MicroRNA-134 inhibits tumor stem cell migration and invasion in oral squamous cell carcinomas via downregulation of PI3K-Akt signaling pathway by inhibiting LAMC2 expression
Free accessResearch articleFirst published online February, 2020
MicroRNA-134 inhibits tumor stem cell migration and invasion in oral squamous cell carcinomas via downregulation of PI3K-Akt signaling pathway by inhibiting LAMC2 expression
Oral squamous cell carcinoma (OSCC) is the most common malignant neoplasm of the mouth. Some studies have found that multiple microRNAs (miRs) participate in OSCC physiological and pathological processes.
METHODS:
We explored the mechanism of action of miR-134 in OSCC involving the PI3K-Akt signaling pathway. Different bioinformatics methods were used to analyze the potential genes and their related miRs in OSCC. Tumor stem cells were separated from OSCCs through magnetic cell sorting. Regulatory pattern between miR-134 and LAMC2 in OSCC was evaluated by ectopic expression, knockdown and reporter assay experiments. The expression of miR-134, LAMC2, genes in PI3K-Akt signaling pathway, and apoptosis-related genes was detected. Cell proliferation was assessed by MTT assay, cell invasion by scratch test, cell migration by Transwell assay, cell cycle and apoptosis by flow cytometry, and cell growth and migration by xenograft tumor in nude mice. LAMC2 was predicted as the crucial factor related to OSCC using different chip data, and miR-134 was predicted to specifically bind LAMC2 in all five databases.
RESULTS:
Overexpressed miR-134 or silenced LAMC2 was observed to inhibit cell proliferation, migration, invasion of OSCC cells, growth of subcutaneous xenograft in nude mice, as well as promote OSCC cell apoptosis. LAMC2, a target gene of miR-134, decreased following miR-134 promotion, while the PI3K-Akt signaling pathway was inactivated following LAMC2 knockdown. Furthermore, we also observed that the effect of overexpressed miR-134 was enhanced when LAMC2 was knocked down.
CONCLUSIONS:
Taken together, these findings suggest that miR-134-mediated direct downregulation of LAMC2 inhibits migration and invasion of tumor stem cells in OSCC by suppressing the PI3K-Akt signaling pathway.
Oral squamous cell carcinoma (OSCC) is one of most common malignant neoplasm worldwide, and its high mortality results from regional and distant metastases as well as local invasiveness [1]. OSCC could possibly be caused by the decline of apoptosis and immunity and leads to the loss of cell differentiation [2]. In the early stages, OSCC patients generally exhibit no symptoms but minimal pain overlaying its clinical diagnosis and prevention [3]. Although there are modern surgical techniques and therapeutic strategies including cisplatin (DDP), cetuximab, and taxanes, the mortality rates of OSCC remain high in a lot of countries, with an overall five-year survival rate below 50% [4]. Therefore, it is necessary to improve the understanding of biological features and underlying molecular mechanisms of OSCC to develop more effective therapies [5].
Multiple microRNAs (miRs) such as miR-145, miR-445, and miR-338, have been reported to play critical roles in the physiological and pathological processes of OSCC, such as cell proliferation, migration, apoptosis, differentiation, and metabolism [6]. MicroRNA-134 (miR-134) is located on chromosome 14q32 [7], which was reported to promote the proliferation and inhibit migration of lung cancer cells [8]. According to previous screening, miR-134 is highly expressed in Head and neck squamous cell carcinoma (HNSCC) tissue [9]. It was predicted that laminin gamma 2 chain (LAMC2) may be a target gene of miR-134 based on biological prediction site, microRNA.org. LAMC2 is a subunit of the heterotrimeric glycoprotein laminin-332, which consists of 3, 3, and 2 chains and could correlates to the metastatic potential and poor prognosis of leukoplakia, colorectal cancer, and lung adenocarcinoma [10]. LAMC2 is over-expressed in various types of cancer including OSCCs [11]. Through direct phosphorylation and inactivation of pro-apoptotic mediators in OSCC, the abnormal activation of phosphatidylinositol 3-kinase (PI3K)-Akt signaling pathway could inhibit cell death [12]. Furthermore, the PI3K signaling pathway regulates several cellular processes, and activation of PI3K has been linked to mitogenesis, survival, differentiation, migration, invasion and actin cytoskeletal reorganization [13, 14]. As for the major component of PI3K signaling pathway, the inhibition of Akt signaling upregulates LAMC2 protein level, while high LAMC2 suppresses Akt signaling [15]. Therefore, we can assume that miR-134 plays a role in OSCCs through LAMC2-mediated regulation of PI3K-Akt signaling pathway. In the present study, we aimed to explore the biological functions and interaction patterns among miR-134, LAMC2, and PI3K-Akt signaling pathway in OSCC.
Materials and methods
Ethical statements
All animal experiments in this experiment are carried out in accordance with the social norms of the Chinese Animal Ethics Committee. The feeding of all animals in the experiment as well as experimental operation accorded with the animal welfare. The protocol was approved by the Institutional Animal Care and Use Committee of Shanghai Ninth People’s Hospital, College of Stomatology, Shanghai Jiao Tong University School of Medicine.
Gene expression omnibus (GEO) data analysis
GEO (https://www.ncbi.nlm.nih.gov/geo/) is a public functional genomics data repository supporting MIAME-compliant data submissions. In this database, “oral squamous cell carcinoma” was chosen as the key word. The species and research group were set to human and expression profiling chip respectively. The sequencing chip that has larger number of samples and includes both OSCC group and control group was selected. Finally, three data sets of GSE74530, GSE23558 and GSE30784 were selected. According to the description of these data sets in the GEO database, samples of each data set were grouped into control group and OSCC group respectively. The differential expression analysis between control group and OSCC group of three data sets was carried out using R language in “limma” package and the difference threshold was set to logFC 2, Value 0.05. At the same time, the expression infrared image of 35 genes was constructed using the R language in “pheatmap” package.
Venn analysis
Using the online analysis website (http://bioinformati cs.psb.ugent.be/webtools/Venn/), the intersection of elements in multiple data sets was calculated and the Venn diagram was generated.
MiRNA prediction
Five databases were used to predict possible miRNA, including microRNA.org (http://34.236.212.39/micror na/home.do), mirDIP (http://ophid.utoronto.ca/mirDIP/ index.jsp#r), RNA22 (http://cm.jefferson.edu/rna22/Pre computed), TargetScan (http://www.targetscan.org/vert _71/) and MiRWalk (http://129.206.7.150). To predict the regulatory gene, LAMC2 was input in the five databases and the species were set to human. Based on the number differences of prediction result, all prediction results in microRNA.org, the first 100 results in mirDIP, the first 150 results in TargetScan and RNA22 and the first 200 in miRWalk were chosen to construct Venn diagram and analyze the intersection of prediction results from five databases.
OSCC cell lines (Cal27) were derived from squamous cells (the American Type Culture Collection [ATCC], Manassas, VA, USA) of tongue cancer patients. Cal27 cells were incubated in Dulbecco’s Modified Eagle Medium (DMEM) (#11-0550, Biological Industries, Beit Haemek, Israel) with 10% fetal bovine serum (FBS) (15950-017, Beijing ZEPING Bioscience & Technology Co., Ltd., Beijing, China) in a humidified incubator with 5% CO at 37C. To obtain OSCC stem cells, Cal27 cells were cultured in serum-free tumor sphere medium DMEM/F-12 (#12500-062, Gibco, Carlsbad, CA, USA), N2 supplementation (#17502048, Gibco, Carlsbad, CA, USA), 20 ng/mL recombinant human basic fibroblast growth factor (bFGF, #100-18B, PeproTech, Rocky Hill, NJ, USA) and 20 ng/mL epidermal growth factor (EGF, #AF-100-15, PeproTech, Rocky Hill, NJ, USA). The medium was changed to selective medium. Cells were cultured in a culture dish at density of 7.5 10 cell/10 mm. The medium was replaced every other day until the tumor spheres were formed.
