Having participants perform 50 maximal isokinetic contractions, as introduced by Thorstensson and Karlsson (1976), is to this day one of the most commonly used fatigue protocols. Purpose: To determine to what extent a Thorstensson fatiguing protocol affects isokinetic and isometric performance characteristics.
METHODS:
Twenty-five college-aged men performed an isokinetic fatigue protocol consisting of 50 maximal elbow flexions at a moderate speed (180s). Pre- and post-tests were used to calculate fatigue indices (FI%) for the following variables: isometric and isokinetic peak torque (PT), isometric rate of torque development (RTD), and maximum acceleration (ACC).
RESULTS:
Each of the four dependent variables significantly decreased with fatigue (each demonstrated 0.001), but not of the same magnitude. Isometric PT FI (18.7%) was significantly less than isokinetic PT FI (45.1%, 0.001) and ACC FI (26.3%) was significantly less than isometric RTD FI (54.9%, 0.001). There was a significant positive relationship between isometric and isokinetic PT FI% ( 0.60, 0.002) as well as between isometric PT FI% and RTD FI% ( 0.40, 0.046). There was no significant relationship observed between RTD FI% and ACC FI% nor between isokinetic PT FI% and ACC FI%.
CONCLUSIONS:
This investigation observed different patterns of response in maximal strength between isometric and isokinetic assessments following a isokinetic fatigue protocol. Additionally, the ability to rapidly generate strength and velocity had significantly different responses to fatigue between isometric and dynamic assessments. Due to these different responses, we recommend that assessments of fatigue at least include the same testing modality as the modality of the fatigue protocol.
Fatigue has been defined as any reduction in the force-producing capacity of a muscle during a maximal voluntary contraction [1], a reduction in the power-production of the muscle, as well as an inability to maintain a given maximal strength level when performing repeated maximal contractions [2]. When research investigations started examining the decline in strength characteristics (i.e., peak torque) during fatiguing isokinetic muscle actions, it was observed that the linearity for the relationships was dependent on the number of repetitions performed [3]. Thorstensson and Karlsson [3] utilized isokinetic strength testing in an attempt to estimate percentages of fast-twitch and slow-twitch muscle fibers. During that study, 10 men performed 50 and 100 repeated maximal concentric isokinetic muscle actions of the leg extensors, and the peak torque values for the first and last three muscle actions were used to calculate participant’s percent decline. Thorstensson and Karlsson [3] concluded that the majority of the decrease in the peak torque values occurred early in the repetition series, and that there was a positive correlation ( 0.86) between the percent decline and the percentage of fast-twitch muscle fibers for the vastus lateralis. Since then, numerous studies [4, 5, 6, 7, 8, 9, 10, 11, 12] have emerged using Thorstensson and Karlsson’s [3] fatiguing isokinetic protocol (50 isokinetic muscle actions at a controlled angular velocity of 180s) to examine muscle function and acute responses between age, sexes and training status.
While decreases in performance may occur post-fatigue, considerable evidence suggests that fatigue is not caused by a single factor, and, that the mechanisms underlying the force reduction are task specific [13, 14]. For instance, a significant amount of research has been conducted on dynamic fatiguing protocols and acute recovery responses (pre vs. post isometric assessment) [15, 16, 17, 18, 19], however, discrepancies in acute responses from fatigue could be related to the different fatigue interventions that were implemented (overall percent fatigue, repetition scheme, time under tension, etc.). For example, Klass et al. [17] examined fatigue responses for isometric peak torque of the triceps surae; however, the investigators utilized a dynamic plantar flexion fatigue intervention. Although the authors observed a significant decrease in peak torque for the plantar flexors, they observed a sizable difference ( 10% reduction) in torque depending on the angle at which the ankle was tested. The implementation of a dynamic assessment prior to and immediately following their intervention may have elucidated the disparity in the isometric fatigue responses they observed. Additionally, Marshall et al. [18] investigated the effects of fatiguing squat protocols on maximal isometric force of the squat. While their study revealed a significant decrease in maximal and rapid force, the implementation of a fatiguing isometric squat protocol may have provided additional information about the fatigue response mechanisms for isometric versus dynamic contractions. Furthermore, it is important to note that muscle power has been shown to be negatively affected more than maximal force following a dynamic fatigue protocol [20] and this reduction in force-producing capacity may lead to deficits in subsequent muscular performance, thus hindering physical ability post-exercise. Consequently, assessing multiple modalities may provide different results and may provide more answers about the mechanisms regarding fatigue responses.
