Gastric cancer is one of the leading causes of death worldwide. MicroRNA-30a (miR-30a) has been demonstrated to be involved in several types of cancer development.
OBJECTIVE:
We aimed to identify the molecular mechanism of miR-30a in gastric cancer.
METHODS:
We investigated the expression of miR-30a in gastric cancer tissues by qRT-PCR. The role of miR-30a on the metastasis and proliferation of gastric cancer was evaluated by cell migration assay, CCK-8 assay and tumor peritoneal dissemination model. The target of miR-30a in gastric cancer was identified.
RESULTS:
We discovered that miR-30a was significantly downregulated in gastric cancer tissues compared with adjacent nonmalignant tissues. The expression of miR-30a was inversely correlated with progression of gastric cancer. Gain- and loss-of function revealed that miR-30a acted as a potent tumor suppressor in gastric cancer. Re-expressed miR-30a inhibited gastric cancer cells migration, knock down miR-30a have the opposite effects. Furthermore, overexpression of miR-30a suppressed tumor peritoneal dissemination in vivo. We identified that fibroblast activation protein (FAP) was a direct target of miR-30a. The relative expression of FAP was significantly higher in gastric cancer tissues compared with adjacent nonmalignant tissues. Inhibition of FAP could recapitulate the effects of miR-30a, and overexpression of FAP could abrogate the effect of miR-30a.
CONCLUSION:
MiR-30a inhibited gastric cancer metastasis by targeting FAP, suggesting that miR-30a may function as a novel tumor suppressor in gastric cancer.
Gastric cancer is one of the commonly diagnosed and leading causes of cancer death in China. Incidence and mortality of gastric cancer in 2015 are estimated as 679,100 cases and 498,000 cases, respectively, according to the data from 72 population-based cancer registries in China from 2009–2011 [1]. Based on the early diagnose and rapid development of therapeutic methods, the incidence and mortality of gastric cancer were slowly down in the last decade in both male and female Chinese [1]. However, the outcomes of patients, especially with metastatic disease are still poor. So far there have no high quality data to verify whether surgery benefits to patients with metastatic disease or not [2], which indicates that further discovery of the molecular mechanisms involved in the development and progression of gastric cancer is essential.
MicroRNAs (miRNAs) have been identified as important post-transcriptional regulators of gene expression in various diseases including malignancies. Ueda et al. proved that the expression of unique miRNAs was associated with progression and prognosis of gastric cancer [3]. Our and other’s groups have identified a series of miRNAs (including miR-25, miR-141, miR-22 miR-29c, and miR-106b-25 cluster et al) were involved in regulating the growth, migration and invasion of gastric cancer [4, 5, 6, 7, 8, 9]. In our previous study, we discovered that miR-30b, which belongs to miR-30 family, was able to suppress gastric cancer growth and regulate the apoptosis of cancer cells though targeting plasminogen activator inhibitor-1 [10]. It prompted us to investigate the roles of other members of miR-30 family in gastric cancer. We analyzed the expression of five members of miR-30 family in gastric cancer from TCGA database and found that miR-30a had the greatest change in gastric cancer tissues compared to adjacent tissues. Recently, miR-30a has been identified as a tumor suppressor in several types of carcinoma, such as colorectal cancer [11, 12, 13], breast cancer [14] and non-small cell lung cancer (NSCLC) [15, 16, 17]. In gastric cancer, miR-30a was identified as a potent tumor suppressor by inhibiting intestinal metaplasia of epithelial cells and epithelial-to-mesenchymal transition (EMT) of cancer cells [18, 19]. However, the detailed roles and underlying mechanisms of miR-30a in the progression of gastric cancer need to be further studied. Therefore we tried to investigate other possible mechanisms of miR-30a during gastric cancer development.
Fibroblast activation protein (FAP) is a homodimeric integral membrane gelatinase belonging to the serine protease family and mainly expressed in reactive human tumor stromal fibroblasts [20] and several epithelial-derived tumor cells including gastric cancer [21, 22, 23, 24, 25, 26]. In gastric cancer, high FAP expression in cancer tissues was associated with Lauren classification, the degree of differentiation, depth of tumor invasion and TNM (T: size of the primary tumor, N: degree of spread to regional lymph nodes, M: presence of distant metastasis) stages [27]. FAP-positive cancer associated fibroblasts (CAFs) promoted MGC-803 cell migration and invasion [27, 28]. However, there is little known about the function of FAP in gastric cancer cells. In the present study, we further identified FAP was upregulated in gastric cancer tissues and was directly targeted by miR-30a. Taken together, miR-30a acted as a potent tumor suppressor in gastric cancer by targeting FAP, resulting in decreased tumor metastasis both in vitro and in vivo. These findings suggested that miR-30a and FAP may function as regulators of gastric cancer progression and may be potential targets for gastric cancer treatment.
Materials and methods
Patient and specimens
Twenty nine pairs of frozen gastric cancer tissue samples and their adjacent nonmalignant tissues ( 6 cm laterally from the edge of tumor region) were obtained from the Southwest Hospital of Third Military Medical University (Chongqing, China). Samples were collected from patients during operation and stored in liquid nitrogen until use. Detailed patients and disease characteristics are documented in Table 1. All samples were collected with consent of patients and the experiments were approved by the Ethics Review Board at Third Military Medical University.
Association between miR-30a expression and clinicopathologic variables of patients with gastric cancers
Variables
29
Average expression of miR-30a (mean SD)
value
Age (years)
50
10
0.89 0.29
0.692
50
19
1.03 0.19
Gender
Male
20
0.94 0.19
0.661
Female
9
0.89 0.57
TNM stage
I–II
12
11.47 0.31
0.009
III–IV
17
0.64 0.11
Lymph node metastasis
Positive
22
1.05 0.18
0.525
Negative
7
0.80 0.37
Cell culture
Gastric cancer cell lines SGC-7901, BGC-823, AGS and HGC-27 were obtained from Cell Bank of Chinese Academy of Sciences (Shanghai, China). HEK293T cells were obtained from HanBio Company (Shanghai, China). Cells were cultured with RPMI 1640 (SGC-7901, AGS and HGC-27) or Dulbecco’s modified Eagle’s medium (DMEM) (BGC-823 and HEK293T) supplemented with 10% fetal calf serum individually at 37C in 5% CO.
