
Abstract
Liver failure after liver resection for cirrhosis is a critical problem, and no effective therapy except liver transplantation is currently available. The objective of this study was to examine whether hepatocyte transplantation (HT) reduces the poststandard liver resection mortality rate of rats with nonalcoholic steatohepatitis (NASH)-related cirrhosis. Liver resection for hepatocellular carcinoma (HCC) combined with NASH-related cirrhosis has become increasingly common. We developed a rat model of acute liver failure after two-thirds partial hepatectomy (PH) for NASH-related cirrhosis. The mechanism by which HT improved the survival of the model rats was examined in short- and long-term investigations. Female DPPIV- recipient F344 rats were fed the choline-deficient l-amino acid (CDAA)-defined diet for 12 weeks. Some of the rats were transplanted with male F344 DPPIV+ rat hepatocytes 24 h before undergoing PH. The overall post-PH survival of each group was evaluated, and short- and long-term pathological and molecular biological evaluations were also performed. Overall survival was significantly longer in the HT group than the non-HT group (7-day survival rates: 46.7% and 7.7%, respectively). Compared with the recipient livers of the non-HT group, numerous Ki-67+ hepatocytes and few TUNEL+ hepatocytes were observed in the livers of the HT group. At 6 months after the HT, the DPPIV+ hepatocytes had partially replaced the recipient liver and formed hepatocyte clusters in the spleen. Preoperative HT might improve the survival of rats with NASH-related cirrhosis after PH by preventing the host hepatocytes from accelerating their growth and falling into apoptosis.
Keywords
Introduction
Hepatocellular carcinoma (HCC) is the third leading cause of cancer-related death in the world, and its mortality rate has recently been increasing (41). Most cases of HCC in Eastern countries are associated with hepatitis B viral infection, whereas the majority of cases in Western countries involve hepatitis C viral infection (43). In addition, the incidence of nonalcoholic steatohepatitis (NASH)-associated HCC is also increasing all over the world (3,9,14). The annual cumulative incidence of HCC has been reported to range from 0.05% to 2.6% in patients with NASH-related cirrhosis (3,6), and surgical treatment for NASH-related HCC has become more common.
Liver transplantation is the best treatment for HCC, but its prognosis is poor in patients that do not meet the Milan criteria (34). On the other hand, liver resection and liver transplantation produce comparable clinical outcomes in patients who maintain good liver function and meet the Milan criteria (10,40). Therefore, liver resection is still considered to be an effective therapy for HCC in patients with NASH. Poor liver function is the main obstacle to liver resection, as massive hepatectomy in patients with poor liver function often leads to postoperative liver failure and death (1). Therefore, improving the liver function of patients with poor liver function would improve the short- and long-term clinical outcomes of patients with HCC derived from NASH-related cirrhosis.
The acceleration of remnant liver regeneration and function, the amelioration of liver fibrosis, and the protection of liver cells against various chemical mediators would help to avoid liver failure after hepatectomy (4). If a treatment that would restore these functions could be administered preoperatively to patients with advanced cirrhosis, postoperative liver failure might be avoidable. Hepatocyte transplantation (HT) has been performed in patients suffering from acute liver failure or chronic liver disease, including those with congenital metabolic conditions, to support their liver function and provide a bridge to liver transplantation (12,15,36,47). In addition, HT has been suggested to have potential as a therapeutic strategy for supporting liver function after massive hepatectomy (13). Although HT has been shown to improve the liver function of decompensated cirrhosis patients (2,24), the exact clinical benefits of HT for patients with NASH-related cirrhosis who undergo massive hepatectomy are largely unknown. Furthermore, the fate of the transplanted cells is poorly understood.
The consumption of the choline-deficient l-amino acid (CDAA)-defined diet induces NASH-related fibrosis and cirrhosis in rats (37,46). In the present experiment, we extended the feeding term of the CDAA-defined diet to examine whether long-term consumption of the diet leads to severe liver fibrosis and a high mortality rate after massive hepatectomy. As a result, we found that most rats that consumed the CDAA-defined diet for 12 weeks died after two-thirds partial hepatectomy (PH), and hence, we considered that these rats could be used as a model of NASH-related cirrhosis leading to postoperative liver failure. The aims of this study were 1) to examine the ability of HT to prevent high mortality after PH in rats with NASH-related cirrhosis, 2) to elucidate the mechanism by which HT supports liver functions in this liver failure rat model, and 3) to determine the fate of the transplanted hepatocytes in the recipient liver via long-term observation.