Cell sorting
CDl33/CD44 cells were sorted with a MACS separator (130-092-545, Miltenyi Biotec Co., Ltd., Bergisch Gladbach, Germany). The Cal27 cells were treated with trypsin, centrifuged at 1000 r/min for 5 min and counted. Then, the cells were transferred to centrifuge tubes (about 2 10 cells/mL for each tube) and resuspended in 200 L phosphate buffer saline (PBS) (10010023, Thermo Fisher Scientific Inc., Waltham, MA, USA). After an addition of 20 L mouse anti-human CD133 polyclonal antibody (AB1582258, Thermo Fisher Scientific Inc., Waltham, MA, USA) with a final concentration of 15 L/mL, cells were incubated at 4C for 30 min and centrifuged at 1000 r/min for 5 min and then resuspended (2 10 cells/mL). Cell resuspension was added with 20 L of 4 10/mL goat anti-mouse immunoglobulin G (IgG) immunomagnetic beads (YB-22160, Shanghai Yubo Biological Technology Co., Ltd., Shanghai, China), mixed, and then incubated at room temperature for 15 min. The test tube was placed on the magnetic separator for 3 min to remove the supernatant with CD133-negative Cal27 cells, and then DNase release buffer was added to separate the magnetic beads from the Cal27 CDl33-positive cells. After that, the test tube was placed on the magnetic separator for 1 min, and the magnetic beads were adsorbed on the tube wall near the magnetic separator. The supernatant was removed and cells were prepared into cell resuspension (2 10 cells/mL). After an addition of mouse anti-human CD44 monoclonal antibody with a final concentration of 15 L/mL (ab6124, Abcam, Cambridge, MA, USA), cells were incubated at 4C for 30 min and then centrifuged at 1000 r/min for 5 min with the supernatant removed, and prepared into cell resuspension (2 10 cells/mL). The cell suspension was added with 20 L of 4 10/mL goat anti-mouse IgG immunomagnetic beads, mixed, and incubated at room temperature for 15 min. The test tube was placed on the magnetic separator for 3 min to remove the supernatant rich in CD44-negative Cal27 cells, and then DNase release buffer was added to separate the magnetic beads from the CDl33 (CD44-positive cells). The test tube was placed on the magnetic separator for 1 min, and the magnetic beads were adsorbed on the tube wall near the magnetic separator with the supernatant discarded. Cells sorted by magnetic bead were cultured with RPMI 1640 medium containing 10% FBS, 20 ng/mL recombinant human epidermal growth factor (EGF) and 10 ng/mL recombinant human basic fibroblast growth factors (bFGF) in a humidified incubator of 5% CO at 37C.
Immunofluorescence
The round coverslips of CDl33/CD44 and CDl33 /CD44 were made. After cell adherence, the cells were fixed using 4% paraformaldehyde (P6148, Shanghai Benhui Biotec Co., Ltd., Shanghai, China) at room temperature for 15 min, and then cryopreserved at 20C. The cryopreserved round coverslips were taken out and thawed at room temperature. After antigen closure with 10% goat serum, round coverslips were incubated with mouse anti-human CD44 antibody IgG1 and rabbit anti-human CD133 polyclonal antibody overnight at 4C. After cell incubation with an addition of goat anti-rabbit secondary antibody labeled with tetramethyl rhodamine isothiocyanate (TRITC) (BA1089, Boster Biological Technology Co., Ltd., Hubei, China) and goat anti-mouse secondary antibody labeled with fluorescein isothiocyanate (FITC) (BA1101, Boster Biological Technology Co., Ltd., Hubei, China), the nucleus was stained with Hochest (BA1089, Shanghai Gene line bioscience Co., Ltd., Shanghai, China) and observed by fluorescent microscopy (2628, Shanghai Weitu Instrument Technology Development Co., Ltd., Shanghai, China).
Cell grouping and transfection
The sorted cancer stem cells in OSCC were classified into six groups, including blank (cells without treatment), negative control (NC) (cells transfected with miR-134 blank plasmid), miR-134 mimic (cells transfected with miR-134 mimic plasmid), miR-134 inhibitor (cells transfected with miR-134 inhibitor plasmid), siRNA-LAMC2 (cells transfected with siRNA-LAMC2 plasmid) and miR-134 mimic siRNA-LAMC2 (cells transfected with miR-134 mimic and siRNA-LAMC2 plasmids) groups. All plasmids for transfection were purchased from Biomics Biotec Co., Ltd. The cancer stem cells at logarithmic growth phase were seeded in a 6-well plate. When cell confluency reached 30% 50%, the transfection was carried out according to instructions of Lipofectamine 2000 (1168019, Beijing T&L Biological Technology Co., Ltd., Beijing, China). The cells (100 pmol) were diluted and mixed with 250 L serum-free Opti-MEM medium (hz-C2105, Shanghai Hu Zheng Industrial Co., Ltd., Shanghai, China), and then incubated at room temperature for 5 min. A total of 5 L Lipofectamine 2000 were diluted with 250 L serum-free Opti-MEM, mixed, and incubated at room temperature for 5 min. A mixture of the above two substances was incubated at room temperature for 20 min and then cultured in cell culture wells in a humidified incubator of 5% CO at 37C for 6 to 8 h. For subsequent experiment, the cells were cultured for another 24 to 48 hours in complete medium.
Dual-luciferase reporter assay
MicroRNA.org was used to analyze the target genes of miR-134 and to verify whether LAMC2 was a direct target gene of miR-134. The 3’-untranslated region (UTR) of LAMC2 gene (Genbank number: 3918) was artificially synthesized and introduced into pGL3-basic (E1751, Promega Corp., Madison, WI, USA) by endonuclease sites Xho I and Nco I. pGL3-LAMC2-3-wild-type (WT) was constructed and a putative miR-134 binding site in the LAMC2 3’UTR was predicted. pGL3-LAMC2-3’UTR-mutant-type (MUT) was constructed in the same way. Gene synthesis was completed by Beijing OriGene Technology Co., Ltd. The above two luciferase reporter vectors were constructed and independently co-transferred with miR-134 mimic into 293T cells and the activity of luciferase was measured with dual-luciferase reporter assay kits (11402ES60, Shanghai Yeasen Biological Technology Co., Ltd., Shanghai, China). After transfection for 48 h, the cells in each well were added with 100 L Passive Lysis Buffer (PLB), and were slightly shaken for 15 min with the cell lysate collected. The program was set with pre-reading time for 2 s and reading value for 10 s. LARIIStop&Glo Reagent (Promega Corp., Madison, WI, USA) was added with 100 L samples each time. Prepared LARIIStop &Glo Reagent was added into the light pipe or plate (the volume of each sample was 20 L) with cell lysate, and then was put into the bioluminescence detection instrument (Modulus, Turner BioSystems, Sunnyvale, CA, USA). The fluorescence activities of firefly luciferase (LUC) and ranilla luciferase (RL) were detected respectively. The fluorescence activity of RL showed the transfection efficiency of each well and the ratio of the LUC to RL indicated the relative luciferase expression, namely LUC/RL.
Gene expression levels of miR-134, LAMC2, PI3K, Akt, glycogen synthase kinase 3beta (GSK3), P21, caspase 9, Bcl-2-associated X protein (Bax) and B cell lymphoma/lewkmia-2 (Bcl-2) in different types of cells were detected by RT-qPCR. The RNA was extracted from cancer stem cells according to the instructions of Trizol kits (1111, Invitrogen Inc, Carlsbad City, CA, USA). RNA was dissolved by diethylpyrocarbonate (DEPC) treated ultrapure water (A10014, Shanghai Sangon Boitech Co., Ltd., Shanghai, China). The optical density (OD) was measured at 260 nm and 280 nm using ND-1000 ultraviolet/visible spectrophotometer (Thermo Fisher Scientific Inc., Waltham, MA, USA). The concentration was measured and the quality of total RNA was identified. The reverse transcription of RNA was completed by two-step method according to the instructions of reverse transcriptional kits (RR037Q, Takara Biotechnology Ltd., Liaoning, China). The reverse transcription system consisted of 2 L of 5 Prime Script Buffer (for Real Time), 0.5 L Prime Script RT Enzyme Mix I, 0.5 L Oligo dT Primer (50 M), 0.5 L Random 6 mers primer (100 M), and 2 g Total RNA, and RNase Free dHO was added up to 10 L. The reaction was conducted at 42C for 15 min, 85C for 5 s and then 4C forever. The cDNA acquired from reverse transcription was stored at 80C temporarily. Based on SYBR Green method and instructions of the kits (FP205, Tiangen Biochemistry Technology Co., Ltd., Beijing, China), RT-qPCR was performed as follow: pre-denaturation at 95C for 30 s, 40 cycles of denaturation at 95C for 10 s, anneal at 60C for 20 s and extension at 70C for 10 s. The reaction system included 12.5 L SYBR Green Mix, 1 L Forward Primer, 1 L Reverse Primer, and 1 4 L cDNA, and ddHO was added up to 25 L. The real-time fluorescent quantitative PCR (Bio-Rad iQ5, Bio-Rad Company, San Francisco, CA, USA) was applied for detection. U6 was used as the internal reference of miR-134 and GAPDH as the internal reference of other genes. The primer (Table 1) was synthesized by Shanghai Generay Biotech Co., Ltd. (Shanghai, China). The reliability of the PCR reactions was evaluated by melting curve analysis. The threshold cycle (Ct) values were set, and relative expression of every target gene was calculated based on the 2 method [18]. The experiment was conducted three times to obtain the mean value.
