canal (Fig. 3A, C), and very few (~1%) of these cells were positive for MAP2 (Fig. 3C, G). By contrast, in SGN-degenerated cochleae, a few GFP-positive NSCs migrated into the Rosenthal's canal (arrow in Fig. 3D, F), and more (~4%) transplanted NSCs differentiated into MAP2-positive cells than in the controls (Fig. 3F, G).

SGN degeneration promotes the neuronal differentiation of transplanted NSCs. GFP-positive NSCs were transplanted into control or SGN-degenerated cochleae 2 days after ouabain application. Cochleae were fixed 7 days after NSC transplantation and were subjected to immunostaining. Microtubule-associated protein 2 (MAP2; a mature neuron marker) and GFP were used to mark neurons and transplanted NSCs, respectively. (A–C) Spiral ganglion sections from control ears. (D–F) Spiral ganglion sections from ouabain-treated ears. Dotted lines indicate the boundary of the Rosenthal's canal. (A, C) In control cochleae, all transplanted GFP-positive NSCs were located outside of the Rosenthal's canal (where the cell bodies of SGNs were located). In contrast, in SGN-degenerated cochleae, a few GFP-positive NSCs migrated into the Rosenthal's canal (arrows in D, F). More GFP-positive NSCs expressed MAP2 in SGN-degenerated cochleae (F) compared with those in control cochlea (C). Arrow head in (F) indicates a surviving SGN. The H&E-stained normal cochlea image beside indicates the localization and orientation of these sections. Scale bar: 100 μm. (G) There was a fourfold increase in the proportion of cells that colocalized MAP2 and GFP in the ouabain-treated cochlea as compared with the controls. Only the GFP-positive NSCs located within or close to the Rosenthal's canal of the basal turn were observed, whereas those located in other parts of the cochlea were excluded. Data are the mean ± SD of values from 12 samples. *p < 0.05 versus control.
This result suggests that after the degeneration of endogenous neurons, certain factors in the microenvironment of the spiral ganglion may change and affect the neuronal differentiation of transplanted NSCs. We then tried to identify these factors.
SGN Degeneration Modulates Wnt1 Expression in Cochleae
Wnt proteins regulate the neuronal differentiation of NSCs and play an essential role during neural development (9,13,26,27). To verify whether Wnt proteins also participate in the repair and regeneration of neurons after adult SGN degeneration, we investigated the difference in Wnt1 expression between control and SGN-degenerated cochleae by quantitative real-time RT-PCR. As shown in Figure 4A and B, the expression of Wnt1 (a ligand of the canonical Wnt pathway) was markedly upregulated in SGN-degenerated cochleae compared with control cochleae, suggesting that SGN degeneration enhances the expression of Wnt1 in the spiral ganglion microenvironment.

Wnt1 mRNA increased in the spiral ganglion after ouabain application. (A, B) Total RNA was prepared from spiral ganglion samples collected from SGN-degenerated or control adult rat cochleae. The expression of wingless-type mouse mammary tumor virus (MMTV) integration site family protein 1 (Wnt1) was measured by RT-PCR (A) and real-time PCR (B), with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as the reference control. (C) RT-PCR analysis of canonical Wnt signaling pathway components in NSCs. Frizzled family receptor 3 (Fzd3; 120 bp); catenin (cadherin-associated protein), β1, 88 kDa (Ctnnb1; 137 bp); axin1 (137 bp); dishevelled segment polarity protein 1 (Dvl1; 125 bp); lymphoid enhancer-binding factor 1 (Lef1; 131 bp); M: DL2000 marker.
Wnt1 protein is a secreted glycoprotein, and it can bind to cells that express a Wnt1 receptor in an autocrine and/ or paracrine fashion (32,33). We used RT-PCR analysis to reveal that NSC cultures express receptors for Wnt and crucial canonical Wnt signaling pathway components (Fig. 4C), suggesting that NSCs have the ability to bind to Wnt ligands and activate the intracellular Wnt signaling pathway.
Based on these results, we hypothesized that after SGN degeneration, the microenvironment may enhance the neuronal differentiation of transplanted NSCs through Wnt1 signaling.
Cochlear Satellite Cells as the Source of Region-Specific Signals
To examine the source of the increased Wnt1 protein, we observed the region where Wnt1 protein is expressed in control and SGN-degenerated cochleae (Fig. 5). Using an immunofluorescence assay, we found that in control cochleae, Wnt1 is only expressed in the organ of Corti rather than in the spiral ganglion. In contrast, in SGN-degenerated cochleae, not only the organ of Corti but also the spiral ganglion expresses Wnt1. Double staining of S100 (a Schwann cell marker) and Wnt1 showed that in SGN-degenerated cochlea, most of the Wnt1-positive cells were S100-positive satellite cells (Fig. 6). Meanwhile, the few surviving SGNs were also Wnt1 positive (arrow in Fig. 6). The surviving SGNs were identified by their spherical shape. These results indicated that the increased Wnt1 protein was mainly derived from the satellite cells in the SGN-degenerated cochlea.