Materials and Methods
Animals and Liver Injury Model
Male F344 rats [dipeptidyl peptidase IV (DPPIV)+ strain; Sankyo Lab Service Corporation, Inc., Tokyo, Japan] were used as donors, and female F344 rats (DPPIV- strain; Charles River, Wilmington, MA, USA) were used as recipients (45). All rats were maintained at a constant temperature of 23 ± 1°C under a 12-h light-dark cycle and fed standard chow and water ad libitum. After acclimatizing to the conditions, the rats were continuously fed the CDAA-defined diet (Dyets, Inc., Bethlehem, PA, USA). We then performed an experiment to determine the survival rate of rats with severe hepatic fibrosis after PH. In this experiment, 5-week-old female rats were fed the CDAA-defined diet for 4, 8, 10, or 12 weeks, and PH was performed on the last day of treatment. PH was carried out under inhalation-induced anesthesia, which was produced using a mixture of O2/N2 (1:1) and sevoflurane (Maruishi Pharmaceutical Co., Osaka, Japan) (18). After the operation, the rats were fed standard chow. All animals received humane care, and the experimental protocol was approved by the Committee of Laboratory Animals according to the guidelines defined by Sapporo Medical University.
Chemicals
Unless otherwise noted, all chemicals from Sigma-Aldrich Co. (St. Louis, MO, USA).
Isolation and Transplantation of Hepatocytes
Crude hepatocytes were isolated from the liver of a 7-week-old male rat using the two-step collagenase perfusion method (8). The cells were then purified to obtain mature hepatocytes (MHs) using Percoll (GE Healthcare Biosciences, Pittsburgh, PA, USA) gradient centrifugation (28). The viability of the cells was determined to be greater than 90% by the trypan blue (Dojindo, Kumamoto, Japan) exclusion test. The cells were finally resuspended in saline at a concentration of 2 × 107 viable cells/ml for transplantation. Twenty-four hours before the PH, intrasplenic HT was performed. Female rats that had consumed the CDAA-defined diet for 12 weeks were anesthetized using a mixture of O2/N2 (1:1) and sevoflurane. The lower pole of the spleen was exposed via a left flank incision. Then, 0.5 ml of saline with or without MHs (1 × 107 cells) was injected into the splenic pulp. The puncture wound produced by the injection was closed with 4-0 silk sutures (Ethicon, Inc., North Ryde, NJ, USA) to promote hemostasis.
Histological Analysis
The rats were sacrificed at 3, 6, 12, 24, 72, or 168 h after the PH. Then, their livers were removed and immediately cut into 5-mm-thick slices. Some slices were frozen, and others were fixed in 4% paraformaldehyde (PFA; Merck KGaA, Darmstadt, Germany)/phosphate-buffered saline (PBS). The frozen sections were used for the enzyme-based histochemical and immunohistochemical examinations. For all sections, nonspecific peroxidase activity was blocked by immersing the sections in 0.6% H2O2 solution for 30 min. The sections to be frozen were embedded in Tissue-Tek (Sakura Finetechnical Co., Tokyo, Japan), frozen using isopentane/dry ice, and kept at −80°C until use. The slices subjected to 4% PFA fixation were embedded in paraffin and used for hematoxylin-eosin (H&E), Oil red O, Azan-Mallory, or terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick-end labeling (TUNEL) staining. The H&E and Azan-Mallory staining were performed according to standard protocols. The frozen sections were fixed in 4% PFA, stained with Oil Red O (Muto Pure Chemicals Co., Ltd., Tokyo, Japan) for 15 min, and then counterstained with hematoxylin.
Immunohistochemical and Enzyme-Based Histochemical Examinations of DPPIV Expression
To identify the donor cells, an enzyme-based histochemical examination of DPPIV expression was carried out. DPPIV enzyme activity was detected as described previously (45). For the fluorescent immunohistochemical examinations, frozen liver sections were fixed in absolute cold ethanol after being washed with PBS. After nonspecific reactions had been blocked using 10% BlockAce (DS Pharma Biomedical, Osaka, Japan), the sections were incubated with the primary antibodies for 1 h at room temperature, washed with PBS, and then fluorescent-conjugated secondary antibodies (Alexa Fluor Dye; Life Technologies Corp., Grand Island, NY, USA) were applied for 30 min. Double staining was carried out using the former enzyme-based histochemical method (45) to detect DPPIV [rabbit anti-cluster of differentiation 26 (CD26) antibody; 1:50; Santa Cruz Biotechnology, Inc., Dallas, TX, USA] and the latter immunohistochemical method (above) to detect Ki-67. Double immunostaining of Ki-67 (1:100; Santa Cruz)/albumin (1:100; DakoCytomation, Glostrup, Denmark) and Ki-67/cytokeratin 8 (CK8; 1:20; PROGEN Biotechnik GmbH, Heidelberg, Germany) was also performed. Finally, the samples were embedded in 90% glycerol containing 0.01% p-phenylenediamine and 4,6-diamidino-2-phenylindole (DAPI).