Primer sequences for reverse transcription quantitative polymerase chain reaction
Transfected cells in each group were collected into eppendorf (EP) tubes and cracked with appropriate protein lysate (SNM386, Beijing Biolab Technology Co., Ltd., Beijing, China) containing 60% RIPA, 39% sodium dodecyl sulfate (SDS) and 1% protease inhibitor on ice for 30 min, and the tube was centrifuged using high speed freezing centrifuge at 12000 rpm at 4C for 30 min. The supernatant was collected in EP tubes and placed on ice box. The concentrations of proteins were determined using the bicinchoninic acid (BCA) Protein Assay Kit (PC0020, Beijing solarbio science & Technology Co., Ltd., Beijing, China). Subsequently, 10% separation gel and 5% spacer gel were prepared using SDS-polyacrylamide gel electrophoresis (SDS-PAGE) Gel Kit. After electrophoresis on polyacrylamide gel, proteins were transferred onto the polyvinylidene fluoride (PVDF) membrane by wet-transfer method, and blocked with 5% bovine serum albumin (BSA) for 1 h with the sealing fluid removed. After that, the membrane was kept in plastic groove, and added with primary antibodies at corresponding concentrations prepared by 5% BSA: rabbit anti-human LAMC2 polyclonal antibody (1: 1000, ab96327), rabbit anti-human PI3K polyclonal antibody (1: 1000, ab1678), rabbit anti-human Akt polyclonal antibody (1: 500, ab8805), rabbit anti-human GSK3 monoclonal antibodies (1: 5000, ab32391), rabbit anti-human phosphorylase Akt (p-Akt) monoclonal antibody (1: 1000, ab38449), rabbit anti-human phosphorylase GSK3 (p-GSK3) monoclonal antibody (1: 500, ab75745), rabbit anti-human P21 monoclonal antibody (1: 1000, ab109520), rabbit anti-human P27 polyclonal antibody (1: 500, ab215434), rabbit anti-human caspase 9 polyclonal antibody (1 g/mL, ab25758), rabbit anti-human Bax polyclonal antibody (1: 1000, ab53154), and rabbit anti-human Bcl-2 polyclonal antibody (1: 1000, ab59348). All antibodies were purchased from Abcam (Cambridge, MA, USA). The membrane was incubated in a refrigerator at 4C overnight with transferring surface up and then washed with Tris-buffered saline containing Tween 20 (TBST) three times in the next day (10 min each time). In the same way, the membrane was incubated at 4C for 4 6 h with an addition of goat anti-rabbit IgG (1: 2000, ab6721, Abcam Company, UK) secondary antibody marked by horse radish peroxidase. The chemiluminescence reagent A and B (PE0010-25, Beijing Solarbio Science & Technology Co., Ltd., Beijing, China) were mixed as 1:1 and then the intermixture was added to membrane, followed by development using a developer. The results were observed under X-ray machine (36209ES01, Shanghai Qcbio Science&Technologies Co., Ltd., Shanghai, China). The ratio between gray values of target protein bands and internal control protein bands served as the relative expression of proteins. The mean value was calculated based on the average of three replicates.
When cell confluency reached about 80%, the transfected cancer stem cells (20 L) in each group were inoculated in a 96-well plate with 5 10 cells/L. After the incubation for 24 h, 48 h, and 72 h respectively, each well was added with 20 L MTT solution (5 g/L). After further culture for 4 h, the supernatant medium was discarded. Each well was added with 150 L dimethyl sulfoxide (DMSO), and oscillated to completely mix. After 10 min, OD (A) in each well at 490 nm was measured by a multi-function microplate reader (CLARIOstar, Bio-Gene Technology Limited, Hong Kong, China) so as to reflect the vitality of cell growth. The cell growth curve was made.
Scratch test
After transfection for 48 h, cancer stem cells in each group were collected. Cell suspension (20 L) was seeded in a 6-well plate with the density of 1 10 cells/well. When cell confluency reached 90%, 4 horizontal lines and 4 vertical streaks were drawn at the bottom of each well by a sterile tip head (200 L) under the same strength. After the removal of the origin medium, each well was incubated with 2 mL medium with 10% serum. The images were respectively acquired using an inverted microscope at 0 h and 24 h after scratching, and the widths (D) of multiple scratches were measured. The healing rate of each group (D D)/D 100%. The experiment was conducted 3 times to obtain the mean. The cell migration ability of each group was compared.
Transwell assay
Transwell assay was performed to analyze the invasion ability of cell. The upper chamber of Transwell chamber was carpeted with Matrigel gel (356234, BD Bioscience, San Jose, CA, USA) to simulate cell matrix. During the preparation phase, Matrigel gel was diluted with serum-free RPMI 1640 medium (C22400500BT, Beijing Biopeony Technology Co., Ltd., Beijing, China) containing 0.2% BSA at the ratio of 1:5. The upper chamber of transwell chamber was added with 50 L Matrigel and placed in an incubator overnight to solidify Matrigel. The next day, 100 L serum-free RPMI 1640 medium with 0.2% BSA and 600 L serum-free RPMI 1640 medium containing 10% FBS were added to the upper and lower chambers of Transwell chamber respectively for hydration for 30 min. After conventional digestion, cells in each group were resuspended with RPMI 1640 medium at the density of 1 10 cells/mL. A total of 200 L cell suspensions were added to the upper chamber of transwell chamber carpeted with Matrigel gel. After a conventional culture for 24 h, the cells on the upper chamber were removed by cotton bud, fixed with 4% polyoxymethylene for 15 min, stained with 0.5% crystal violet solution (prepared with methanol for 15 min). Images in five random visual fields were acquired with an inverted microscope (XDS-800D, Shanghai Caikon Optical Instrument Co., Ltd., Shanghai, China) (200 ). The cells that had transferred to the lower chamber were counted. Three duplicate wells were prepared for each group. The mean value was calculated based on the average of three replicates.
Flow cytometry
After transfection for 48 h, the cells were detached with trypsin without ethylene diamine tetraacetic acid (EDTA), collected into a flow tube, and centrifuged with supernatant removed. After that, the Annexin-V-FITC/PI was prepared with Annexin-V-FITC, Propidium Iodide (PI), and Hepes buffer solution at the ratio of 1:2:50 based on the instructions of Annexin-V-FITC cell apoptosis detection kit (88-8005-72, Shanghai Yanhui Biotech Co., Ltd., Shanghai, China). Each 100 L dye liquor was used to resuspend 1 10 cells, shaken, and mixed thoroughly. After incubation at room temperature for 15 min, cells were added with 1 mL HEPES buffer solution, shaken, and mixed thoroughly. After that, cells were placed in the flow cytometer (FACSCalibur, BD Biosciences, Franklin Lakes, NJ, USA) to record excitation wavelength. Cell cycle and apoptosis were detected by red fluorescence light at the wavelength of 488 nm. The experiment was conducted three times.
Enograft tumor in nude mice
A total of 36 specific pathogen free (SPF) BALB/c-nu/nu nude mice (weighing 18 22 g) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). All nude mice were randomly assigned into six groups with six mice per group: blank group, NC group, miR-134 mimic group, miR-134 inhibitor group, siRNA-LAMC2 group, and miR-134 mimic siRNA-LAMC2 group. Nude mice in each group were injected with corresponding transfected cells. A total of 0.2 mL single cell suspension (1 10 cells) was injected into the back of nude mice by hypodermic. After inoculation, the mice were fed in animal experimental center with free access to food and water at a constant temperature of 25C 26C and humidity (about 70%) with the illumination time from 6 am to 6 pm. The center was regularly disinfected by ultraviolet with good ventilation. The mice were thoroughly observed every day, whereby special attention was focused on the activity, diet, defecation and skin color of mice. The maximum diameter (L) and minimum diameter (W) of the tumors that appeared in mice were measured using a vernier caliper. The volume of tumor was calculated according to the formula, V W L 0.52 [19]. The size of the tumor and the weight of the mice were measured every week. Thirty-five days later, the mice were killed using cervical dislocation method and the transplanted tumor was removed.