Thus, different modes of assessment (e.g., isometric vs. isokinetic) may reveal dissimilar acute deficits in maximal strength and rapid force characteristics following a dynamic fatigue protocol of the upper extremity. Therefore, the purpose of this investigation was to determine to what extent a Thorstensson fatiguing protocol affects isokinetic (dynamic) and isometric (static) performance characteristics. We hypothesized that all post measurements, independent of testing modality, would be significantly reduced with fatigue. However, we anticipated a difference in the magnitude of the decline. Specifically, we hypothesized that the dynamic assessments for strength and acceleration would yield a significantly greater decline following a dynamic, isokinetic fatigue protocol than static, isometric assessments.
Methods
Participants
Twenty-five college-aged, resistance-trained males (mean SD: age 23.2 2.7 years, height 178.0 6.0 cm, mass 88.5 10.6 kg) volunteered to participate in this study. All participants reported being consistently engaged in a structured resistance-training program involving the upper body for a minimum of 6 months (mean SD: 5.5 3.0 years, 4.7 0.6 days per week) ( 3 times per week) prior to the study. None of the participants reported any current or ongoing musculoskeletal injuries of the upper extremity within the previous 12 months prior to testing. This study was approved by the University Institutional Review Board for human subject’s research, and each participant signed an informed consent document and health history questionnaire prior to testing.
Experimental design
This study was designed to investigate the effects of an isokinetic protocol on isometric and dynamic fatigue indices (FI%) of the elbow flexors (EF). Each participant visited the laboratory on 2 occasions separated by 48–72 hours. During the first visit, participants were familiarized with isometric and isokinetic maximal voluntary contractions (MVCs), as well as the experimental fatigue protocol. During the second visit, participants completed isokinetic and isometric MVCs, in a randomized order, of the EF prior to (pre) and following (post) 50 isokinetic muscle actions at a controlled angular velocity of 180s [3]. Only full range of motion repetitions were counted during the protocols.
Isometric strength assessments
Maximal isometric strength testing was performed on the dominant (right for all participants) arm using a Biodex System 3 isokinetic dynamometer. Participants were seated with restraining straps across the chest and pelvis and the input axis of the dynamometer lever arm was aligned with the axis of rotation of the elbow. Additionally, a Velcro-adjusted elastic band was secured over the upper arm to diminish excessive movement. All isometric torque assessments for the EF were performed with a shoulder angle of 90 in the sagittal plane with an elbow angle of 90 between the arm and forearm [21, 22]. Prior to maximal isometric strength testing, the participants performed a 5-minute warm-up on the upper body ergometer at a pre-determined low-intensity ( 60 revmin). In addition, 3 submaximal isokinetic muscle actions were performed at 60s with approximately 75% of their perceived maximal effort for the EF. Following the sub-maximal contractions and prior to experimental testing, each participant performed 2–3 isometric MVCs of the EF with 1 minute of recovery between each contraction. The participants were verbally instructed to ‘pull,’ “as hard and fast as possible” for a total of 3–4 seconds for all MVCs [23]. This process was also repeated immediately following the experimental fatigue protocol.
Acceleration assessment
Similar to the aforementioned maximal isometric testing procedures, maximal velocity (V) testing was performed to determine maximal limb acceleration (ACC) by setting the isokinetic dynamometer at a velocity of 500s (i.e., velocity of the dynamometer was set above all subjects’ maximum velocity capacities). Each participant performed 3 isokinetic MVCs through 90 of ROM for the EF (starting at 0 to 20 – arm straight out – and finishing at 90 to 110 depending on soft-tissue approximation) with 1 minute of recovery between each contraction. V was used to assess the maximal shortening velocity of the muscle-limb unit in which no resistance (with the exception of the lever arm) was provided throughout the duration of the contraction [24]. The participants were verbally instructed to ‘pull,’ “as fast as possible”. This process was repeated immediately following the experimental fatigue protocol [25, 26].