RNA isolation and quantitative real-time PCR (qRT-PCR)
Total RNA was extracted from gastric cancer tissues or cultured cells using Trizol (Invitrogen, Calsbad, CA, USA) method. Expression of miR-30a was measured using a Taqman MicroRNA Reverse Transcription kit followed by TaqMan MicroRNA Assays (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instruction. The data were normalized to endogenous U6 snRNA. For mRNA detection, reverse transcription was performed using the PrimeScript RT Master Mix (TaKaRa, Dalian, China). Real time-PCR was performed using SYBR (Toyobo, Shanghai, China). The data were normalized to GAPDH. Primers: FAP (sense primer, 5’-AGAACCATGCTTTGGAGATACT-3’ and antisense primer, 5’-ACTGAATTGGACGAGG AAGC-3’), GAPDH (sense primer, 5’-CCACTCCTCC ACCTTTGAC-3’ and antisense primer, 5’-ACCCTGT TGCTGTAGCCA-3’ All the qRT-PCR reactions used Bio-Rad CFX96 real-time PCR system. The results were calculated by 2 method.
Cell migration assay
In vitro cell migration ability was measured using transwell chambers (5 m pore size, Costar, Cambridge, MA, USA). Briefly, SGC-7901 and BGC-823 cells transfected with miR-30a mimics (50 nM) or inhibitors (100 nM) were suspended in serum-free RPMI 1640 or DMEM and seeded onto up chamber (1 10 cells per chamber). In the lower chamber, 300 l RPMI 1640 or DMEM with 10% fetal calf serum was added as chemoattractant. 24 hours later, cells in upper chamber were gently wiped off with a cotton swab. Cells located on the lower surface of chamber were fixed with 4% paraformaldehyde and then stained with 0.1% crystal violet, the extra crystal violet was washed away with phosphate buffer solution (PBS). Then cells were photographed under a microscope in five randomly selected fields ( 100). Then crystal violet adhering to cells was re-dissolved by 30% acetic acid and the absorbance values were detected under 570 nm on a microtiter plate reader (Molecular Devices, San Jose, CA, USA) according to the manufacturer’s protocol. The mean values of triplicate assays were used for statistical analysis. All assays were independently repeated three times.
Cell proliferation assay
To detect effect of miR-30a on cell proliferation, SGC-7901 and BGC-823 cells were transfected with miR-30a mimics, mimic negative controls (NCs, 50 nM) or miR-30a inhibitors, inhibitor NCs (100 nM) (RioBio, Guangzhou, China) respectively. 24 hours after transfection, cells were plated in 96-well plates (1 10 cells per well). 24 hours later, the medium was removed. 100 l serum-free medium plus 10 l CCK8 (Dojindo Laboratories, Kumamoto, Japan) was added to each well and incubated at 37C for 1 hour. Then the absorbance values were detected under 450 nm on a microtiter plate reader (Molecular Devices, San Jose, CA, USA) according to the manufacturer’s protocol.
Cell transfection
MiR-30a mimics, inhibitors, agomirs, FAP siRNAs and respectively corresponding negative controls used in this study were all obtained from Riobio Company (Guangzhou, China). FAP overexpression vector pCMV-FAP (RC204692) and pCMV-MOCK vector (PS100001) were purchased from OriGene (Rockville, MD, USA). Cells were transfected with Lipofectamine 2000 (Invitrogen, Calsbad, CA, USA) according to the manufacturer’s instructions. 24 hours after transfection, cells were used for functional analysis in terms of migration and proliferation. 48 hours after transfection, cells were harvested and used for western blot analysis.
Luciferase assays
A wild type or a mutant 3’UTR of human FAP were inserted into a dual luciferase miRNA expression reporter vector, pMIR-REPORT (Applied Biosystems, Applied Biosystems, Foster City, CA, USA). The mutant vector was constructed by replacing miR-30a binding sites in the 3’UTR of FAP. HEK293T cells were seeded onto 96-well plates (1 10 cells per well) and co-transfected with wild type or mutant vectors (200 ng per well), pRL-TK Renilla luciferase reports (10 ng per well), miR-30a mimics or mimic-NC (50 nM). 24 hours later, the luciferase activities were measured using the Dual Luciferase Reporter Assay (Promega, Madison, MI, USA) on GLOMAX 20/20 luminometer (Promega, Madison, MI, USA). The firefly luciferase activity was normalized to the renilla luciferase activity. All assays were independently repeated three times and in quadruplicate.
In vivo tumor peritoneal dissemination assay
Female BABL/c nude mice (6–8 weeks old) were purchased from Beijing HFK Bioscience Limited Company (Beijing, China). The peritoneal dissemination assay was performed as described before [5]. Briefly, SGC-7901 cells were transfected with agomir-NC or agomir-30a (200 nM) for 24 hours and then collected and resuspended in PBS 1 10SGC-7901 cells in 100 L PBS were injected into the abdominal cavity of each nude mouse (five per group). Six weeks later, the mice were sacrificed and the macroscopic nodules in the abdominal cavity were counted. The animal studies were approved by the Institutional Animal Care and Use Committee of Third Military Medical University.
Western blot assay
Western blot was performed as described before [29] with rabbit polyclonal FAP (1:1000; Abcam, Cambridge, UK). Monoclonal GAPDH antibody (1:5000; Cell Signaling Technology) was used as an internal reference. HRP-conjugated anti-rabbit or anti-mouse immunoglobulin-G antibody was used as the secondary antibody (1:10000; Zhong Shan, Shanghai, China). Signals were detected using the SuperSignal West Dura Extended Duration Substrate Kit (Thermo Scientific Pierce, Rockford, IL, USA). The results were analysed using Image J.
Statistical analysis
The student’s t test was performed to analyze whe-ther two experimental groups have significant difference. Paired or unpaired t-test was used to determine the differences between paired or unpaired samples, respectively. 0.05 was considered statistically significant. All data were analyzed using GraphPad Prism software version 5.0 (Graphpad Software Inc., La Jolla, CA, USA) and expressed as mean standard deviation (SD).