Hematoxylin-eosin (HE) staining
The maximum surface of tumor tissues of the mice in each group was acquired with a thickness of 2 3 mm and the part with overmuch necrotic tissue were avoided. The slices cut from the tumors were stored in 10% formalin, fixed for over 24 h, conventionally dehydrated with 75%, 80%, 90%, 90%, 100% alcohol respectively twice (1.5 h per time), and then cleaned twice (8 min per time) in dimethylbenzene. After that, the tissue blocks were embedded. Then the paraffin-embedded tissue blocks were cut into 5 7 m sections using the slicer (RM2016, Shanghai Leica Instrument Co., Ltd., Shanghai, China). The sections were baked at 55C, dewaxed in dimethylbenzene twice (5 min each time), and dehydrated in 100%, 95%, 85%, 75% alcohol (3 min each concentration). The sections were stained with hematoxylin for 8 min, differentiated with hydrochloric acid alcohol for 5 s, and added with 0.25% ammonia for 1 min to turn back to blue. After that, the sections were stained with eosin for 30 s, dehydrated twice in 85%, 95% and 100% alcohol respectively (2 min each time), and cleared twice with dimethylbenzene (5 min each time). After the dyeing process, neutral balsam was dropped on the slides and then the sections were mounted by clean coverslip. The microscope (XSP-2C, Shanghai Bingyu Optical Instrument Co., Ltd., Shanghai, China) was applied to observe the pathological changes of the tumor.
Statistical analysis
All statistical analysis was conducted by using SPSS 21.0 (IBM Corp., Armonk, NY, USA). Measurement data were expressed as mean standard deviation. Differences between two groups were compared by test while differences among multiple groups were compared by one-way analysis of variance. Enumeration data were expressed in the form of percentage and differences were analyzed by chi-square test. 0.05 indicated statistical significance.
LAMC2 is a target gene of miR-134. A–C, Infrared images of top 35 differential genes in GSE74530, GSE23558 and GSE30784 data sets, respectively. The top bar indicates the types of samples; the blue part represents normal control samples and the red part represents tumor samples. The color gradation on the upper right represents gene expression levels; the color changed from red to green, which means the expression levels changed from high to low. The abscissa represents the sample number and the ordinate indicates the gene name. Each block in the diagram indicates the expression of a gene in a sample; the dendrogram on the left side represents the clustering analysis based on the expression differences of genes; D, The Venn analysis of top 50 differential genes of the three chip data sets. The blue part represents the GSE74530 data set, the red part represents GSE23558 data set, the green part represents the GSE30784 data set, and the area labeled by the blue arrow represents the intersection of the three data sets. E, Prediction of miRNA which regulates LAMC2. The regions of blue, red, brown, yellow and green represents the prediction results of microRNA.org database, mirDIP database, RNA22 database, TargetScan database and mirDIP database, respectively; the middle part labeled by the blue arrow represents the intersection of the five data sets; miR-135, microRNA-135.
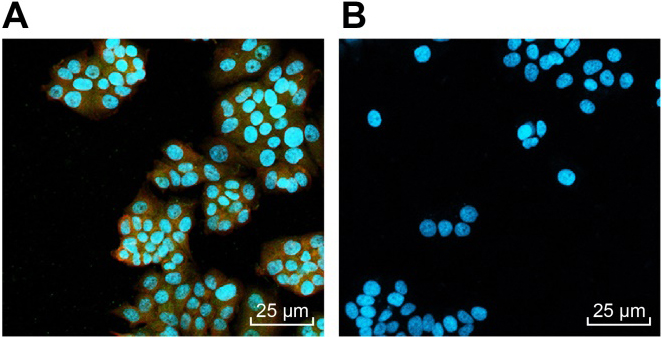
CDl33/CD44 cells are OSCC stem cells. A, CDl33 and CD44 cells shows red fluorescence and green fluorescence under the fluorescence microscope (40 ), respectively; B, CDl33 and CD44 cells exhibits no red or green fluorescence under the fluorescence microscope (40 ).
LAMC2 is negatively regulated by miR-134. A, The presumed binding site between 3’UTR of LAMC2 and miR-134; B, A combination of LAMC2-wt and miR-134 significantly reduces the luciferase activity while a combination of LAMC2-mut and miR-134 does not change the luciferase activity. C, Gene expression levels of LAMC2 analyzed by RT-qPCR; D and E, the protein level of LAMC2 analyzed by western blotting; , 0.05, compared with the NC group; LAMC2, human laminin 2 chain gene; NC, negative control; miR-134, microRNA-134; GAPDH, glyceraldehyde-3-phosphatedehydrogenase; WT, wild type; MUT, mutant type.
Results
miR-134 may regulate LAMC2-mediated PI3K/AKT signaling pathway to affect OSCC
The data of selected sequencing chips of OSCC were analyzed using GEO database. The analysis results showed that many differentially expressed genes (DEGs) were acquired in the GSE74530 data set, the GSE23558 chip data and the GSE30784 data set. Figure 1A–C showed the expression infrared images of top 35 genes of the three databases. In order to further select related genes of OSCC, Venn analysis of the first 50 differential genes of the three data sets was conducted to find their intersection (Fig. 1D). The results showed LAMC2 and MMP1 exited in the intersection of the first 50 differential genes of the three data sets. According to current researches, MMP1 has been found to be associated with a variety of tumor diseases, including OSCC [20, 21, 22, 23]. LMAC2 was rarely reported in OSCC. Only a small number of literatures have pointed out that LAMC2 is associated with OSCC. The specific mechanism of LAMC2 in OSCC is not reported yet. The analysis of OSCC chips data showed that LAMC2 showed significantly high expression in OSCC samples in three microarrays. Further retrieval of the signal pathway related to OSCC showed that the PI3K-Akt signaling pathway may be closely related to the development of OSCC [24, 25, 26]. However, it is still necessary to further study whether LAMC2 affects the PI3K-Akt signaling pathway in OSCC at present. In order to have a better understanding of the role of LAMC2 in OSCC, the miRNAs which regulated LAMC2 was predicted in five databases including microRNA.org, mirDIP, RNA22, TargetScan and miRWalk. Venn analysis was conducted to find the intersection among the forecast results from five databases including all results of microRNA.org, the first 100 results of mirDIP, the first 150 miRNAs predicted by TargetScan, the first 150 miRNAs with the lowest folding energy predicted by RNA22 and the first 200 prediction results of miRWalk. Eventually, only hsa-miR-134-5p (hsa-miR-134) was found to exist in the intersection. These results implied that hsa-miR-134 could regulate LAMC2 in OSCC, and then influence the progression of OSCC through PI3K-Akt signaling pathway.
CDl33/CD44 cells are OSCC stem cells
Magnetic bead sorting was conducted to sort cancer stem cells from OSCC. Results (Fig. 2A and B) indicated that two sorted cell subgroups CDl33/CD44 and CDl33/CD44 were all adhered to the wall and the proliferation was quite active. Under the phase contrast microscopy, these two types of cells were round or polygonal, but CDl33/CD44 cells exhibited obvious cell protuberances with larger cell body. CD133 presented red fluorescence and CD44 presented green fluorescence under the fluorescence microscope. The results showed that the selected CDl33/CD44 cells were able to present red fluorescence and green fluorescence, suggesting the high expression of CDl33 and CD44 of these cells (Fig. 2A). However, no red or green fluorescence was found in CDl33/CD44 cells, suggesting that both CDl33 and CD44 were hardly expressed in these cells (Fig. 2B). The results showed that CDl33/CD44 cells were cancer stem cells.
miR-134 suppresses LAMC2 and mediates PI3K-Akt signaling pathway in tumor stem cells. A, The gene expression levels of miR-134, LAMC2, PI3K and Akt determined by RT-qPCR; B and C, the protein levels of LAMC2, PI3K and Akt, and the extent of Akt phosphorylation determined by western blot; , 0.05, compared with the blank group and the NC group; , 0.05 compared with the miR-134 mimic group and the siRNA-LAMC2 group; NC, negative control; miR-134, microRNA-134; GAPDH, glyceraldehyde-3-phosphatedehydrogenase; LAMC2, human laminin 2 chain gene; PI3K, phosphoinositide 3-kinase.