Polymerase Chain Reaction (PCR)
For DNA analysis, genomic DNA from the liver and spleen were extracted using a GenElute Mammalian Genomic DNA kit (Sigma-Aldrich). PCR for the rat sex-determining region Y (SRY) gene was performed using the following SRY primers: sense, 5′-CAGAGATCAGC AAGCATCTGG-3′; antisense, 5′-TCTGGTTCTTGGA GGACTGG-3′, for 28 cycles of 94°C for 30 s, 57°C for 30 s, and 72°C for 30 s. Rat glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as a control (sense, 5′-ACCACAGTCCATGCCATCAC-3′; antisense, 5′-TC CACCACCCTGTTGCTGTA-3′) (Clontech Laboratories, Inc., Palo Alto, CA, USA) at 94°C for 30 s, 57°C for 30 s, and 72°C for 1 min with 26 cycles. TaKaRa Taq PCR kits and a TaKaRa thermal cycler (Takara Bio Inc., Shiga, Japan) were used for all PCR reactions.
Digital Image Analysis of Liver Fibrotic Area
NIH Image V1.62 (National Institutes of Health, Bethesda, MD, USA) was used to evaluate liver fibrosis. Tissue sections underwent Azan–Mallory staining, which stains the fibrotic area blue. Adobe Photoshop CS (Adobe Systems Incorporated, San Jose, CA, USA) was used to ensure that this was depicted as the only blue signal. Fifteen different fields in three different specimens were examined for each sample with one field containing at least 10 portal tracts. The area of blue staining was thus determined (23).
Western Blotting
Liver tissue was cut into small pieces (20–50 mg), dissolved in lysis solution [20 mM Tris–HCl (pH 7.4), 150 mM NaCl, 2.5 mM ethylenediaminetetraacetic acid (EDTA), 1% Triton X-100, 1% aprotinin, and 20 mg/ml leupeptin], and homogenized using a Dounce homogenizer (NIPPON Genetics, Tokyo, Japan). The protein concentrations of the samples were then normalized using a bicinchoninic acid (BCA) assay kit (Pierce, Rockford, IL, USA) and NanoDrop 2000 (Thermo Scientific, Waltham, MA, USA). The proteins (20 μg/lane) in the samples were separated by 4–12% Bis-Tris gel (Invitrogen, Carlsbad, CA, USA) electrophoresis, before being transferred electro-phoretically to nitrocellulose membranes (GE Healthcare Biosciences) using an iBlot apparatus (Invitrogen). The membranes were then probed with primary antibodies diluted in primary antibody diluent (Invitrogen Western Breeze kit), before a chemiluminescence detection method (Pierce) was performed using Kodak MR film. Mouse anti-proliferating cell nuclear antigen (PCNA; 1:250; DakoCytomation), rabbit anti-nuclear factor k-light-chain-enhancer of activated B-cells (NF-kB; 1:250; Abcam, Cambridge, UK), rabbit anticleaved caspase 3 (1:250; Cell Signaling Technology, Inc., Danvers, MA, USA), goat anti-annexin V (1:250; Santa Cruz), and goat anti-GAPDH (1:250; Santa Cruz) antibodies were used as primary antibodies.
Coomassie Blue Staining
The gel after being blotted to a membrane was incubated in 0.1% Coomassie blue dye (Sigma-Aldrich) in 40% ethanol and 10% acetic acid for 1 h at RT. The gel was washed with 10% ethanol and 7.5% acetic acid for several days until the signal bands were clear to see.
TdT-Mediated dUTP Nick-End Labeling (TUNEL) Staining
To detect apoptotic cells, paraffin-embedded sections were stained with the TUNEL technique using an In Situ Apoptosis Detection Kit (Takara Shuzo, Ohtsu, Japan), according to the manufacturer's instructions. For semiquantitative analysis, the numbers of TUNEL-positive cells were counted in 30 randomly selected fields (magnification: 400×). The mean percentages of TUNEL-positive cells in the HT and control groups were calculated at 3, 6, 12, and 24 h after PH.
Multiple Cytokine Detection
Serum obtained after euthanasia was kept at −80°C until submission to a company (Upstate United States, Inc., Charlottesville, VA, USA) for analysis. Briefly, multianalyte profiling was performed on a Luminex 100 system and the XY Platform (Luminex Corporation, Austin, TX, USA). Calibration microspheres for classification and reporter readings, as well as sheath fluid, from the Luminex Corporation. Acquired fluorescence data were analyzed using MASTERPLEXTM QT (Ver. 1.2; MiraiBio, Inc., South San Francisco, CA, USA). Serum concentrations of tumor necrosis factor (TNF)-α and interleukin (IL)-6 were measured with an Upstate Beadlyte Mouse Multicytokine Bead master kit (Upstate United States, Inc.). All analyses were performed according to the industrial protocols.