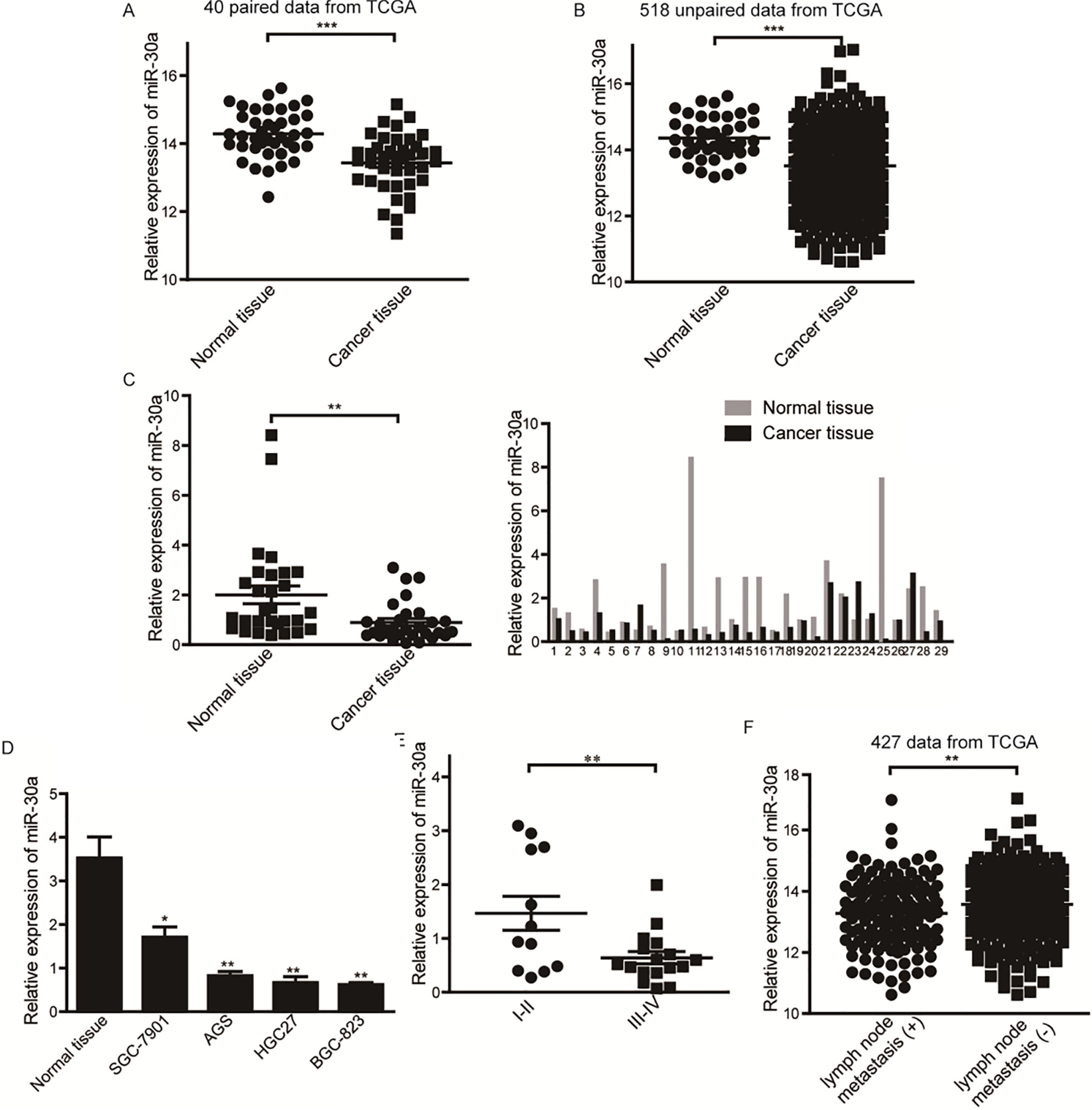
Expression of miR-30a in gastric cancer tissues and the relationship between miR-30a level and clinical characters. (A) Expression of miR-30a in 40 paired gastric cancer tissues and matched nonmalignant tissues from TCGA. (B) Expression of miR-30a in 518 unpaired gastric cancer tissues and matched nonmalignant tissues from TCGA (42 normal vs 476 cancerous). (C) Relative expression of miR-30a in 29 pairs of gastric cancer tissues and matched nonmalignant tissues quantified by qRT-PCR. U6 was used as an internal inference. (D) Expression of miR-30a in nonmalignant tissues and gastric cancer cell lines. All assays were performed in triplicates. (E) Correlation of miR-30a expression with TNM stages (I II vs III IV). (F) Expression of miR-30a in gastric cancer tissues with or without lymph node metastasis from TCGA. 0.05, 0.01, 0.0001.
Results
Downregulation of miR-30a in gastric cancer tissues is correlated with gastric cancer progression and lymph node metastasis
We firstly analyzed the expression of five members of miR-30 family in gastric cancer from TCGA database. We found that miR-30a had the greatest downregulation in 40 paired and 518 unpaired cases (Fig. 1A and B).We then verified the dysregulation of miR-30a in 29 pairs of gastric adenocarcinoma tissues and the matched adjacent nonmalignant tissues by qRT-PCR. The results showed that the expression of miR-30a was significantly decreased in tumor tissues (Fig. 1C). The expression of miR-30a in gastric cancer cell lines was also decreased compared with nonmalignant gastric tissues (Fig. 1D). We further investigated the relationship between miR-30a levels and clinicopathological characteristics. We found that the expression of miR-30a was inversely correlated with TNM stages of gastric cancer (Table 1). Lower expression of miR-30a was significantly associated with a more aggressive tumor stage (Fig. 1E, stage I II vs III IV). Considering the limitation of relatively small cohort size, we next analyzed a larger scale clinical data (427 cases) from TCGA and found that the expression of miR-30a was inversely correlated with lymph node metastasis in gastric cancer tissues (Fig. 1F). These results suggest that miR-30a is consistently downregulated in gastric cancer and the expression of miR-30a is inversely correlated with gastric cancer progression and lymph node metastasis.