MTT assay suggests that miR-134 suppresses cell proliferation of OSCC tumor stem cells via LAMC2. , 0.05, compared with the blank group and the NC group; , 0.05, compared with the miR-134 mimic group and the siRNA-LAMC2 group; NC, negative control; miR-134, microRNA-134; LAMC2, human laminin 2 chain gene; PI3K, phosphoinositide 3-kinase.
The scratch test and transwell assay show that miR-134 suppresses OSCC migration and invasion through PI3K-Akt signaling pathway. A and B, cell migration ability in different groups; C and D, cell invasion ability in different groups; , 0.05, compared with the blank group and the NC group; , 0.05, compared with the miR-134 mimic group and the siRNA-LAMC2 group; NC, negative control; miR-134, microRNA-13; LAMC2, human laminin 2 chain gene.
Upregulated miR-134 inhibits cell proliferation but promotes OSCC apoptosis through its effect on related genes. A, the gene expression levels of GSK3, P21, P27, caspase-9, Bax and Bcl-2 determined by RT-qPCR; B and C, the protein levels of GSK3, P21, P27, caspase-9, Bcl-2 and Bax and the extent of GSK3 phosphorylation; , 0.05, compared with the blank group and the NC group; , 0.05, compared with the miR-134 mimic group and the siRNA-LAMC2 group; NC, negative control; miR-134, microRNA-134; LAMC2, human laminin 2 chain gene; GSK3, glycogen synthase kinase-3; Bcl-2, B cell lymphoma 2; Bax, Bcl-2-associated X protein.
miR-134 targets LAMC2 and negatively regulates its expression
The binding region between LAMC2 and miR-134 were predicted by biological prediction website (http://www.microrna.org). To verify the target relationship of LAMC2 and miR-134, the dual-luciferase reporter assay was performed and the results are shown in Fig. 3A and B. Firstly, compared with the NC group, the luciferase activity of wild type 3’UTR of LAMC2 was significantly inhibited by miR-134, while the luciferase activity of mutant 3’UTR was not inhibited. It suggested that miR-134 is able to specifically bind to LAMC2.
The expression differences of LAMC2 between the NC group and the miR-134 mimic group detected by RT-qPCR and western bolt analysis are shown in Fig. 3C–E. The mRNA and protein levels of LAMC2 in the miR-134 mimic group were significantly decreased compared with the NC group ( 0.05). The results suggested that upregulation of miR-134 could specifically inhibit the expression of LAMC2.
Upregulated miR-134 inactivates the PI3K-Akt signaling pathway by inhibiting LAMC2 in OSCC cells
The expression of miR-134, LAMC2, PI3K, Akt and the extent of Akt phosphorylation in each transfected group were detected by RT-qPCR and western blotting and the results are shown in Fig. 4. The results showed that there was no difference between the blank group and the NC group ( 0.05). In comparison with the blank and NC group, the mRNA and protein levels of LAMC2, PI3K, and Akt and the extent of Akt phosphorylation in the miR-134 mimic group, the siRNA-LAMC2 group, and the miR-134 mimic siRNA-LAMC2 group were significantly decreased; miR-134 expression were remarkably increased ( 0.05) in the miR-134 mimic group and the miR-134 mimic siRNA-LAMC2 group; the mRNA and protein expressions of LAMC2, PI3K and Akt were significantly increased and the extent of Akt phosphorylation was increased in the miR-134 inhibitor group ( 0.05). Compared with the miR-134 mimic group and the siRNA-LAMC2 group, the miR-134 mimic siRNA-LAMC2 group exhibited downregulated mRNA and protein expressions of LAMC2, PI3K and Akt as well as the extent of Akt phosphorylation (all 0.05). The results suggested that increase of miR-134 is able to inhibit LAMC2 as well as the activation of the PI3K-Akt signaling pathway.
miR-134 inhibits OSCC cell proliferation via LAMC2 downregulation
Cells in each group were seeded in a 96-well plate and cell proliferation was observed by MTT assay. The results (Fig. 5) suggested that all groups showed no obvious difference in proliferation after 24 h ( 0.05). After 48 h and 72 h, compared with the blank group and the NC group, the cell growth rate of cancer stem cells in the miR-134 mimic siRNA-LAMC2 group, the miR-134 mimic group and the siRNA-LAMC2 group slowed significantly ( 0.05) while that of cancer stem cells in the miR-134 inhibitor group quickened remarkably ( 0.05). In comparison with the miR-134 mimic and siRNA-LAMC2 groups, the growth rate of tumor stem cells in the miR-134 mimic siRNA-LAMC2 group was significantly slower ( 0.05). These findings provided evidence that the increase of miR-134 inhibits the proliferation of tumor stem cells by inhibiting LAMC2.
miR-134 suppresses cell migration and invasion of OSCC tumor stem cells via LAMC2 downregulation
Scratch test and transwell assay were conducted to detect effects of miR-134 on cell migration and cell invasion respectively. The results of scratch test are shown in Fig. 6. Cell migration after 48 h did not differ between the blank group and the NC group ( 0.05). Compared with the blank group and the NC group, cell migration rates of the miR-134 mimic group, the siRNA-LAMC2 group and the miR-134 mimic siRNA-LAMC2 group were significantly reduced ( 0.05), while the cell migration rate of the miR-134 inhibitor group was remarkably increased ( 0.05). In comparison with the miR-134 mimic group and the siRNA-LAMC2 group, the miR-134 mimic siRNA-LAMC2 group exhibited reduced cell migration rate ( 0.05). The results suggested that miR-134 is a negative regulator of cell migration via LAMC2.
Results of transwell assay are shown in Fig. 6. There was no significant difference between the blank group and the NC group ( 0.05). Compared with the blank group and the NC group, the cell invasion of the miR-134 mimic group, the siRNA-LAMC2 group, and the miR-134 mimic siRNA-LAMC2 group was significantly reduced ( 0.05), while the cell invasion of the miR-134 inhibitor group was remarkably increased ( 0.05). In comparison with the miR-134 mimic group and the siRNA-LAMC2 group, the miR-134 mimic siRNA-LAMC2 group showed significantly decreased cell invasion ( 0.05). From these results, we could conclude that through negatively regulating LAMC2, miR-134 inhibits cell invasion.
Upregulated miR-134 inhibits cell proliferation but promotes cell apoptosis through its effect on related genes
In order to further explore the effect of miR-134 on proliferation and apoptosis, expression of GSK3, the extent of GSK3 phosphorylation, P21, P27, caspase 9, Bax and Bcl-2 in each transfected group was detected by RT-qPCR and western blotting. The results (Fig. 7) revealed that no statistical difference was found between the blank group and the NC group ( 0.05). Compared with the blank group and the NC group, the miR-134 mimic group, the siRNA-LAMC2 group and the miR-134 mimic siRNA-LAMC2 group greatly increase the expression of P21, P27, caspase 9, GSK3 and Bax, but reduced the expression of Bcl-2 at mRNA and protein levels of, while enhanced the extent of GSK3 phosphorylation (all 0.05). At the mRNA and protein levels, P21, P27, caspase 9, GSK3 and Bax were significantly decreased, but Bcl-2 were significantly increased, while the extent of GSK3 phosphorylation was decreased in the miR-134 inhibitor group (all 0.05). There was no statistical difference between the miR-134 mimic group and the siRNA-LAMC2 group ( 0.05). Compared with the miR-134 mimic group and the siRNA-LAMC2 group, the mRNA and protein levels of P21, P27, caspase 9, GSK3 and Bax were significantly elevated, the mRNA and protein levels of Bcl-2 were significantly decreased and the extent of GSK3 phosphorylation was upregulated in the miR-134 mimic siRNA-LAMC2 group ( 0.05). The results showed that the increase of miR-134 could inhibit cell proliferation but promotes cell apoptosis through its effect on related genes.
The flow cytometry analysis of PI staining indicates that miR-134 inhibits cell cycle entry in OSCC stem cells through PI3K-Akt signaling pathway. A, the DNA content of PI-stained cells at the G0/G1, S, and G2/M phases; B, the percentage of PI-stained cells at the G0/G1, S, and G2/M phases; , 0.05, compared with the blank group and the NC group; , 0.05, compared with the miR-134 mimic group and the siRNA-LAMC2 group; LAMC2, human laminin 2 chain gene; NC, negative control; miR-134, microRNA-134; G, Gap phase; S, synthesis phase.