Statistical Analysis
The Student's t test was used to compare the group means as appropriate. Overall survival rates were calculated using the Kaplan–Meier method, and comparisons were performed using the log-rank test. All calculations were performed using the StatView 5.0 software package (Abacus Concepts Inc., Berkeley, CA, USA) or the software SPSS 16.0 (SPSS Inc., Chicago, IL, USA). All results are expressed as the mean ± standard deviation (SD). Values of p < 0.05 were considered to be statistically significant.
Results
Induction of Liver Fibrosis and the Survival Rate After Two-Thirds PH
To determine the severity of the liver injuries that caused the death of most rats after PH, rats were fed the CDAA-defined diet for 4, 6, 8, 10, or 12 weeks. On the last day of treatment, PH was performed, and then the rats were observed for 1 week. All of the rats that continued to eat the CDAA-defined diet after the PH died within a few days, regardless of the duration of their CDAA-defined diet treatment, and so their diets were changed to standard chow after the operation. Although all of the rats that consumed the diet for 4 or 6 weeks survived the PH, some of the rats that consumed the diet for 8, 10, or 12 weeks died, and their survival rates at 7 days were 67% (6/9), 44% (4/9), and 11% (1/9), respectively (Fig. 1A). Therefore, a 12-week CDAA-defined diet consumption period was chosen for the HT experiment. Figure 1B (a, d, g) shows intraoperative macroscopic views of the liver taken during the PH. In the rats that consumed the CDAA-defined diet for 8 weeks, the surface of the liver was irregular, and some small nodules were found on each lobe, whereas the rats that consumed the CDAA-defined diet for 10 weeks displayed yellowish livers consisting of various numbers of nodules with diameters of 2–3 mm. The nodules in the livers of the rats that consumed the CDAA-defined diet for 12 weeks were larger, and some of the largest nodules were redder than the others. Histological analyses using Azan (Fig. 1B, b, e, h) and Oil red O (Fig. 1B, c, f, i) staining revealed that consuming the CDAA-defined diet for 8 weeks induced bridging fibrosis and fat deposition in most hepatocytes. In addition, a large part of the cell cytoplasm was occupied by lipid droplets. The livers of the rats that had consumed the CDAA-defined diet for 10 weeks consisted of small nodules that were almost lobule sized. The nodules were separated by thick collagen fibers, and most hepatocytes stored fat in their cytoplasm, which was indicative of severe steatosis. The livers of the rats that had consumed the CDAA-defined diet for 12 weeks possessed irregularly sized nodules. In some of these nodules, most of the hepatocytes contained lipid droplets. Compared to the livers of the rats that had consumed the diet for 10 weeks, the size and number of lipid droplets were decreased, and the fibrotic area was increased in the livers of the rats that had consumed the diet for 12 weeks. As shown in Figure 1C, the degree of hepatic fibrosis increased with the period of CDAA-defined diet consumption. At 10 weeks or more after the treatment, more than 20% of the liver was occupied by connective tissue including portal triads. In addition, serum TNF-α and IL-6 levels in the rats gradually increased with increasing feeding time on the CDAA diet (data not shown). High levels of serum TNF-α were observed in rats that did not undergo HT, but the variation in the levels between animals meant that no significant differences were observed (data not shown).

Survival rates of rats and the results of histological examinations of the liver after the consumption of the CDAA-defined diet. The rats were fed the choline-deficient l-amino acid (CDAA)-defined diet for 4, 6, 8, 10, or 12 weeks. The survival rates of the rats fed the CDAA-defined diet for 8 weeks (circles), 10 weeks (triangles), or 12 weeks (squares) after two-thirds partial hepatectomy (PH), which were determined by Kaplan–Meier analysis, are shown in (A). Intraoperative macroscopic views of the liver (a, d, g) obtained during PH, and the results of Azan–Mallory (b, e, h) and Oil red staining (c, f, i) are shown (B). Scale bar: 100 mm. The fibrotic area was calculated by subjecting Azan–Mallory-stained samples to image analysis (purple areas) using ImageJ (C).
Post-PH Survival of Rats That Underwent Hepatocyte Transplantation
Twenty-four hours before the PH, freshly isolated MHs (1 × 107 viable cells) were transplanted into the livers of some of the rats that had been administered the CDAA-defined diet for 12 weeks. As shown in Figure 2, within 24 h, 5 of the 13 (38.5%) rats that did not undergo HT (control) had died, whereas only 1 of the 15 (6.7%) rats that underwent HT had died. At 7 days after the operation, 8 of the 15 rats that had undergone HT were alive, whereas 12 of the 13 control rats were dead. On the other hand, when hepatocytes were transplanted during the PH, all of the rats died within 7 days (11 of 11 rats died; data not shown). Therefore, the administration of fresh HT prior to PH significantly improved the post-PH survival rate of the cirrhotic rats (log-rank test; p = 0.031).