MiR-30a prevents gastric cancer cells migration in vitro
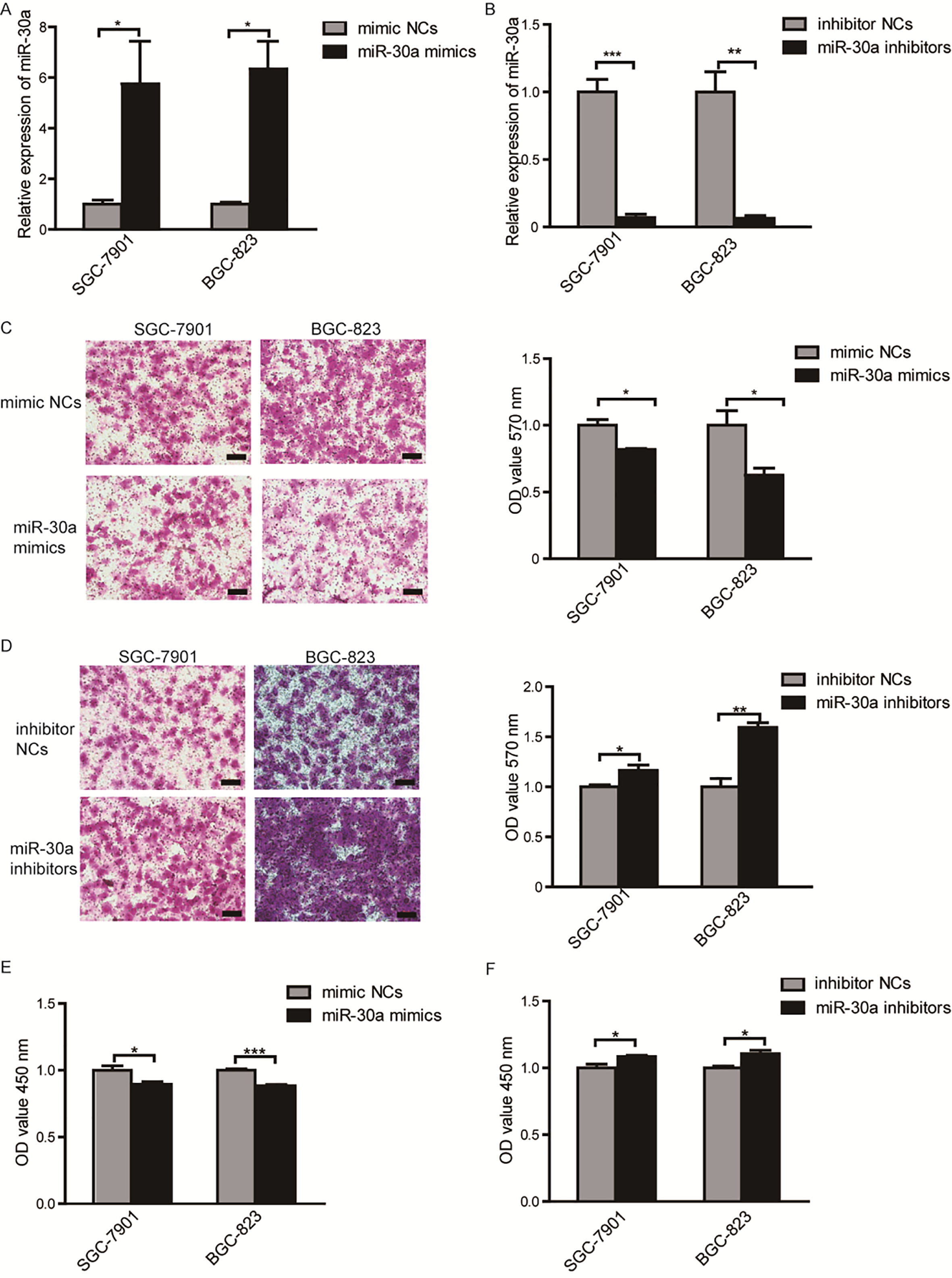
To investigate the function of miR-30a in gastric cancer, SGC-7901 and BGC-823 cell lines were transfected with miR-30a mimics (50 nM), mimic-NCs (50 nM) or miR-30a inhibitors (100 nM), inhibitor NCs (100 nM). Overexpression or inhibition of miR-30a was confirmed using qRT-PCR as shown in Fig. 2A and B. To determine the role of miR-30a on cellular migration, transwell assays were conducted and the results showed that overexpression of miR-30a significantly reduced the migration abilities of both SGC-7901 and BGC-823 cells (Fig. 2C), and inhibition of miR-30a promoted cells migration (Fig. 2D). We also evaluate the effects of miR-30a on cells proliferation by CCK8 assays. As shown in Fig. 2E, overexpression of miR-30a significantly reduced cells proliferation at 24 hours after transfection, and downregulation of miR-30a promoted cells proliferation (Fig. 2F). However, the effect of miR-30a mimics or inhibitor on cell proliferation was not strong enough ( 15% change extent). These results suggest that miR-30a could effectively prevent migration of gastric cancer cells and slightly regulate proliferation of cells in vitro.
MiR-30a is capable of inhibiting migration and proliferation of SGC-7901 and BGC-823 cells. (A) The expression of miR-30a after transfection with miR-30a mimics or inhibitors (B) in SGC-7901 and BGC-823 cells were determined by qRT-PCR. (C) Migration of cells transfected with miR-30a mimics or inhibitors (D) was determined by transwell assays. Representative photomicrographs of transwell assays were taken under 100 magnification (left) and the absorbance of eluted crystal violet was measured at 570 nm (right). Scale bar 50 m. 3 (E) Proliferation of cells transfected with miR-30a mimics or inhibitors (F) was detected using CCK8 assays. 5. All of the experiments were performed at least for three independent times. 0.05, 0.01, 0.001.