The flow cytometry analysis of annexin V-FITC/PI staining indicates that miR-134 promotes OSCC stem cell apoptosis through LAMC2-dependent PI3K-Akt signaling pathway. A, tumor stem cells in the scatter plots in which the upper left quadrant identifies the necrotic cells (annexin V/PI), the upper right quadrant identifies the late apoptotic cells (annexin V/PI), the lower left quadrant identifies the live cells (annexin V/PI), and the lower right quadrant identifies the early apoptotic cells (annexin V/PI); B, the percentage of early and late apoptotic cells; , 0.05 compared with the blank group and the NC group; , 0.05 compared with the miR-134 mimic group and the siRNA-LAMC2 group; LAMC2, human laminin 2 chain gene; NC, negative control; miR-134, microRNA-134; PI, Propidium Iodide.
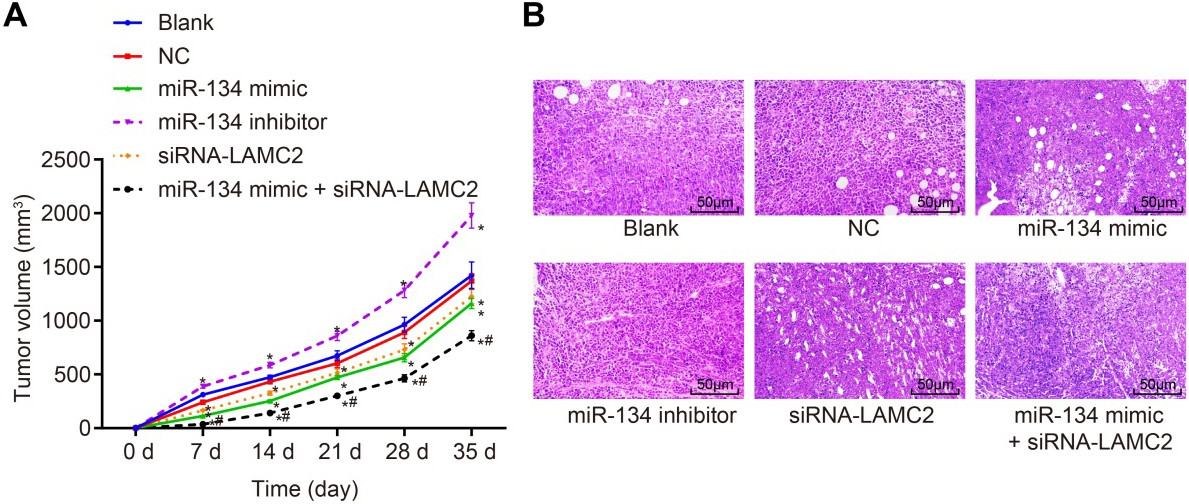
miR-134 inhibits OSCC tumor growth and metastasis via LAMC2. A, Growth curve of subcutaneous tumor of nude mice ( 6); B, HE staining (400 ) of the subcutaneous tumor of nude mice; , 0.05, compared with the blank group and the NC group; , 0.05, compared with the miR-134 mimic group and the siRNA-LAMC2 group; LAMC2, human laminin 2 chain gene; NC, negative control; miR-134, microRNA-134.
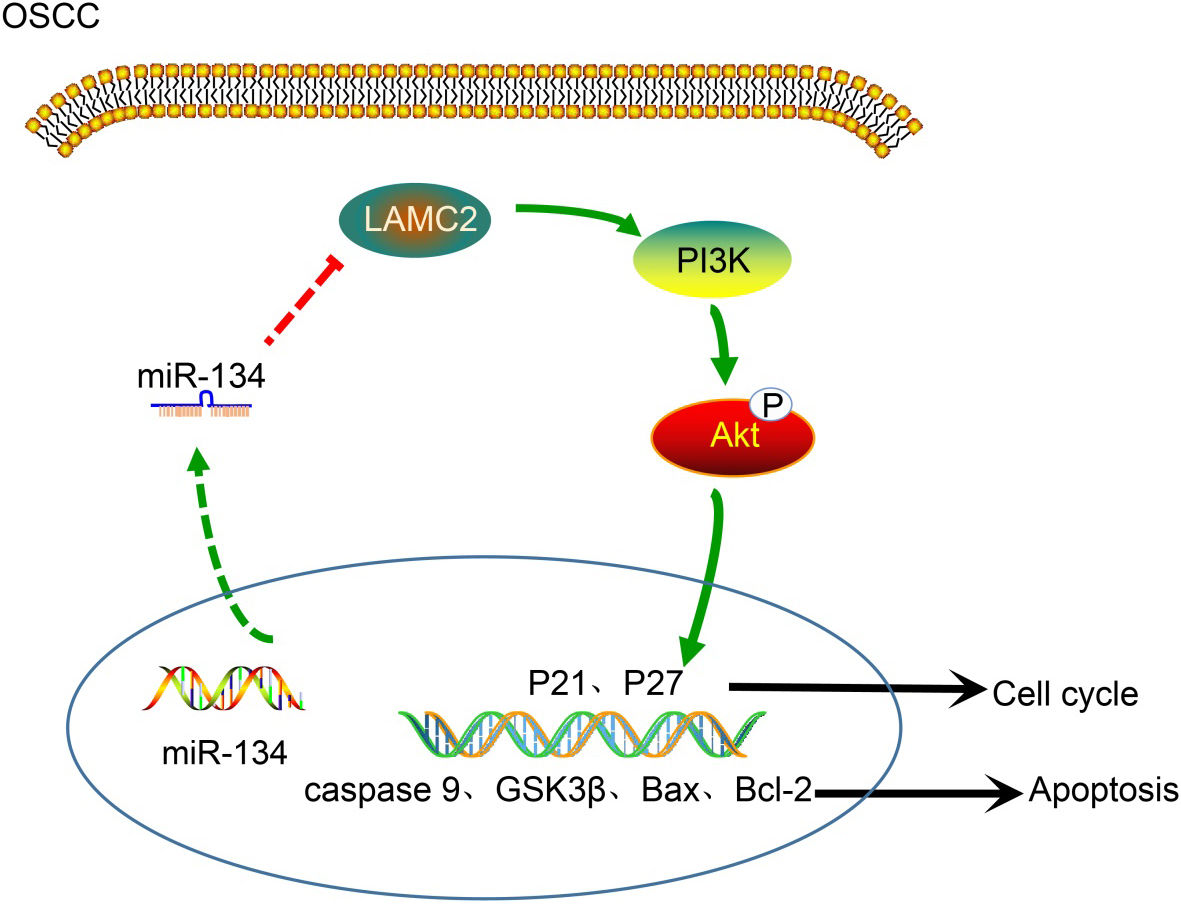
miR-134 blocks PI3K-Akt signaling pathway by inhibiting LAMC2, thus inhibiting cell proliferation and metastases of tumor stem cells in OSCC. In oral squamous cell carcinoma, overexpression of miR-134 decreased the expressions of LAMC2, PI3K, Akt, and p-Akt and significantly raised the expressions of P21, P27, caspase 9, Bax, GSK3, the extent of GSK3 phosphorylation. Besides, the expression of Bcl-2 is reduced, the proliferation of tumor stem cells obviously slowed and its apoptosis rate increased; miR-134, microRNA-134; LAMC2, human laminin 2 chain gene; GSK3, glycogen synthase kinase-3; Bcl-2, B cell lymphoma 2; Bax, Bcl-2-associated X protein.
LAMC2 is responsible for the inhibitory effect of miR-134 on OSCC stem cell cycle entry
PI single staining was conducted to determine cell cycle, and the results are shown in Fig. 8. Compared with the blank group and the NC group, the G0/G1 phase of tumor stem cells was lengthened and the S phase was shortened in the miR-134 mimic group, the siRNA-LAMC2 group and the miR-134 mimic siRNA-LAMC2 group ( 0.05), while the G0/G1 phase of tumor stem cells was shortened and the S phase was lengthened in the miR-134 inhibitor group (all 0.05). Compared with the miR-134 mimic group and the siRNA-LAMC2 group, the G0/G1 phase of tumor stem cells was lengthened and the S phase was shortened in the miR-134 mimic siRNA-LAMC2 group ( 0.05). The results elucidates that miR-134 upregulation inhibits LAMC2 to block cell cycle of tumor stem cells.