Overall survival rates after two-thirds partial hepatectomy (PH) in the rats fed the CDAA-defined diet for 12 weeks with or without hepatocyte tansplantation. Solid line (-): transplanted group (N = 15); dotted line (▪▪▪): nontransplanted group (N =13). Log-rank test: p = 0.031.
To clarify the reason why HT increased the post-PH survival rate of the cirrhotic rats, the fate of the transplanted cells was examined. As shown in Figure 3A, at 24 h after the HT, many donor cells remained in the spleen. Hepatocyte aggregates were observed in the red pulp of the spleen, whereas only a few DPPIV+ cells were found in the liver. The number of DPPIV+ cells in the spleen decreased with time after HT, and 9.5%, 5.8%, 4.6%, and 4.3% of the spleen contained DPPIV+ cells at 1, 3, 5, and 7 postoperative days, respectively (Fig. 3B). On the other hand, at day 7, small clusters of DPPIV+ hepatocytes were observed in the liver (Fig. 3A, d), but only 0.4% of the liver was occupied by DPPIV+ cells (Fig. 3B). An examination of SRY gene expression in the spleen and liver also suggested that the number of transplanted cells in the liver gradually increased and that many donor cells remained in the spleen (Fig. 3C). At day 7, SRY gene expression was detected in about 7% and 15% of the transplanted cells in the liver and spleen, respectively. Double immunohistochemical staining of DPPIV and Ki-67 revealed that most of the DPPIV+ cells in the liver were Ki-67 positive (Fig. 3D, a), indicating that the donor cells gradually proliferated in the liver. On the other hand, immunohistochemical staining of albumin revealed that the transplanted hepatocytes in the spleen expressed albumin even at 7 days after transplantation (Fig. 3D, b).

Exploration of transplanted cell fate. Enzyme-based histochemical examination of dipeptidyl peptidase IV (DPPIV) expression in rats with HT with hematoxylin background staining (A). Livers (a–d) and spleens (e–h) at days 1 (a, e), 3 (b, f), 5 (c, g), and 7 (d, h) after PH. The DPPIV-positive area was calculated by image analysis of each specimen (B). PCR analysis of sex-determining region Y (SRY) gene expression in the liver and spleen after PH was performed (C,a), and the relative expression level [to glyceraldehyde 3-phosphate dehydrogenase (GAPDH)] of the SRY gene was measured in the liver and spleen after PH using densitometric analysis (C,b). Double immunohistochemical staining of Ki-67 (green) and DPPIV (red) was carried out in the liver (D,a) at day 7 after HT. Double immunohistochemical staining of albumin (red) and 4′,6-diamidino-2-phenylindole (DAPI; blue) was performed in the spleen (D,b) at day 7 after HT. Scale bar: 200 μm. The columns and bars in (B) and (C) represent the mean ± SD, respectively.
Responses of Resident Cells in the Liver
Next, we examined whether the resident hepatocytes were able to respond to growth stimuli after PH. In the livers subjected to HT, many Ki-67+ cells were detected, especially at 24 h after the PH (Fig. 4A, c and d). Double immunohistochemical staining of Ki-67 and CK8 indicated that most of the Ki-67+ cells in the livers subjected to HT were positive for CK8 (Fig. 4A, e and f), while most of the Ki-67+ cells in the control livers were located in fibrotic lesions (Fig. 4A, a and b). The number of Ki-67+CK8+ cells in the liver was measured at 6 and 24 h after the PH (Fig. 4B). The livers subjected to HT contained 18 ± 3.2 and 26 ± 5.2 Ki-67+ hepatocytes at 6 and 24 h, respectively. In addition, the frequency of Ki-67+CK8+ cells was higher in the livers of the rats subjected to HT than in those of the rats that did not undergo HT. These results were confirmed by examination of the expression levels of growth-related proteins. As shown in Figure 4C, the protein expression levels of PCNA and NF-kB were markedly increased in the livers subjected to HT at both 6 and 24 h posttransplantation compared with those observed in the livers that were not subjected to HT.