MiR-30a directly targets FAP in gastric cancer cells
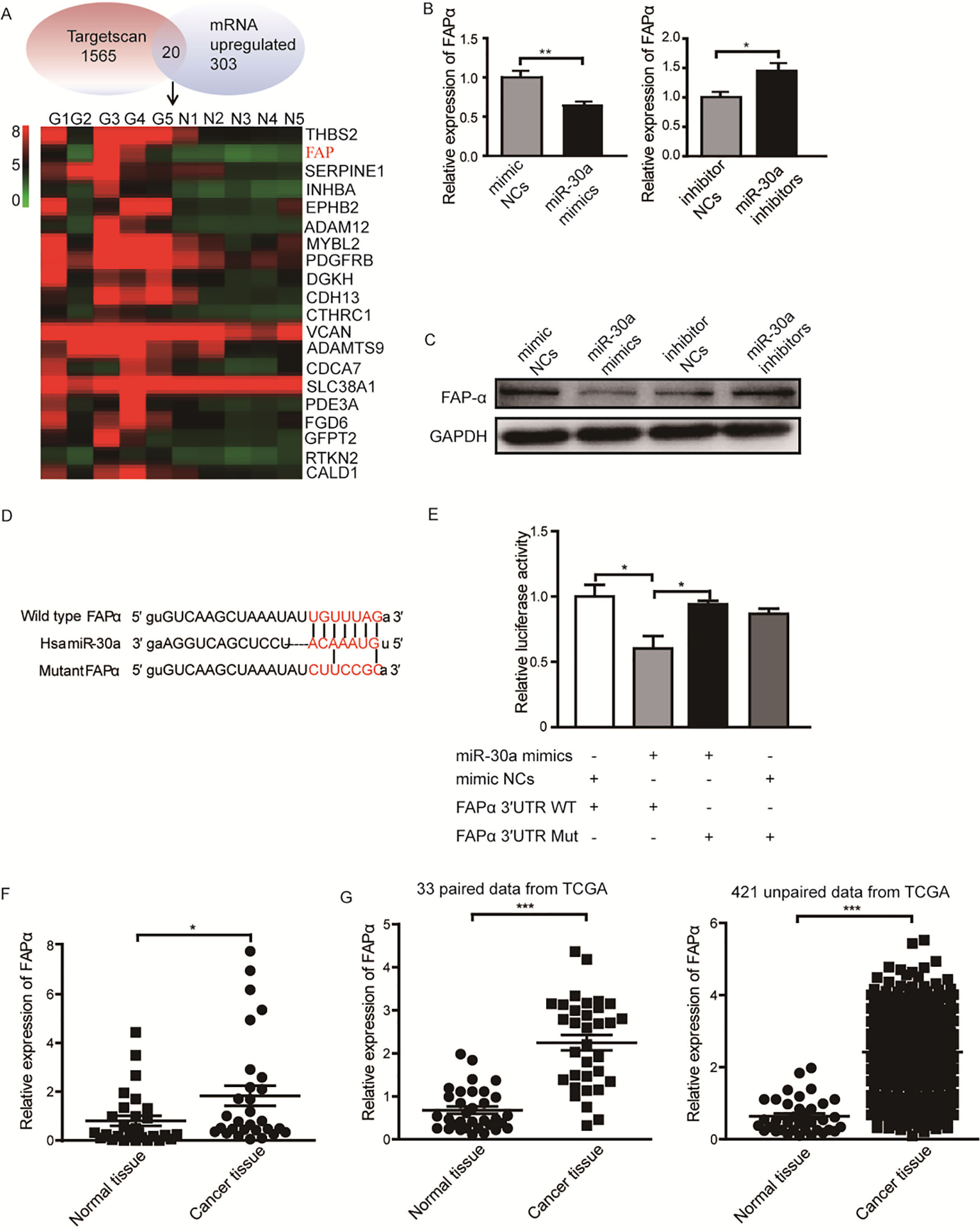
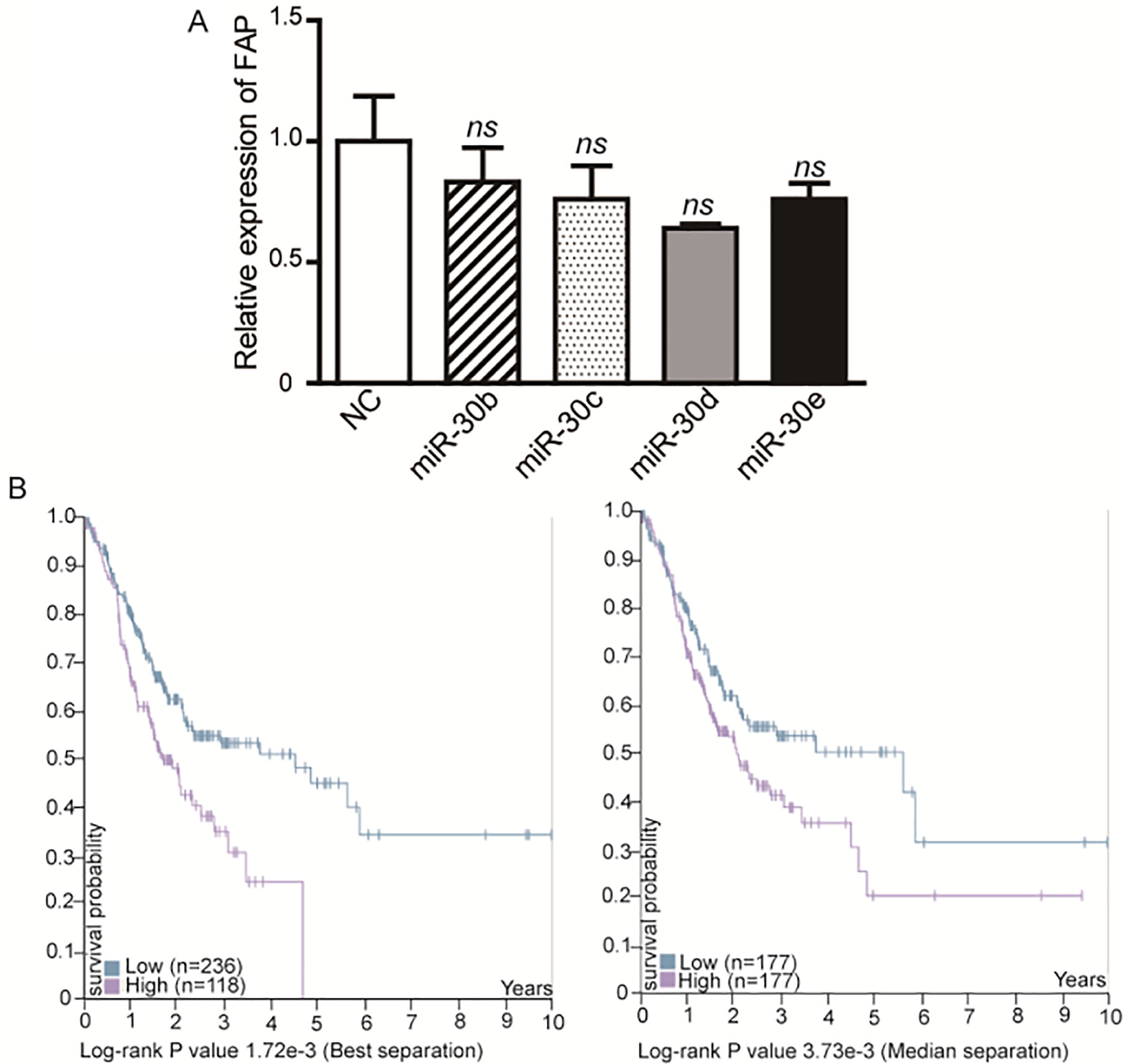
We next explored the molecular mechanism of miR-30a in gastric cancer. In our previous study, we identified the mRNA expression profile in five paired gastric cancer tissues and nonmalignant tissues using microarray [5]. In consideration of the downregulation of miR-30a in gastric cancer tissues, we focused on the upregulated genes (fold change 2, 0.05) in cancer tissues. Meanwhile, we predicted the potential targets of miR-30a using TargetScan. Combined the gene lists from TargetScan and microassay, we picked 20 genes as the candidate targets of miR-30a in gastric cancer (Fig. 3A). Based on the tumor-suppressor function of miR-30a, we focused on potential target genes which function as oncogenes. After screening by qRT-PCR and western blot, FAP was found to be inhibited by miR-30a-overexpression on both mRNA (Fig. 3B) and protein levels (Fig. 3C). We analyzed the 3’UTRs sequence of FAP and found only one binding site matched to the seed sequence of miR-30a. We next constructed luciferase reporters contained wild type 3’UTRs of FAP (wild-type vector) or mutant miR-30a binding site in the 3’UTR (mutant vector) (Fig. 3D). We performed dual luciferase assay to investigate whether miR-30a could directly bind to 3’UTR of FAP. The results showed that the relative luciferase activity was reduced by 39.8% in wild-type vector, but not in mutant vector (Fig. 3E). To rule out the possibility of other miR-30 family members could regulate the expression of FAP, we then checked the effect of mimics of miR-30b, c, d, e on the expression of FAP. We found that other miR-30 family members were not able to significantly inhibit the mRNA level of FAP (Supplementary Fig. 1A). We then detected the level of FAP in 29 pairs of gastric adenocarcinoma tissues and the matched nonmalignant tissues. The results showed that the expression of FAP was upregulated in cancer tissues (Fig. 3F). A larger scale of data from TCGA presented the same conclusion that FAP was significantly upregulated in both paired and unpaired gastric cancer tissues (Fig. 3G), and the higher expression of FAP was associated with poor overall survival of patients (Supplementary Fig. 1B). Overall, these results reveal that FAP is a direct target of miR-30a in gastric cancer cells.