LAMC2 is responsible for the contributory effect of miR-134 on OSCC stem cell apoptosis
Cell apoptosis in each group was detected by annexin V-FITC/PI staining, and the results are shown in Fig. 9. In comparison with the blank group and the NC group, the apoptosis rates of tumor stem cells were increased in the miR-134 mimic group, the siRNA-LAMC2 group and the miR-134 mimic siRNA-LAMC2 group ( 0.05), while that was decreased in the miR-134 inhibitor group (all 0.05). The apoptosis rate of tumor stem cells in the miR-134 mimic siRNA-LAMC2 group was significantly higher than that in the miR-134 mimic group and the siRNA-LAMC2 group (all 0.05). The results suggested that the increase of miR-134 could inhibit the expression of LAMC2, thus promoting the apoptosis of tumor stem cells.
LAMC2 is responsible for the inhibitory effect of miR-134 on OSCC tumor growth and metastasis
Effects of miR-134 on tumor growth and metastasis were evaluated by conducting Xenograft tumor in nude mice and HE staining, and the tumor growth curve is shown in Fig. 10A. Compared with the blank group and the NC group, the volume of subcutaneous tumor was obviously smaller in the miR-134 mimic, siRNA-LAMC2 and miR-134 mimic siRNA-LAMC2 groups ( 0.05), while the volume of subcutaneous tumor was significantly larger in the miR-134 inhibitor group ( 0.05). Compared with the miR-134 mimic group and the siRNA-LAMC2 group, the miR-134 mimic siRNA-LAMC2 group showed significantly smaller subcutaneous tumor ( 0.05). The results showed that the increase of miR-134 decreases LAMC2 to inhibit the tumor growth.
The morphological changes of tumor in different groups were observed using HE staining, and the results are shown in Fig. 10B. The tumor cells in the blank group, the NC group and the miR-134 inhibitor group showed close arrangement, active growth and invasion of surrounding tissues. However, the tumor cells in groups of miR-134 mimic, siRNA-LAMC2 and miR-134 mimic siRNA-LAMC2 were relatively loose with different sizes and shapes. HE staining showed that the increase of miR-134 could inhibit the expression of LAMC2 as well as the activation of the PI3K-Akt pathway to inhibit the metastasis of tumor cells.
Discussion
OSCC is one of the most common occurring solid cancers worldwide [27]. There is evidence demonstrating that alteration of miRNAs participates in the development of OSCC [28, 29]. In this study, we aim to explore the role of miR-134 in OSCC. Our study suggested that miR-134 suppresses invasion and migration of tumor stem cells in OSCC by inhibiting the activation of PI3K-Akt signaling pathway via LAMC2.
Our study demonstrated that overexpression of miR-134 inhibits growth, proliferation, migration and invasion but promotes apoptosis of tumor stem cells in OSCC. Similarly, Yiyang Liu and his team also found that decreased miR-134 was observed in renal cell carcinoma (RCC) specimens and cells [30]. Wang et al. suggested that miR-134 was a tumor suppressor in various cancers, such as prostate cancer [31]. Furthermore, miR-134 has been reported to be dysregulated in OSCC and correlated with clinical pathology [32]. Besides, a previous study found that the upregulated miR-134 could suppress proliferation in gastric cancer cells and tumor formation in xenograft tumor assays, therefore conferring the antioncogenic role of miR-134 [33]. Moreover, a prior study also found that miR-134 was significantly inhibited in tumor and metastasis tissues and that ectopic overexpression of miR-134 significantly suppressed cell proliferation, colony formation and migration and invasion in tumor cells [34].
In the study, we observed a significant increase in P21, P27, GSK3 and p-GSK3 expression, and a reduction LAMC2, PI3K, Akt and the extent of Akt phosphorylation expression when tumor stem cells were treated with miR-134 mimics. When LAMC2 expression was decreased by siRNA, OSCC stem cell proliferation, migration and invasiveness were inhibited and apoptosis were promoted. Accumulating evidences revealed that miR-134 is found associated with oral cancer progression, including OSCC, and was altered in OSCC [9, 32, 35]. In addition, we found that LAMC2 is a putative target gene of miR-143 according to the evidence in the online analysis software www.microrna.org. LAMC2, located on chromosome 1q25-q31, is a subunit of the heterotrimeric glycoprotein laminin-332, which consists of the 3, 3, and 2 chains [10]. LAMC2 was upregulated in OSCC, and the propagation of LAMC2-positive cells was concerned with cancer invasion [11]. LAMC2 promotes invasion, migration and epithelial-mesenchymal transition (EMT) of lung cells [36]. Furthermore, the inhibition of Akt signaling pathway upregulates LAMC2 protein level, while high LAMC2 inhibits Akt signaling pathway [15]. Moreover, Yuan et al. have illuminated that PI3K is associated with malignant behavior including cell growth, survival, and migration [37]. It has been reported that activated PI3K-Akt signaling pathway inhibited cell death through direct inhibition of pro-apoptotic mediators in OSCC [12]. PI3K-Akt pathway has been showed to be altered in many cancers, and its activation is responsible for decreased apoptosis, and increased cell survival and drug resistance [38]. Besides that, we found that miR-134 promoted cell apoptosis through its indirect effects on anti-apoptotic gene Bcl-2 and pro-apoptotic genes, such as Bax and caspase-9. Consistent with our study, a prior study revealed that overexpressed miR-134 decreased Bcl-2 expression [39]. The results of RT-qPCR demonstrates that ectopic expression of miR-134 increases the mRNA levels of caspase-9 and Bax, and decreases the mRNA level of Bcl-2.
To conclude, our study showed that miR-134 blocks PI3K-Akt signaling pathway through the downregulation of LAMC2, thus inhibiting cell invasion, migration and metastases of tumor stem cells in OSCC (Fig. 11). We speculate that miR-134 may be a promising new direction in the development of therapeutic treatments for OSCC. However, the results of the present study are preliminary, and additional study is still required.
Footnotes
Acknowledgments
We acknowledge and appreciate our colleagues for their valuable efforts and comments on this paper. This work was supported by the National Natural Science Foundation of China (81400513), Innovative research team of high-level local universities in Shanghai, National Key R&D Program of China (2017 YFC0840100, 2017YFC0840110), and Shanghai’s top priority clinical medicine center (2017ZZ01011).
Conflict of interest
None.
References
1.
YamamotoK.KawaguchiM.ShimomuraT.IzumiA.KonariK.HondaA.LinC.Y.JohnsonM.D.YamashitaY.FukushimaT. and KataokaH., Hepatocyte growth factor activator inhibitor type-2 (HAI-2)/SPINT2 contributes to invasive growth of oral squamous cell carcinoma cells, Oncotarget9 (2018), 11691–11706.
2.
H.S., Apoptosis-inducing activity and tumor-specificity of antitumor agents against oral squamous cell carcinoma, Japanese Dental Science Review46 (2010), 173–187.
3.
LiS.ChaJ.KimJ.KimK.Y.KimH.J.NamW. and ChaI.H., Insulin-like growth factor II mRNA-binding protein 3: a novel prognostic biomarker for oral squamous cell carcinoma, Head Neck33 (2011), 368–374.
4.
BorgesG.A.RegoD.F.AssadD.X.ColettaR.D.De Luca CantoG. and GuerraE.N., In vivo and in vitro effects of curcumin on head and neck carcinoma: a systematic review, J Oral Pathol Med46 (2017), 3–20.
5.
WangJ.LiL.GaoL.GuanC.SuK.LiL.LuoW.ChenH. and JiP., Identification of differentially expressed genes in oral squamous cell carcinoma TCA8113 cells, Oncol Lett14 (2017), 7055–7068.
6.
WangT.RenY.LiuR.MaJ.ShiY.ZhangL. and BuR., miR-195-5p suppresses the proliferation, migration, and invasion of oral squamous cell carcinoma by targeting TRIM14, Biomed Res Int2017 (2017), 7378148.
7.
KitamuraK.SeikeM.OkanoT.MatsudaK.MiyanagaA.MizutaniH.NoroR.MinegishiY.KubotaK. and GemmaA., MiR-134/487b/655 cluster regulates TGF-beta-induced epithelial-mesenchymal transition and drug resistance to gefitinib by targeting MAGI2 in lung adenocarcinoma cells, Mol Cancer Ther13 (2014), 444–453.
8.
ZhangX.WangH.ZhangS.SongJ.ZhangY.WeiX. and FengZ., MiR-134 functions as a regulator of cell proliferation, apoptosis, and migration involving lung septation, In Vitro Cell Dev Biol Anim48 (2012), 131–136.
9.
LiuC.J.ShenW.G.PengS.Y.ChengH.W.KaoS.Y.LinS.C. and ChangK.W., miR-134 induces oncogenicity and metastasis in head and neck carcinoma through targeting WWOX gene, Int J Cancer134 (2014), 811–821.