Experimental protocol
Five minutes following all pre-testing, participants performed the experimental fatigue protocol consisting of 50 continuous repetitions of isokinetic contractions of the EF at 180s (medium velocity) through 90 ROM [4]. Participants were seated with restraining straps identical to all MVC testing procedures. Additionally, a neutral handgrip, with the elbow secured to the arm pad (using a flexible velcro band/strap), was used in order to perform elbow flexion throughout the protocol. During the experimental protocol, participants were asked to provide maximal effort for each muscle action and were verbally encouraged to ‘pull’ as hard as they can throughout the entire protocol and relax on the extension back to the starting position. Experimental testing was terminated at the completion of all 50 maximal repetitions. Verbal encouragement was provided to the participants during the entire protocol. Immediately following (within 10 s) the fatigue protocol MVC assessments were repeated.
Signal processing
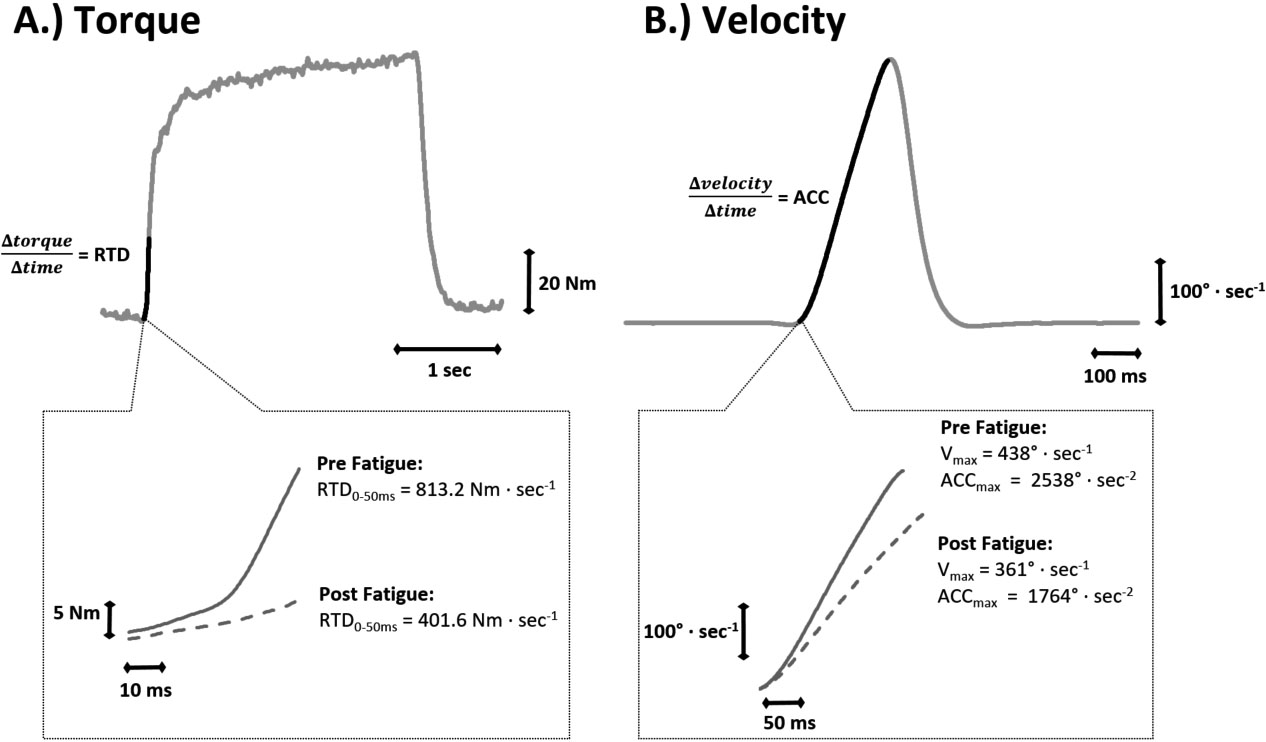
Torque (Nm) and angular velocity (degs) signals were sampled simultaneously at 2 kHz with a Biopac data acquisition system, stored on a personal computer, and processed off-line with custom-written software. The torque signal was smoothed using a 25 ms zero-shift moving average. All subsequent analyses were performed on the scaled and filtered torque signal. Isometric MVC peak torque (PT) was determined as the highest 25 ms epoch during the entire 3–4 s MVC plateau [23, 27]. Isokinetic PT was attained from the first and last 3 reps of the fatigue protocol. Absolute RTD was calculated from the linear slope of the torque-time curve (torque/time) across the interval of 0–50 ms. V (degs) was calculated as the highest velocity attained during the unloaded MVC (500s). ACC (degs) was determined as the 10 ms interval that demonstrated the highest linear slope of the velocity-time curve (velocity/time) from the V contraction (see Fig. 1). These procedures were used to obtain the linear portion of the rate of rise in velocity, while simultaneously excluding the deceleration or “rounding off” of the signal observed at the edge of the velocity plateau. The onset of velocity was determined as the point when the velocity signal reached a threshold 2 degs above baseline [24]. The isometric MVC with the highest PT and RTD were used for all analyses. Isokinetic 180s FI% was calculated using initial and final PT, which consisted of the average of the three muscle actions with the highest and lowest PT values during the fatigue protocol, respectively. All fatigue indices were calculated as “(Final – Initial) Initial 100”.