Wnt1 expression increased in the spiral ganglion after ouabain application. In control cochleae, Wnt1 was expressed only in the organ of Corti but not in the spiral ganglion. In SGN-degenerated cochleae, Wnt1 was expressed not only in the organ of Corti but also in the spiral ganglion. Dotted lines indicate the boundary of the Rosenthal's canal. The arrows show the position of the organ of Corti. Scale bars: 100 μm.
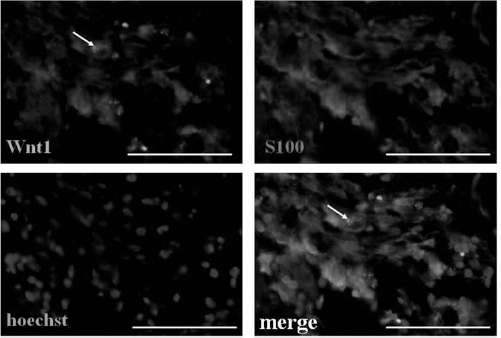
The increased Wnt1 expression was mainly derived from satellite cells in the spiral ganglion. Spiral ganglion sections from ouabain-treated ears were stained for immunofluorescence analysis. S100 (a satellite cell marker) was used to mark satellite cells. The arrow shows a surviving SGN. Scale bars: 50 μm.
Based on these results, we hypothesized that after SGN degeneration, cochlear satellite cells may enhance the neuronal differentiation of transplanted NSCs through Wnt1 signaling.
Wnt1-Expressing Schwann Cells Promote the Neuronal Differentiation of NSC Cultures
We used an in vitro coculture system of NSCs and Wnt1-expressing Schwann cells that models the interaction of these cell populations in the SGN-degenerated cochlea.
Cochlear Schwann cells were isolated, purified, and identified with S100β (Fig. 7A). Forty-eight hours after purification, cultures were ~86% S100-positive Schwann cells. Cochlear Schwann cell cultures were infected with a lentivirus encoding either RFP alone (control) or both RFP and Wnt1 (Fig. 7B), and Western blotting was used to determine Wnt1 protein production in these cells (Fig. 7C). A marked increase in Wnt1 protein expression was observed in the cells infected with the lentivirus encoding both RFP and Wnt1 (Fig. 7C), indicating that after infection, Wnt1 production in cochlear Schwann cells increased.

Identification and infection of cochlear Schwann cells. (A) Purified cochlear Schwann cell cultures are identified with S100. Scale bar: 100 μm. (B) Cochlear Schwann cell cultures infected with lentivirus encoding both RFP and Wnt1. Arrows show the uninfected Schwann cells, and arrowheads show the infected Schwann cells. Scale bars: 100 μm. (C) Lentivirus infection increases Wnt1 protein production in cochlear Schwann cells. Cochlear Schwann cells were infected with a lentivirus encoding either RFP alone (control) or both RFP and Wnt1. Western blotting was used to determine the production of Wnt1 protein. β-Actin was used as an internal control. Western blots shown are representative of three different experiments with similar results.
The cochlear Schwann cells infected with the lentivirus encoding either RFP alone (control) or both RFP and Wnt1 together were subsequently transwell cocultured for 4 days with GFP-positive NSCs in the presence or absence of a Wnt inhibitor, Dkk1. Neuronal differentiation was evaluated on the 5th day by quantifying the percentage of GFP-positive NSCs expressing the mature neuronal marker MAP2 (Fig. 8). More than 27% of the NSCs cocultured with Wnt1-expressing Schwann cells were MAP2 positive after 4 days of coculture, compared with only 14% of those cocultured with control Schwann cells (Fig. 8). In the presence of Dkk1, the percentage of MAP2-positive cells only slightly increased (~16%) compared with control, suggesting that the Wnt signaling in NSCs, which enhances the neuronal fate commitment of NSCs, was suppressed by Dkk1. These results suggested that Wnt1-expressing Schwann cells activate the Wnt signaling pathway in NSCs. By contrast, the percentage of GFAP-positive glial cells was not significantly increased (date not shown), which indicated that Wnt1-expressing Schwann cells specifically enhanced the generation of neurons.