10.
DingJ.YangC. and YangS., LINC00511 interacts with miR-765 and modulates tongue squamous cell carcinoma progression by targeting LAMC2, J Oral Pathol Med47 (2018), 468–476.
11.
NguyenC.T.OkamuraT.MoritaK.I.YamaguchiS.HaradaH.MikiY.IzumoT.KayamoriK.YamaguchiA. and SakamotoK., LAMC2 is a predictive marker for the malignant progression of leukoplakia, J Oral Pathol Med46 (2017), 223–231.
12.
YangJ.RenX.ZhangL.LiY.ChengB. and XiaJ., Oridonin inhibits oral cancer growth and PI3K/Akt signaling pathway, Biomed Pharmacother100 (2018), 226–232.
13.
MartiniM.CiraoloE.GulluniF. and HirschE., Targeting PI3K in cancer: any good news? Front Oncol3 (2013), 108.
14.
LiP.XiaoL.Y. and TanH., Muc-1 promotes migration and invasion of oral squamous cell carcinoma cells via PI3K-Akt signaling, Int J Clin Exp Pathol8 (2015), 10365–10374.
15.
RaoG.PierobonM.KimI.K.HsuW.H.DengJ.MoonY.W.PetricoinE.F.ZhangY.W.WangY. and GiacconeG., Inhibition of AKT1 signaling promotes invasion and metastasis of non-small cell lung cancer cells with K-RAS or EGFR mutations, Sci Rep7 (2017), 7066.
16.
ChiouS.H.YuC.C.HuangC.Y.LinS.C.LiuC.J.TsaiT.H.ChouS.H.ChienC.S.KuH.H. and LoJ.F., Positive correlations of Oct-4 and nanog in oral cancer stem-like cells and high-grade oral squamous cell carcinoma, Clin Cancer Res14 (2008), 4085–4095.
17.
HungK.F.LiaoP.C.ChenC.K.ChiuY.T.ChengD.H.KawasumiM.KaoS.Y. and LoJ.F., ASB6 promotes the stemness properties and sustains metastatic potential of oral squamous cell carcinoma cells by attenuating ER stress, Int J Biol Sci15 (2019), 1080–1090.
18.
TuoY.L.LiX.M. and LuoJ., Long noncoding RNA UCA1 modulates breast cancer cell growth and apoptosis through decreasing tumor suppressive miR-143, Eur Rev Med Pharmacol Sci19 (2015), 3403–3411.
19.
KimC.J.TeradoT.TambeY.MukaishoK.I.SugiharaH.KawauchiA. and InoueH., Anti-oncogenic activities of cyclin D1b siRNA on human bladder cancer cells via induction of apoptosis and suppression of cancer cell stemness and invasiveness, Int J Oncol52 (2018), 231–240.
20.
LuX.Z.ZouY.G.YinX.M.ChenW.T. and ZhangC.P., Expression of MMP1 mRNA in oral squamous cell carcinoma and paired normal tissues, Nan Fang Yi Ke Da Xue Xue Bao28 (2008), 1362–1364.
21.
SapkotaD.BrulandO.CosteaD.E.HaugenH.VasstrandE.N. and IbrahimS.O., S100A14 regulates the invasive potential of oral squamous cell carcinoma derived cell-lines in vitro by modulating expression of matrix metalloproteinases, MMP1 and MMP9, Eur J Cancer47 (2011), 600–610.
22.
MingX.Y.ZhangX.CaoT.T.ZhangL.Y.QiJ.L.KamN.W.TangX.M.CuiY.Z.ZhangB.Z.LiY.QinY.R. and GuanX.Y., RHCG suppresses tumorigenicity and metastasis in esophageal squamous cell carcinoma via inhibiting NF-kappaB signaling and MMP1 expression, Theranostics8 (2018), 185–198.
23.
ShenT.C.ChangW.S.TsaiC.W.ChaoC.Y.LinY.T.HsiaoC.L.HsuC.L.ChenW.C.HsiaT.C. and BauD.T., The contribution of matrix metalloproteinase-1 promoter genotypes in taiwan lung cancer risk, Anticancer Res38 (2018), 253–257.
24.
GaoP.LiC.ChangZ.WangX. and XuanM., Carcinoma associated fibroblasts derived from oral squamous cell carcinoma promote lymphangiogenesis via c-Met/PI3K/AKT in vitro, Oncol Lett15 (2018), 331–337.
25.
QiuY.LiC.WangQ.ZengX. and JiP., Tanshinone IIA induces cell death via Beclin-1-dependent autophagy in oral squamous cell carcinoma SCC-9 cell line, Cancer Med7 (2018), 397–407.
26.
TangY.QiaoG.XuE.XuanY.LiaoM. and YinG., Biomarkers for early diagnosis, prognosis, prediction, and recurrence monitoring of non-small cell lung cancer, Onco Targets Ther10 (2017), 4527–4534.
27.
KongD.ZhangG.MaH. and JiangG., miR-1271 inhibits OSCC cell growth and metastasis by targeting ALK, Neoplasma62 (2015), 559–566.
28.
CaiZ.HaoX.Y. and LiuF.X., MicroRNA-186 serves as a tumor suppressor in oral squamous cell carcinoma by negatively regulating the protein tyrosine phosphatase SHP2 expression, Arch Oral Biol89 (2018), 20–25.
29.
WuB.LeiD.WangL.YangX.JiaS.YangZ.ShanC.YangX.ZhangC. and LuB., MiRNA-101 inhibits oral squamous-cell carcinoma growth and metastasis by targeting zinc finger E-box binding homeobox 1, Am J Cancer Res6 (2016), 1396–1407.
30.
LiuY.ZhangM.QianJ.BaoM.MengX.ZhangS.ZhangL.ZhaoR.LiS.CaoQ.LiP.JuX.LuQ.LiJ.ShaoP.QinC. and YinC., miR-134 functions as a tumor suppressor in cell proliferation and epithelial-to-mesenchymal Transition by targeting KRAS in renal cell carcinoma cells, DNA Cell Biol34 (2015), 429–436.
31.
WangW.L.ChatterjeeN.ChitturS.V.WelshJ. and TenniswoodM.P., Effects of 1alpha, 25 dihydroxyvitamin D3 and testosterone on miRNA and mRNA expression in LNCaP cells, Mol Cancer10 (2011), 58.
32.
XuR.ZengG.GaoJ.RenY.ZhangZ.ZhangQ.ZhaoJ.TaoH. and LiD., miR-138 suppresses the proliferation of oral squamous cell carcinoma cells by targeting Yes-associated protein 1, Oncol Rep34 (2015), 2171–2178.
33.
LiuY.SunY. and ZhaoA., MicroRNA-134 suppresses cell proliferation in gastric cancer cells via targeting of GOLPH3, Oncol Rep37 (2017), 2441–2448.
34.
El-DalyS.M.AbbaM.L.PatilN. and AllgayerH., miRs-134 and -370 function as tumor suppressors in colorectal cancer by independently suppressing EGFR and PI3K signalling, Sci Rep6 (2016), 24720.
35.
ShiZ.JohnsonJ.J.JiangR.LiuY. and StackM.S., Decrease of miR-146a is associated with the aggressiveness of human oral squamous cell carcinoma, Arch Oral Biol60 (2015), 1416–1427.
36.
MoonY.W.RaoG.KimJ.J.ShimH.S.ParkK.S.AnS.S.KimB.SteegP.S.SarfarazS.Changwoo LeeL.VoellerD.ChoiE.Y.LuoJ.PalmieriD.ChungH.C.KimJ.H.WangY. and GiacconeG., LAMC2 enhances the metastatic potential of lung adenocarcinoma, Cell Death Differ22 (2015), 1341–1352.
37.
YuanT.L. and CantleyL.C., PI3K pathway alterations in cancer: variations on a theme, Oncogene27 (2008), 5497–5510.
38.
SmolenskyD.RathoreK.BournJ. and CekanovaM., Inhibition of the PI3K/AKT pathway sensitizes oral squamous cell carcinoma cells to anthracycline-based chemotherapy in vitro, J Cell Biochem118 (2017), 2615–2624.
39.
HuangW.LiuX.CaoJ.MengF.LiM.ChenB. and ZhangJ., miR-134 regulates ischemia/reperfusion injury-induced neuronal cell death by regulating CREB signaling, J Mol Neurosci55 (2015), 821–829.