Example measures pre- and post-fatigue. Plot A shows an example torque signal during an isometric maximal voluntary contraction (MVC). The darker section of the signal highlights the 50 ms epoch used to assess rate of torque development (RTD). The subplot below shows the 50 ms epoch magnified (note the different axis scales) for both the pre-fatigue MVC as well as the post-fatigue MVC from the same subject. Plot B shows an example velocity signal during a dynamic, unloaded MVC (pre-fatigue). The darker section of the signal highlights the onset of movement to peak velocity (V). This highlighted epoch is magnified in the subplot below, and again shows both a pre-fatigue and post-fatigue MVC for the same subject. Acceleration was calculated with a 10 ms moving window, and the highest 10 ms value was considered the peak acceleration (ACC).
Test-retest reliability
Test-retest reliability was determined by intraclass correlation coefficients, SEM, and minimal difference was performed using pretest values from testing days for each dependent variable. The -value represents the type 1 error rate for the one-way repeated measures ANOVA across two visits and SEM is expressed as absolute values and percentages of the mean. Isometric PT displayed the following, -value 0.76, ICC 0.97, SEM 76.35, and SEM% 4.78. Isokinetic PT, -value 0.67, ICC 0.95, SEM 58.55, and SEM% 6.94. RTD, -value 0.62, ICC 0.98, SEM 0.62, and SEM% 4.03. ACC, -value 1.00, ICC 0.98, SEM 0.10, and SEM% 4.53.
Statistical analysis
Four separate paired samples -tests were used for each dependent variable (PT, RTD, ACC, and FI%). Pearson correlation coefficients were computed to assess the relationships between FI% for each dependent variable. PASW software version 21.0 was used for all statistical analyses. An alpha level of 0.05 was considered significant for all comparisons.
Results
Paired samples -tests
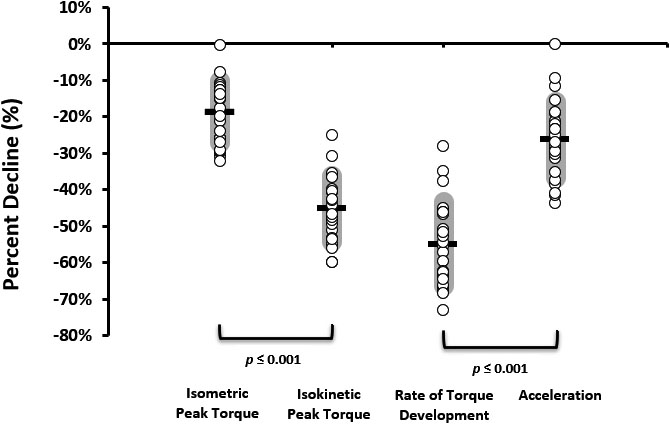
Distribution plot showing individual responses to fatigue for the four dependent variables. Each circle represents an individual subject’s data point. The horizontal black line is the mean, and the shaded areas depict the standard deviations. The brackets denote significant difference between variables ( 0.001).
Means, standard deviations, and distributions for all dependent variables FI% are presented in Fig. 2. Isometric PT FI% (M 18.71, SD 8.38) was significantly lower than isokinetic PT FI% (M 45.08, SD 8.87; 0.001). Additionally, ACC FI% (M 26.27, SD 8.82) was significantly less than isometric RTD FI% (M 54.89, SD 11.48; 0.001). Each of the four dependent variables (isometric PT, isokinetic PT, isometric RTD, and ACC) observed significant decreases with fatigue (all four demonstrated 0.001).