Prediction and validation of miR-30a target gene in gastric cancer cells. (A) The heat map shows 20 candidate genes which have miR-30a binding site and are upregulated in gastric cancer tissues. (B) The relative mRNA and protein (C) level of FAP in SGC-7901 cells transfected with miR-30a mimics or mimic NCs were determined by qRT-PCR and western blot. GAPDH was used as internal reference. (D) Constructs of wild type vector (top) and mutant vector (bottom). (E) Luciferase activity in HEK293T cells transfected with wild type or mutant luciferase vectors, and either miR-30a mimics or mimic NCs. Luciferase activities were normalized to Renilla luciferase. 4. (F) The relative FAP mRNA level was detected in 29 paired gastric cancer tissues and the corresponding nonmalignant tissues. GAPDH was used as an internal reference. (G) Expression of FAP in 33 paired gastric cancer tissues and the matched nonmalignant tissues (left) and in 421 unparied tissues (right) (384 cancerous vs 37 normal) from TCGA database. 0.05, 0.01, 0.001.
FAP is potentially involved in the tumor-suppressor function of miR-30a in gastric cancer
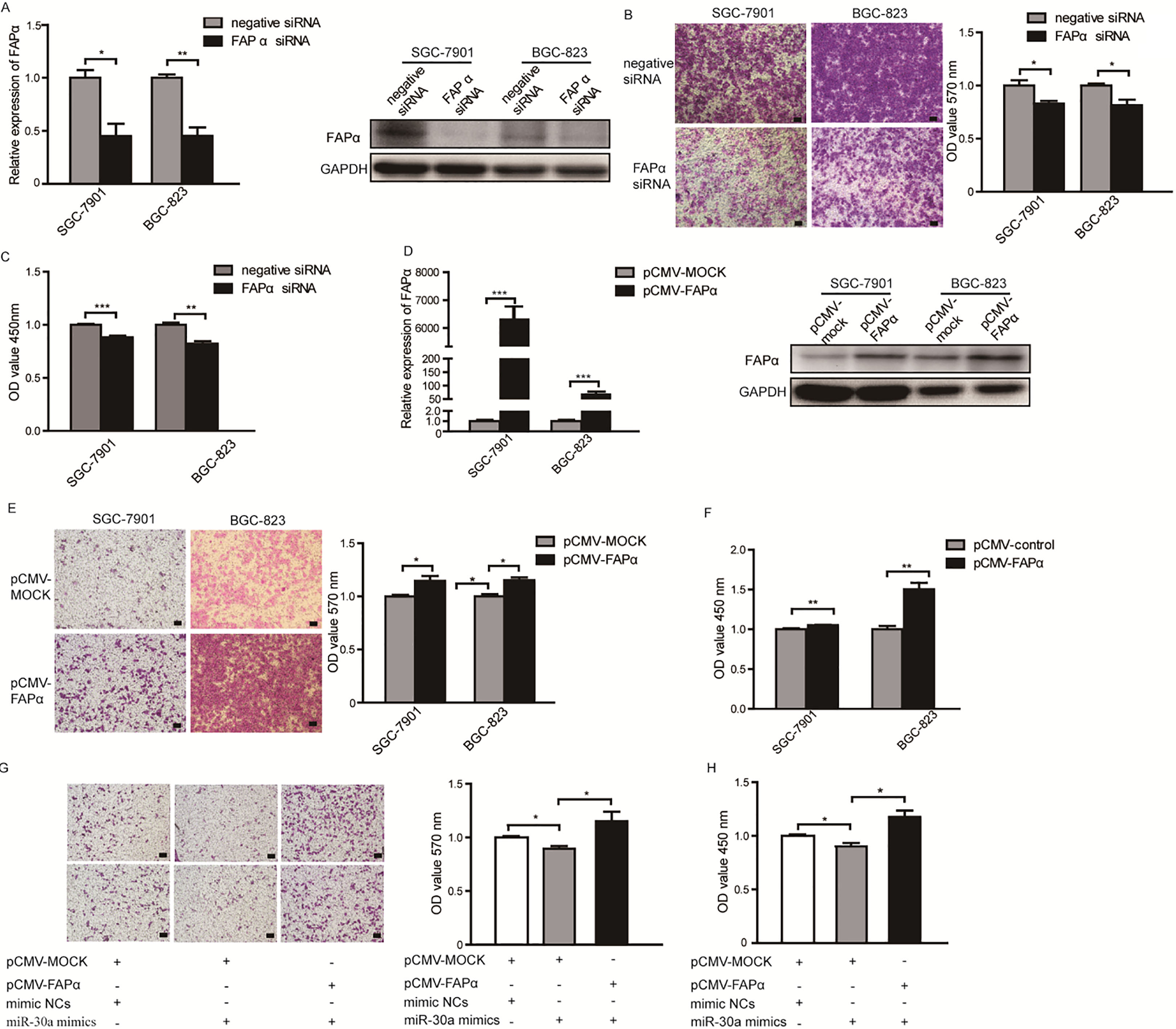
Though FAP plays pivotal roles in tumor environment, the effects of FAP in gastric cancer cells have not been elucidated yet. To investigate whether FAP mediated the suppression function of miR-30a in gastric cancer, we knocked down the expression of endogenous FAP by siRNA to mimic the effects of miR-30a overexpression. When the mRNA and protein levels of FAP were significantly reduced by siRNA in SGC-7901 and BGC-823 cells (Fig. 4A), the migration abilities of these cells were significantly inhibited (Fig. 4B). Meanwhile, inhibition of FAP caused slightly but significantly cell growth inhibition (Fig. 4C), which was comparative to the effect of miR-30a-overexpression. Next, we evaluated the effects of overexpression of FAP in SGC7901 and BGC-823 cells (Fig. 4D). The results showed that upreguation of FAP promoted cells migration (Fig. 4E) and proliferation (Fig. 4F), which were consistent with the effects of miR-30a-inhibition. Moreover, re-expression of FAP in SGC-7901 cells transfected with miR-30a mimics could restore the inhibitory function of miR-30a on cells migration and proliferation (Fig. 4G and H). These findings indicate that miR-30a inhibits migration and proliferation of gastric cancer cells by targeting FAP.