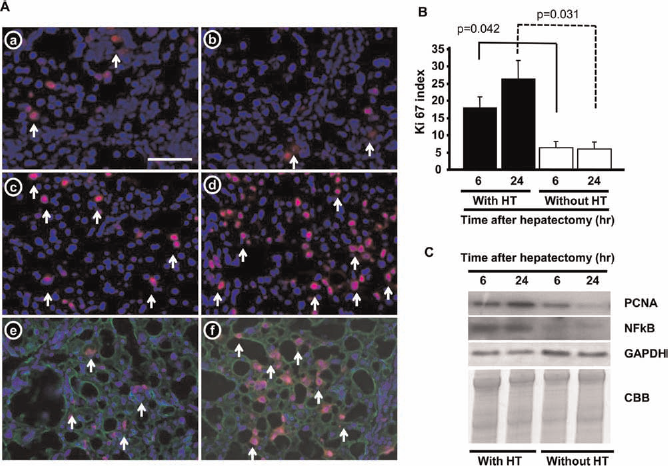
Fluorescent immunohistochemical staining of Ki-67 (red) and CK8 (green) after PH in the livers of rats fed the CDAA-defined diet for 12 weeks. (A) Two-thirds PH was performed in rats that were not (a, b) or were subjected to HT (c–f). Fluorescent images obtained at 6 (a, c, e) and 24 h (b, d, f) after PH are shown. To detect Ki-67+ hepatocytes, rat liver specimens that had been obtained at 6 (e) or 24 h (f) after PH were subjected to triple immunostaining of Ki-67 (red stain), cytokeratin 8 (CK8; green stain), and DAPI (blue stain). Arrows show Ki-67+ nuclei. (B) The Ki-67+ index is shown as a percentage (the number of Ki-67+CK8+ cells divided by the total number of CK8+ cells in the target area × 100). Student's t test was used for comparing with HT and without HT groups. (C) Western blot analysis of proliferating cell nuclear antigen (PCNA) and nuclear factor k-light-chain-enhancer of activated B cells (NF-kB) expression in the transplanted and nontransplanted livers after PH, using GAPDH as loading control. CBB, Coomassie blue staining. Scale bar: 100 μm. The columns and bars in (B) represent the mean ± SD.
Next, we investigated whether performing HT prior to PH prevented the resident hepatocytes from undergoing cell death. Tissue specimens from the livers of rats that did or did not undergo HT at 6 and >24 h after PH and subjected to TUNEL staining. As shown in Figure 5A, many large cells with positive nuclei and clear cytoplasm that displayed the morphological characteristics of hepatocytes were observed in the livers that were not subjected to HT (Fig. 5A, c and d), whereas in the livers subjected to HT, a few cells, which were located in or near fibrotic septa, possessed TUNEL+ nuclei (Fig. 5A, e and f). TUNEL+ cells were found to be more than four times more common in the livers that were not subjected to HT than in those that were (Fig. 5B). Although the number of TUNEL+ cells decreased with time after PH in both groups, it was significantly higher in the livers that were not subjected to HT at both time points. Furthermore, the protein expression levels of both cleaved caspase 3 and annexin V, which are markers of apoptosis, were markedly increased in the livers that were not subjected to HT at 6 h after the operation (Fig. 5C). These results were concordant with the TUNEL staining results.

TUNEL staining was performed in the livers of rats fed the CDAA-defined diet for 12 weeks. (A) The livers of the rats fed the CDAA-defined diet for 12 weeks (a) or normal chow (b) without PH or HT were used as controls. Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) staining at 6 (c and e) and 24 h (d and f) after PH in the livers of rats that were not (c and d) or were subjected to HT (e and f). Arrows show TUNEL-positive nuclei. (B) The TUNEL index is shown as a percentage (the number of TUNEL-positive cells divided by the total number of cells in the target area × 100). Student's t test was used for comparing with HT and without HT group. (C) Liver specimens obtained from rats that did or did not undergo HT after PH were subjected to Western blot analysis of cleaved caspase 3 and annexin V expression, with GAPDH as loading control. CBB, Coomassie blue staining. Scale bar: 50 mm. The columns and bars in (B) represent the mean ± SD.
Repopulation of the Recipient Liver by the Transplanted Hepatocytes in the Surviving Rats
The fate of the donor cells was investigated in the rats that survived for more than 1 month. At 6 months after the PH, although the areas of fibrosis in the liver were clearly smaller than those observed at 7 days after the PH, the liver consisted of large nodules surrounded by thin sections of connective tissue. Hepatic steatosis was improved at 6 postoperative months compared with that observed at 7 days after the PH, and the morphology of the hepatocytes had normalized. An enzyme-based histochemical examination of DPPIV expression in the liver revealed that many DPPIV+ hepatocytes had formed large foci and that large areas of the recipient liver had been replaced by donor hepatocytes (Fig. 6A and B). On the other hand, in the spleen, DPPIV+ cells formed large clusters that consisted of many hepatic cord-like structures (Fig. 6C and D). The DPPIV+ cells expressed albumin (data not shown). Although all of the rats displayed many donor cells in their spleens during the early period after HT, at 6 postoperative months, the size and number of DPPIV+ cell clusters in the spleen varied among the rats, and the area of donor cells was smaller than that in the liver (data not shown). Some DPPIV+ cells in both the liver (Fig. 6E–G) and spleen (data not shown) expressed Ki-67 protein in their nuclei. These results indicate that the transplanted hepatocytes had gradually proliferated in both organs in the absence of definitive growth stimuli. In addition, some rats that underwent HT were able to survive for more than 2 years, and no tumors were observed in their livers; however, their livers consisted of large nodules surrounded by thin sections of connective tissue, which displayed cirrhotic histology.