Pearson correlation coefficients
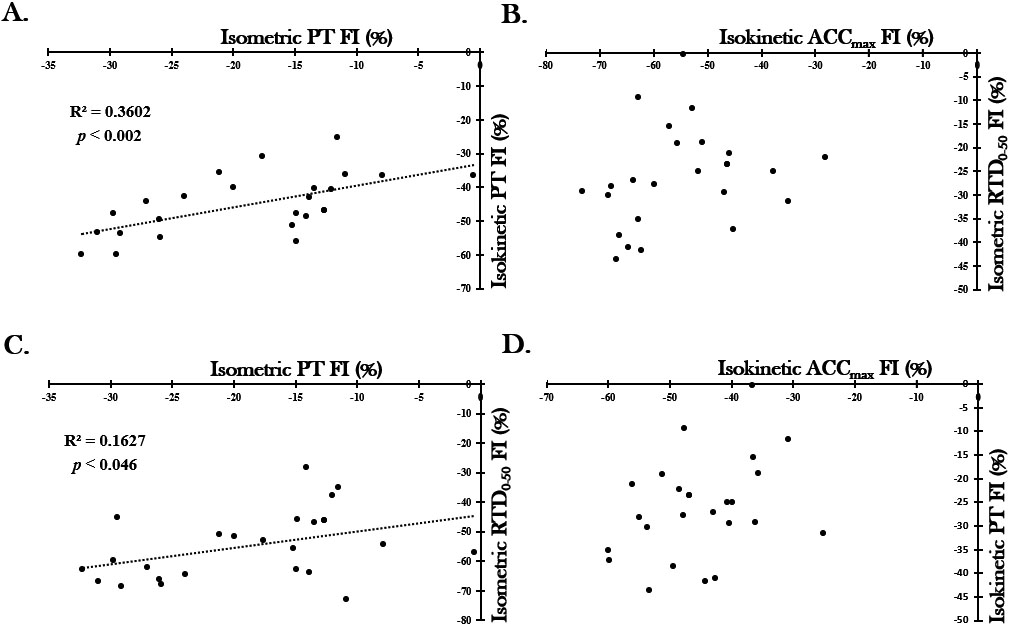
There was a significant positive relationship between isometric PT FI% and isokinetic PT FI% ( 0.60, 0.002) as well as between isometric PT FI% and isometric RTD FI% ( 0.40, 0.046). There was no significant relationship observed between isometric RTD FI% and ACC FI% nor between isokinetic PT FI% and ACC FI% (see Fig. 3).
Correlations between the four dependent variables. Trend lines are shown only for the significant relationships ( 0.05). PT peak torque, FI fatigue index, ACC peak acceleration, RTD rate of torque development.
Discussion
The primary findings of the present study revealed different acute responses between isometric PT FI% and isokinetic PT FI% following a dynamic fatiguing isokinetic protocol (Fig. 2). While different muscles were assessed (quadriceps), the present study’s findings for isometric (18.71%) and isokinetic PT (45.08%) fatigue responses are similar to James et al. [20] who reported (25% vs. 50%) reductions respectively. An important difference to note between isometric and isokinetic contractions, is the overall production of work [28]. Specifically, Motzkin et al. [28] suggested a distinct difference in the physiologic mechanisms of fatigue exists between contractions that produce work and those that do not (i.e. isometric). Furthermore, James et al. [20] proposed the differences in fatigue between the two modes of contraction may be related to the slowing of the contractile properties of the muscle, which is common with fatigued muscles. These responses may be attributed to a slower turnover of cross-bridge cycling rates, negatively affecting shortening velocity and power output, thus, creating larger deficits in isokinetic compared to isometric force production [29, 30]. As a result, this difference may account for the dissimilarity between isometric and isokinetic FI% observed in the current investigation.