In vitro functional analysis of FAP on gastric cancer cells migration and proliferation. (A) FAP mRNA level (left) and protein level (right) in cells transfected with FAP siRNA or siRNA control. (B) Migration of cells transfected with FAP siRNA or siRNA control was measured by transwell assays. Representative photomicrographs of transwell assays were taken under 100 magnification (left) and the absorbance of eluted crystal violet was measured at 570 nm (right). Scale bar 50 m. (C) Proliferation of cells transfected with FAP siRNA or siRNA control was detected using CCK8 assays. 5. (D) FAP mRNA level (left) and protein level (right) in cells transfected with pCMV-MOCK vector or pCMV-FAP vector. (E) Migration ability of FAP-overexpressing cells was measured using transwell assays. Representative fields were taken under 100 amplification (left). Scale bar 50 m. Stained violet was redissolved and the OD value was detected at 570 nm (right). (F) Proliferation of FAP-overexpressing cells was measured with CCK8 assays. 5. (G) Migration of SGC-7901 cells transfected with miR-30a mimics or co-transfection with miR-30a mimics and pCMV vectors was measured using transwell assays. Scale bar 50 m. (H) Proliferation of SGC-7901 cells individually transfected with miR-30a mimics or co-transfection with miR-30a mimics and pCMV vectors was measured using CCK8 assays. 5. All of the experiments were performed at least for three independent times. 0.05, 0.01, 0.001.
MiR-30a inhibits peritoneal dissemination of SGC-7901-engrafted tumors in vivo
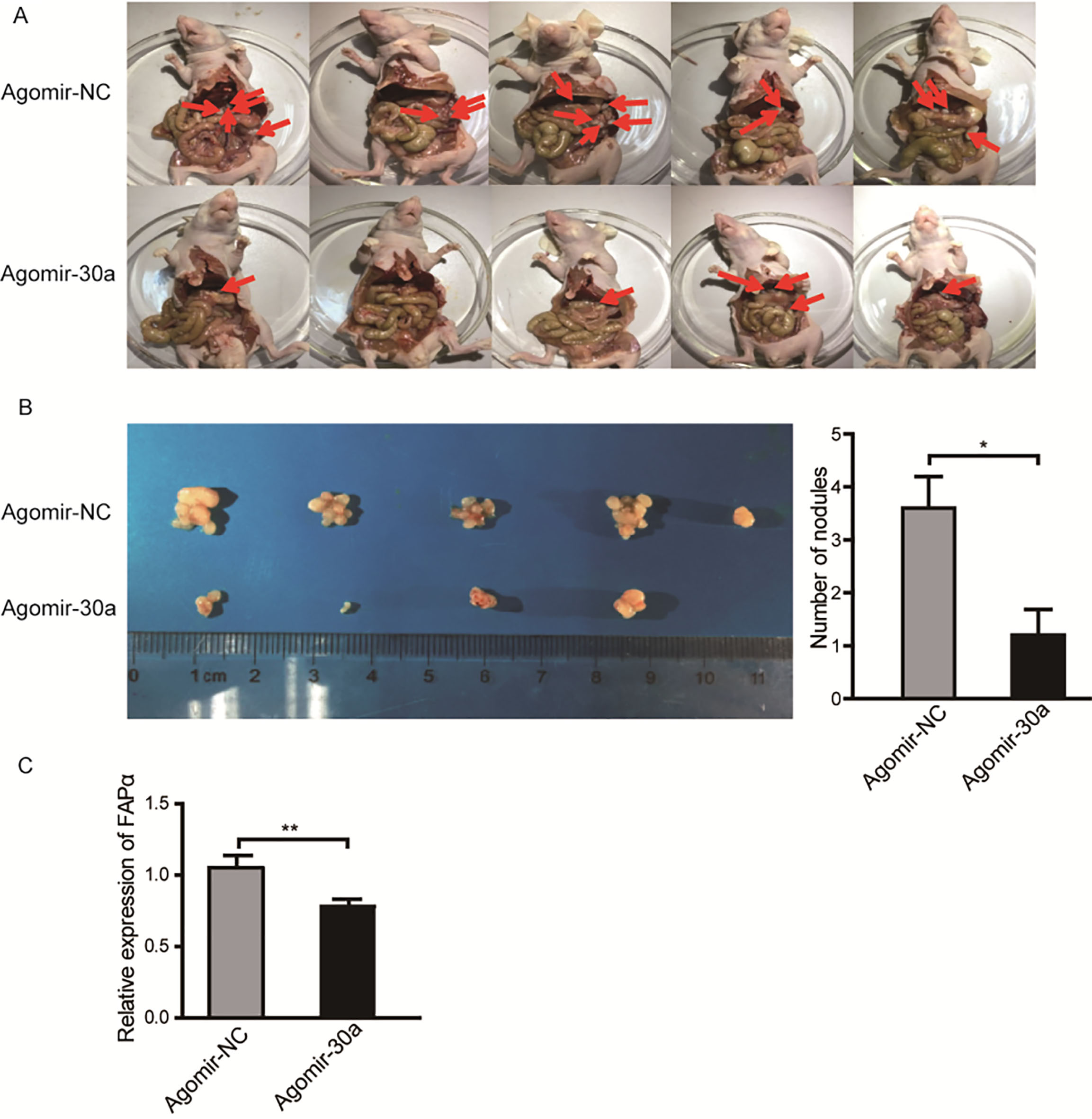
Early invasiveness to peritoneal cavity is the common character of gastric cancer. To further investigate the function of miR-30a in vivo, we selected SGC-7901 cell which has been well characterized to perform the tumor peritoneal dissemination assay via BALB/c nude mice models. The SGC-7901 cells were collected after transfection with agomir-30a (200 nM) or agomir-NC (200 nM) for 24 hours and injected into the peritoneal cavity of nude mice individually. After six weeks, mice were sacrificed and tumor nodules in peritoneal cavity were counted. The results showed that mice injected with miR-30a-overexpressing SGC-7901 cells exhibited significantly reduced number of macroscopic nodules than control group (Fig. 5A and B). The results suggest that miR-30a inhibits the peritoneal dissemination of gastric cancer in vivo.