Enzyme-based histochemical analysis of DPPIV expression in the recipient rat liver (A and B) and spleen (C and D) at 6 months after HT. Double staining of Ki-67 (fluorescent immunohistochemistry) and DPPIV (enzyme histochemistry) was performed in the hepatocyte-transplanted livers at 6 months after PH; Ki-67 (E), DPPIV (F), and a merged image (G). Scale bar: 1.0 mm (A, C), 100 μm (B, E). Image (D) was taken at the same magnification as (B).
Discussion
The clinical importance of NASH was first realized in 1980 when NASH was distinguished from alcoholic liver disease (31). Various NASH models have been studied to elucidate the mechanism responsible for the disease and establish suitable therapies (46). The CDAA-defined diet and methionine/choline-deficient diet (MCDD) are often used to induce NASH-like disease in rats (37). MCDD model rats develop steatohepatitis as well as severe body weight loss and hypersensitivity to insulin (29,42,46). Although CDAA-defined diet model rats histologically display steatohepatitis in their livers, similar to those fed the MCDD, no obvious phenotypes have been reported (46). In addition, the CDAA-defined diet causes liver cancer at a much higher incidence than MCDD (37). Therefore, we used the CDAA-defined diet to develop a postoperative liver failure model that is similar to the condition experienced by humans, even though the pathophysiological mechanism responsible for the liver failure suffered by the model might be different from that seen in humans.
Clinically, patients with steatohepatitis display high rates of morbidity and mortality after hepatectomy (5,26). To increase the utility of hepatectomy for steatohepatitis patients who develop liver cancer, it is very important to understand the pathophysiology of the disease. However, it has proved very difficult to establish suitable animal models of NASH (38). Vetelainen et al. (48) reported that liver regeneration after PH was impaired in rats with severe steatosis induced by the consumption of an MCDD for 5 weeks. One possible reason why our model successfully reproduced the clinical situation experienced by human NASH-related cirrhosis patients was the duration of the feeding period. Our results suggest that steatosis severity before PH plays an important role in determining the postoperative outcome in this patient group. Therefore, we employed long-term CDAA-defined diet feeding to reproduce the clinical situation of NASH-related cirrhosis patients in the present experiment. In fact, the post-PH survival rates of the rats worsened as the duration of the CDAA-defined diet consumption period increased beyond 8 weeks, and most of the rats that consumed the diet for 12 weeks died within a week. The liver's ability to regenerate is lost once its spare hepatic function falls below a certain threshold (16). Thus, the livers of the rats that consumed the CDAA-defined diet for 12 weeks probably lost the ability to regenerate and maintain homeostasis after PH, resulting in hepatic failure.
In the present experiment, if HT was not performed before the PH, or if the MHs were transplanted during the operation, more than 66% of the rats died within 2 days of PH, indicating that the preconditioning of the liver is an important factor in the postmassive hepatectomy survival of rats with NASH-related cirrhosis. It has been reported that in the healthy rat liver, most transplanted hepatocytes are destroyed within 24–48 h (17,22). Most transplanted hepatocytes fail to engraft due to their entrapment in portal vein radicles, and it seems that these cells are primarily cleared by macrophage-mediated processes. In addition, in rats treated with retrorsine followed by PH, it was found that most intrasplenically transplanted hepatocytes did not remain within the spleen (25). In the present experiment, many transplanted hepatocytes remained in the spleen (about 15% of the donor cells at day 7), which resulted in the maintenance of hepatic functions, such as albumin production, for more than a week, whereas a small number of the donor cells infiltrated the liver. Although we did not measure blood pressure, portal hypertension caused by cirrhosis might retard the translocation of hepatocytes from the spleen to the liver. Therefore, the fact that the transplantation of a relatively small number of hepatocytes (1 × 107 cells) was able to save about half of the rats with NASH-related cirrhosis suggests that the maintenance of the newly supplied healthy hepatocytes in the spleen prepared the rats for the partial loss of their hepatic function and, hence, prevented them from falling into hepatic failure and provided time for the recipient hepatocytes to regenerate.