Deficits in maximal and rapid torque characteristics as well as ACC have commonly been observed following bouts of isokinetic exercise. In agreement, the present study observed a significant decrease in ACC for the EF immediately following the moderate-velocity protocol. These findings are similar to Nguyen et al. [31], who reported a significant decrease in EF ACC (240s) immediately following an isokinetic eccentric protocol (6 sets of 10 repetitions at 30s). The acute response for ACC for the EF post-fatigue (26.27% declined) could be attributed to muscle fiber type, since the biceps brachii have been reported to be fast-twitch (FT) dominant (57.7%) [32]. Thus, the immediate fatigue-related deficits for ACC may be attributed to greater fatigue for the FT muscle fibers in the biceps brachii. In addition, RTD also observed a significant decrease post-fatigue (54.89%). The immediate deficits in rapid strength may be attributed to cross-bridge cycling rates, muscle fiber type, and calcium kinetics [33, 34, 35, 36, 37]. Unexpectedly, ACC displayed a significantly lower fatigue index than RTD, which was in contradiction of our initial hypothesis. In order to examine this disparity, we normalized RTD to peak torque. However, the normalized RTD FI% (44.53%), although reduced by 10% from absolute RTD, was still significantly higher than the ACC FI% ( 0.001). Thus, taken together, these findings suggest that an isokinetic dynamic fatigue protocol (50 maximal repetitions at 180s) may decrease maximal strength and rapid force capacities to a greater extent compared to acceleration.
While the present investigation observed a dissimilar fatigue response between maximal strength, rapid strength characteristics and acceleration, the overall declines in performance may be due to the effects of peripheral fatigue as a consequence of high-intensity muscular contractions from the dynamic isokinetic protocols [20]. Fatigue is a complex process that may involve both metabolic and neural physiological changes. Mechanisms of neuromuscular fatigue have largely been characterized as being peripheral, likely occurring from an inability to restore Na and K gradients across the sarcolemma resulting in large amounts of K being depleted, thereby leading to impaired action potential conduction efficiency [38]. Additionally, metabolic by-product buildup from increases of inorganic phosphate, ADP, AMP, and H could affect the ATPase activity and thus decrease the reuptake of Ca [39]. The decreased Ca reuptake combined with an inhibition of the t-tubules, which reduces sarcoplasmic reticulum calcium release, may lead to decreased sensitivity at the cross-bridge binding sites [39]. Furthermore, it is possible that increases in acidity levels resulting from the numerous amount of maximal voluntary muscular contractions, may lead to reduced rates of glycogenolysis and creatine phosphate resynthesis [40] as well as stimulation of group III and IV chemoreceptor afferent neurons which have inhibitory effects on the -motoneurons innervating the fatigued muscle [41]. Taken together, these fatigue-related physiological alterations may be a contributing factor to the immediate responses for strength and rapid force characteristics. Future studies should look to incorporate such measures as electromyography, dynamic musculoskeletal ultrasonography, and near-infrared spectroscopy to provide further insight into the effects of fatigue on neurological and muscular properties.
Conclusion
In summary, the present investigation observed dissimilar patterns of response in maximal strength between isometric and dynamic assessments following an isokinetic fatigue protocol. Additionally, the ability to generate strength and velocity rapidly (i.e., RTD and ACC) had significantly different responses to fatigue between isometric and dynamic assessments. Specifically, the deficit observed for isokinetic PT ( 45%) was more than double that seen with the isometric PT ( 19%). Conversely, the deficit observed for isometric RTD ( 55%) was more than double that seen with the dynamic ACC ( 26%). This demonstration could indicate that the differences in sensitivity of the two types of contractions is a major factor accounting for the loss of force and power during exercise. As previously stated, this may be due to the amount of work produced and/or slowing of the contractile properties of the muscle. Thus, it is important that studies utilizing a fatiguing intervention include testing in the same modality as the fatiguing protocol. Knowing that a number of investigators implement various fatigue modalities, the present findings suggest that pre- and post-isometric and dynamic assessments may vary greatly depending on the type of contractions used during the fatiguing protocol.
The findings of the present investigation could have practical and clinical relevance. Due to the results observed in this study, coaches, trainers, clinicians and practitioners who utilize fatigue resistance as an evaluation tool for recovery following training or injury may consider implementing a task specific assessment. Thus, clinicians, practitioners, and strength and conditioning professionals may use caution designing, implementing, and assessing fatigue interventions, as maximal strength and velocity characteristics may reveal differential results depending on the type of contraction utilized.
Author contributions
Conception: Cameron S. Mackey, Ryan M. Thiele and Jason M. DeFreitas.
Performance of work: Cameron S. Mackey.
Data analysis: Cameron S. Mackey and Jason M. DeFreitas.