Wnt1-expressing Schwann cells promote neuronal differentiation of NSC cultures. Cochlear Schwann cell cultures were infected with a lentivirus encoding either RFP alone (control) or both RFP and Wnt1 (coculture). The cells were then transwell cocultured with GFP-positive NSCs for 4 days in the presence or absence of a Wnt inhibitor, Dkk1. (A) After 4 days of coculture, more GFP-positive NSCs cocultured with Wnt1-expressing Schwann cells expressed MAP2, compared with those cocultured with control Schwann cells. Nuclei were counterstained with Hoechst 33342. Scale bars: 50 μm. (B) There was an approximately twofold increase in the proportion of cells that colocalized MAP2 and GFP in the coculture group as compared with the controls. This increase can be repressed by Dkk1. *p < 0.05.
Discussion
Previous findings indicated that SGN-degenerated cochleae provide a more permissible environment than normal cochleae for stem cells to survive (23,29), migrate into the injured tissue (36), and differentiate into neurons (14). Hu et al. (14) showed that the neuronal differentiation ratio of transplanted NSCs was discouragingly low in normal cochleae. Interestingly, after transplantation into a chemically deafened inner ear, NSCs showed increased neuronal differentiation (14). These results suggested a link between microenvironment alterations and modulation of transplanted stem cells. Our results were consistent with these findings. In the first part of this study, using an SGN degeneration rat model, we demonstrated that a greater amount of transplanted NSCs differentiated into MAP2-positive neurons in SGN-degenerated cochleae compared with control cochleae (Fig. 3), suggesting that SGN-degenerated cochleae release unknown factors that enhance the neuronal differentiation of transplanted NSCs.
Next, we attempted to clarify the molecular mechanisms contributing to the link between the microenvironment and transplanted stem cells. Theoretically, factors in the microenvironment regulate the maintenance, proliferation, and neuronal fate commitment of transplanted NSCs. In light of the clarified link between the Wnt signaling pathway and the neuronal lineage commitment of NSCs during neural development (9,13,26,27), we sought to determine whether there is a similar link during the repair and replacement of degenerated SGNs. Thus, we used quantitative real-time RT-PCR to assess the expression of Wnt1 (a ligand implicated in canonical Wnt signaling) in control and SGN-degenerated cochleae. The expression of Wnt1 increased significantly in the SGN-degenerated cochleae compared with the expression in the control cochleae (Fig. 4A, B). According to these results, we hypothesized that the increased Wnt1 expression provides a molecular background for the neuronal differentiation of transplanted NSCs.
Previous reports showed that Schwann cells revert to nonmyelinating phenotypes in the deafened rat cochlea (16). In the ouabain-induced SGN-degeneration model, hyperplasia and hypertrophy of the satellite cells in spiral ganglion were observed (22). In this study, we observed that the shape of the satellite cells in SGN-degenerated spiral ganglion changed compared with their shape in the control ganglion (Fig. 2C, C′). In addition, satellite cells in the control ganglion did not express Wnt1, whereas those in SGN-degenerated ganglion expressed Wnt1 (Fig. 5, 6). Based on these changes, we hypothesized that the increased Wnt1 protein was derived from the satellite cells in the SGN-degenerated cochlea.
Furthermore, using RT-PCR, we showed that NSC cultures from the olfactory bulb express receptors and signaling components for Wnt proteins (Fig. 4C). In brief, we found that Schwann cells express Wnt1 and NSCs express receptors for Wnts and crucial canonical Wnt signaling pathway components. Based on these expression patterns, we hypothesized that Schwann cell-derived Wnt1 and Wnt signaling might be involved in the regulation of neuronal differentiation of the transplanted NSCs.
In the second part of this study, we tested our hypothesis in vitro in a coculture system of NSCs and Wnt1-expressing Schwann cells that models the interaction of these cell populations in the SGN-degenerated cochlea. For in vitro studies, we transduced a lentiviral vector expressing Wnt1 into cochlear Schwann cell cultures and cocultured them with NSCs from the olfactory bulb. Coculture with Wnt1-expressing Schwann cells resulted in a significant increase in the percentage of NSCs that differentiated into neurons, whereas the percentage only slightly increased in the presence of Dkk1 (a canonical Wnt pathway inhibitor) (Fig. 8), indicating that the Wnt1 signal from Schwann cells promotes the neuronal differentiation of NSCs.
In conclusion, we emphasize two aspects of these findings: (i) Wnt1 appears to function as an extracellular cue that promotes neuronal differentiation of transplanted NSCs; (ii) Schwann cells take charge of the microenvironment alteration, and they synthesize and secrete the canonical Wnt signaling ligand Wnt1. Our data indicate that Schwann cells are a regulator of the SGN-degenerated microenvironment and have a role in the repair of degenerated neurons. Wnt1 secreted from the local Schwann cell population participates in regulating the neuronal differentiation of transplanted NSCs. However, in SGN-degenerated cochleae, the neuronal differentiation rate is still very low, and it was estimated that only ~4% of the transplanted cells actually differentiated into neurons within the cochleae (Fig. 3), which indicates that the self-modulation of SGN-degenerated cochleae is not adequate to sufficiently enhance the neuronal commitment of NSCs. To reach the final functional replacement and repair through NSC therapy, further studies are needed to explore various effective pretransplantation manual interventions to sufficiently enhance the neuronal differentiation of NSCs, such as cotransplantation of NSCs with Wnt1-expressing Schwann cells. According to the significance of the microenvironment revealed by this study, it may also be beneficial to change the microenvironment into a more permissible one for neuronal commitment of NSCs.
Footnotes
Acknowledgments
Lentiviral Vector Construction was performed by BiAng Biotech, Shanghai, China. This study was supported in part by grants from the National Natural Scientific Foundation of China (81120108008, 30930098, 81100713, 81170911) and the National Key Basic Research Development Plan (No. 2011CB504505). We thank Dr. J. Chen for his excellent technical assistance. The authors declare no conflict of interest.