In liver regeneration, hepatocyte activation requires the cells to be “primed” by inflammatory cytokines, such as IL-6 and TNF-α (16,19). These cytokines are released by Kupffer cells in response to factors carried by the hepatic portal system, such as lipopolysaccharides. On the other hand, it is known that in normal rodents, liver injury after PH is mediated by TNF-α (20,51) and that death receptor ligands such as TNF-α and Fas ligand are involved in the induction of apoptosis and in triggering the destruction of the liver (32). In a previous study, the administration of exogenous TNF-α in combination with other toxins induced fulminant hepatic failure and hepatocyte apoptosis (30). In addition, increased preoperative TNF-α and IL-6 levels were observed in steatotic rats, which were indicative of chronic Kupffer cell activation (35,48,49). TNF-α facilitates apoptosis by activating caspases (7), and the ligand binding of TNF-α to its receptor also leads to the activation of the NF-kB pathway. In the present study, after PH, many hepatocytes in the livers of the rats with NASH-related cirrhosis fell into apoptosis, as shown by TUNEL staining and the increased protein expression of cleaved caspase 3 and annexin V, whereas a relatively large number of apoptotic hepatocytes was observed in the livers of the rats that underwent HT. On the other hand, a group of cytokine-triggered pathways that deliver survival signals to hepatocytes has been demonstrated to exist (33). Under certain conditions, the actions of these survival pathways can counteract the death signal. In fact, liver failure might represent the deregulation of hepatic homeostasis as a result of an imbalance between damaging and protective signals that are very tightly regulated under physiological conditions (32). IL-6 activates the transcription factor NF-kB (27), which plays an important role in increasing the survival and decreasing the apoptosis of hepatic cells (21,39). Impaired NF-kB activation is associated with defective proliferation and increased apoptosis of hepatocytes in chronic liver disease (11,44,50). In this study, the protein expression levels of NF-kB and PCNA were markedly increased, and many hepatocytes in the livers subjected to HT before PH were Ki-67+. These results suggest that HT prevents resident hepatocytes from dying and provides energy for their survival. Therefore, we consider that increased serum cytokine levels, especially increased IL-6 and TNF-α levels, will be observed in rats that are, and are not, subjected to HT, respectively. After the operation, we measured the concentrations of TNF-α and IL-6 in the sera of the surviving rats. Although the rats that did not undergo HT tended to display high TNF-α levels, the TNF-α levels of each rat varied greatly, and no significant difference was observed between the TNF-α levels of the rats that did and did not undergo HT (data not shown). It is known that Kupffer cell activation is also caused by cell transplantation (17,22), so HT might also affect the serum TNF-α levels. Further studies should be performed to clarify the pathophysiology of rats with NASH-related cirrhosis after massive hepatectomy.
Hepatocyte transplantation not only prevented liver failure in rats with NASH-related cirrhosis after PH but also replaced the injured hepatocytes with healthy ones. At 1 week after the transplant, although only about 0.4% of the area of the liver contained donor cell-derived hepatocytes, the repopulated areas gradually expanded (Fig. 6), and many DPPIV+ hepatocytes were observed at more than 1 year after the transplant. In addition, part of the spleen had also been replaced with donor hepatocytes, which gradually proliferated and reconstructed liver-like structures. In the present study, the CDAA-defined diet was replaced with a normal one after the PH, and the model rats' steatosis of the liver rapidly improved, although thin sections of fibrotic tissue persisted even at 2 postoperative years. The fact that the donor hepatocytes were able to proliferate, and survived for a long period, might indicate that a chronic growth stimulus is generated by the hepatocyte loss caused by steatosis-induced injury. Although cirrhosis has been considered to be irreversible for a long time, recent therapies have been demonstrated to decrease fibrosis in cirrhotic livers and improve the patient's condition. However, the liver regeneration of patients with NASH-related cirrhosis might be impaired by their damaged hepatocytes. Thus, in such patients, preoperative HT might decrease the risk of hepatic failure and gradually replace damaged hepatocytes with healthy ones.
Footnotes
Acknowledgments
We thank Ms. Minako Kuwano, Yumiko Tsukamoto, and Chieko Futamata for their technical assistance. We also thank Mr. Kim Barrymore for his help in preparing this manuscript. This study was supported by a grant from the Ministry of Education, Culture, Sports, Science, and Technology, Japan; a Grant-in-Aid for Young Scientists (B) for Y. Nakamura (No. 23791489); a Grant-in-Aid for Scientific Research (C) for T. Mizuguchi (No. 23591993); Grants-in-Aid for Scientific Research (B) for K. Hirata (No. 22390259) and T. Mitaka (No. 21390365); and a grant from the Yuasa Memorial Foundation for T. Mizuguchi. The authors declare no conflicts of interest.