Manuscript preparation: Cameron S. Mackey, Ryan M. Thiele, David J. Sanders and Jason M. DeFreitas.
Revision for important intellectual content: Cameron S. Mackey, Ryan M. Thiele, David J. Sanders and Jason M. DeFreitas.
Supervision: Cameron S. Mackey and Jason M. DeFreitas.
Ethical considerations
This study was approved by the University Institutional Review Board for human subject’s research (ED-15-153, January 2016), and each participant signed an informed consent document and health history questionnaire prior to testing.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Footnotes
Acknowledgments
The authors have no acknowledgments.
Conflict of interest
None of the authors have any conflict of interest to disclose.
References
1.
GandeviaS. Some central and peripheral factors affecting human motoneuronal output in neuromuscular fatigue. Sports Medicine.1992; 13(2): 93-8.
2.
MathiassenSE. Influence of angular velocity and movement frequency on development of fatigue in repeated isokinetic knee extensions. European Journal of Applied Physiology and Occupational Physiology.1989; 59(1-2): 80-8.
3.
ThorstenssonAKarlssonJ. Fatiguability and fibre composition of human skeletal muscle. Acta Physiol Scand.1976; 98(3): 318-22.
4.
BeckTHoushTJohnsonGWeirJCramerJCoburnJ, et al. Comparison of Fourier and wavelet transform procedures for examining the mechanomyographic and electromyographic frequency domain responses during fatiguing isokinetic muscle actions of the biceps brachii. J Electromyog Kinesiol.2005; 15(2): 190-9.
5.
MackeyCSThieleRMConcholaECDeFreitasJM. Comparison of fatigue responses and rapid force characteristics between explosive- and traditional-resistance-trained males. Eur J Appl Physiol. 2018.
6.
StockMBeckTDeFreitasJ. The effects of diverting activities on recovery from fatiguing concentric isokinetic muscle actions. J Strength Cond Res.2011; 25(7): 1911-7.
7.
StockMBeckTDeFreitasJYeX. Sex comparisons for relative peak torque and electromyographic mean frequency during fatigue. Res Quart Ex Sport.2013; 84(3): 345-52.
8.
KomiPVTeschP. EMG frequency spectrum, muscle structure, and fatigue during dynamic contractions in man. Eur J Appl Physiol Occup Physiol.1979; 42(1): 41-50.
9.
TeschPSjodinBThorstenssonAKarlssonJ. Muscle fatigue and its relation to lactate accumulation and LDH activity in man. Acta Physiol Scand.1978; 103(4): 413-20.
10.
NilssonJTeschPThorstenssonA. Fatigue and EMG of repeated fast voluntary contractions in man. Acta Physiol Scand.1977; 101(2): 194-8.
11.
LarssonLKarlssonJ. Isometric and dynamic endurance as a function of age and skeletal muscle characteristics. Acta Physiol Scand.1978; 104(2): 129-36.
12.
MaffiulettiNAJubeauMMunzingerUBizziniMAgostiFDe ColA, et al. Differences in quadriceps muscle strength and fatigue between lean and obese subjects. Eur J Appl Physiol.2007; 101(1): 51-9.
13.
EnokaRMStuartDG. Neurobiology of muscle fatigue. J Appl Physiol.1992; 72(5): 1631-48.
14.
GandeviaSC. Spinal and supraspinal factors in human muscle fatigue. Physiol Rev.2001; 81(4): 1725-89.
15.
ConcholaECThieleRMPalmerTBSmithDBThompsonBJ. Acute postexercise time course responses of hypertrophic vs. power-endurance squat exercise protocols on maximal and rapid torque of the knee extensors. J Strength Cond Res.2015; 29(5): 1285-94.
16.
ChiuLZFFryACSchillingBKJohnsonEJWeissLW. Neuromuscular fatigue and potentiation following two successive high intensity resistance exercise sessions. Eur J Appl Physiol.2004; 92(4): 385-92.
17.
KlassMGuissardNDuchateauJ. Limiting mechanisms of force production after repetitive dynamic contractions in human triceps surae. J Appl Physiol.2004; 96(4): 1516-21.
18.
MarshallPWMRobbinsDAWrightsonAWSieglerJC. Acute neuromuscular and fatigue responses to the rest-pause method. J Sci and Med in Sport.2012; 15(2): 153-8.
19.
WalkerSDavisLAvelaJHäkkinenK. Neuromuscular fatigue during dynamic maximal strength and hypertrophic resistance loadings. Journal of Electromyography and Kinesiology.2012; 22(3): 356-62.
20.
JamesCSaccoPJonesD. Loss of power during fatigue of human leg muscles. J Physiol.1995; 484(1): 237-46.
21.
BeckTKasishkePStockMDeFreitasJ. Neural Contributions to Concentric vs. Eccentric Exercise – Induced Strength Loss. J Strength Cond Res.2012; 26(3): 633-40.
22.
BilodeauMErbMDNicholsJMJoinerKLWeeksJB. Fatigue of elbow flexor muscles in younger and older adults. Muscle & Nerve.2001; 24(1): 98-106.
23.
ThompsonBJRyanEDSobolewskiEJConcholaECCramerJT. Age related differences in maximal and rapid torque characteristics of the leg extensors and flexors in young, middle-aged and old men. Exp Gerontol.2013; 48(2): 277-82.
24.
ThompsonBJConcholaECPalmerTBStockMS. Effects of aging on maximal and rapid velocity capacities of the leg extensors. Exp Gerontol.2014; 58: 128-31.
25.
ThompsonBJSmithDBSobolewskiEJFiddlerREEverettLKlufaJL, et al. Influence of acute eccentric exercise on the H: Q ratio. Inter J Sports Med.2011; 32(12): 935-9.
26.
AagaardPSimonsenEBAndersenJLMagnussonSPBojsen-MøllerFDyhre-PoulsenP. Antagonist muscle coactivation during isokinetic knee extension. Scand J Med Sci Sports.2000; 10(2): 58-67.
27.
ConcholaECThompsonBJSmithDB. Effects of neuromuscular fatigue on the electromechanical delay of the leg extensors and flexors in young and old men. Eur J Appl Physiol.2013; 113(9): 2391-9.
28.
MotzkinNCahalanTMorreyBAnKChaoE. Isometric and isokinetic endurance testing of the forearm complex. Am J Sports Med.1991; 19(2): 107-11.
29.
HaanAJonesDSargeantA. Changes in velocity of shortening, power output and relaxation rate during fatigue of rat medial gastrocnemius muscle. Pflügers Archiv Eur J Physiol.1989; 413(4): 422-8.
30.
JonesDBigland-RitchieB. Electrical and contractile changes in muscle fatigue. Inter Series Sport Sci.1986; 16: 377-92.
31.
NguyenDBrownLCoburnJJudelsonDEurichAKhamouiA, et al. Effect of delayed-onset muscle soreness on elbow flexion strength and rate of velocity development. J Strength Cond Res.2009; 23(4): 1282-6.
32.
BellemareFWoodsJJohanssonRBigland-RitchieB. Motor-unit discharge rates in maximal voluntary contractions of three human muscles. J Neurophysiol.1983; 50(6): 1380-92.
33.
AagaardPSimonsenEBAndersenJLMagnussonPDyhre-PoulsenP. Increased rate of force development and neural drive of human skeletal muscle following resistance training. Journal of Applied Physiology.2002; 93(4): 1318-26.
34.
AndersenLLAndersenJLZebisMKAagaardP. Early and late rate of force development: Differential adaptive responses to resistance training? Scandinavian Journal of Medicine & Science in Sports.2010; 20(1): e162-e9.
35.
BottinelliRCanepariMPellegrinoMReggianiC. Force-velocity properties of human skeletal muscle fibres: Myosin heavy chain isoform and temperature dependence. The Journal of Physiology.1996; 495(Pt 2): 573.
36.
BrodyIA. Regulation of isometric contraction in skeletal muscle. Experimental Neurology.1976; 50(3): 673-83.
37.
LarssonLMossR. Maximum velocity of shortening in relation to myosin isoform composition in single fibres from human skeletal muscles. The Journal of Physiology.1993; 472(1): 595-614.
38.
GreenHJ. Mechanisms of muscle fatigue in intense exercise. J Sports Sci.1997; 15(3): 247-56.
Bigland-RitchieBDawsonNJohanssonRLippoldO. Reflex origin for the slowing of motoneurone firing rates in fatigue of human voluntary contractions. J Physiol.1986; 379(1): 451-9.