Overexpression of miR-30a inhibits gastric cancer peritoneal dissemination in vivo. (A) Abdominal photographs of mice injected with SGC-7901 cells transfected with agomir-NC or agomir-30a. (B) Volume and number of the tumor nodules in the abdominal cavity of mice from each group. 5. 0.05.
Discussion
Currently, a series of miRNAs have been reported to be involved in regulating gastric cancer development and progression, while the exact molecular mechanisms underlying these miRNAs are still not fully understood. Usually, one miRNA can regulate numbers of target genes to play different roles during tumorigenesis. Thus, a comprehensive understanding of target genes regulatory network of miRNAs will be beneficial for all-round understanding the function of miRNAs.
In the present study, we discovered that miR-30a was significantly decreased in gastric cancer tissues and cell lines compared with adjacent nonmalignant tissues. Moreover, the expression of miR-30a was inversely correlated with gastric cancer progression and lymph node metastasis. Gain- and loss-of function revealed that miR-30a could effectively inhibit gastric cancer cell migration in vitro. miR-30a could also inhibit proliferation of gastric cancer cells but the effect was not strong. Peritoneal dissemination assay verified that miR-30a inhibited gastric cancer metastasis in vivo.
MiR-30a is a member of the miR-30 family which is versatile player in many types of cancers. Increasing evidence shows that miR-30a is a potent tumor suppressor by regulating tumor cell proliferation [11, 12, 14, 16, 30, 31], metastasis and invasion [13, 14, 17, 32], EMT [15, 17, 19, 33], autophagy [34] and chemoresistance [35] in different types of tumors.
Identifying the role and clinical application of miR-30a in gastric cancer has also attracted much attention. Liu et al. found that miR-30a was induced by runt-related transcription factor 3 (RUNX3) and targeted vimentin to regulate EMT process in gastric cancer [19]. Zou et al. found that miR-30a targeted the DNA replication protein RPA1, induced inhibition of DNA replication and DNA fragment, ultimately led to SGC-7901 cells apoptosis [31]. Liu et al. found that miR-30a inhibited the development of H. pylori-induced gastric cancer by targeting BCL9 [36]. Intestinal metaplasia is neoplastic precursor of gastric adenocarcinoma, miR-30a has been found to inhibit the development of pre-neoplastic lesions through targeting HNF4 [18]. Furthermore, Fan D’s group identified a seven-miRNA signature including miR-30a to evaluate overall survival and relapse-free survival of gastric cancer patients [37]. In addition to these, the detailed function and molecular mechanisms of miR-30a in gastric cancer have not been fully elucidated yet. In the present study, we further elaborated the tumor-suppressor function of miR-30a. Importantly, we firstly discovered that miR-30a could effectively inhibited peritoneal dissemination of gastric cancer in nude mice tumor model. Peritoneal dissemination is a devastating form of gastric cancer progression with a poor prognosis. The 5-year survival rate of patients with peritoneal dissemination is about 2%, however the molecular mechanism of peritoneal dissemination in gastric cancer remains elusive [38]. These data suggest that miR-30a is a key regulator of gastric cancer metastasis and provide new insights into the role of miR-30a in regulating peritoneal dissemination of gastric cancer.
Using bioinformatics tools and mRNA expression array of gastric cancer, we further identified FAP as a novel target of miR-30a. FAP is a homodimeric integral membrane gelatinase belonging to the serine protease family and has both dipeptidyl peptidase activity and a collagenolytic activity capable of degrading gelatin and type I collagen [20]. In tumor environment, FAP is usually expressed on CAFs and plays vital roles. For instance, Koczorowska et al. found that FAP influenced vessel sprouting and matrix stiffness by regulating the secretome of CAFs [39]. Yuan et al. found that FAP-positive CAFs promoted ovarian carcinoma metastasis through promoting angiogenesis, lymphangiogenesis and tumor cell invasion [40]. These studies attracted researchers’ attention to explore whether FAP could be a possible therapeutic target for cancer treatment. Jackson et al. found that inhibition of FAP resulted in fewer microvessels, collagen accumulation and broad apoptotic areas, ultimately suppressed human colon and lung tumor xenografts in mouse models [41]. Depletion of FAP-positive stromal cells abrogated stromal invasion and angiogenesis, reduced extracellular matrix proteins and glycosaminoglycans simultaneously and thereby diminish cancer growth [42, 43, 44]. Notably Amel S et al. identified a five-mRNA signature including FAP to predict outcome, and discriminate preinvasive from invasive digestive cancers disease [45]. Meanwhile, accumulating evidence has proved that FAP could be expressed in several epithelial-derived tumor cells [21, 22, 23, 24, 25, 26]. For instance, in ovarian carcinoma, FAP positive tumor cells indicated 66.7% tumor recurrence rates [25]. In pancreatic carcinoma, FAP was highly expressed in tumor cells and associated with desmoplasia and worse prognosis [24]. In oral squamous cell carcinoma, FAP was upregulated in cancerous tissues and promoted tumor cell proliferation and metastasis by modulating PTEN/PI3K/AKT and Ras-ERK signaling [26]. Overexpression of FAP in colorectal cancer cells was associated with lymph node metastasis [22]. These studies revealed that FAP may be involved in the development of cancer by directly regulating cancer cells.
In gastric cancer, FAP was found to be expressed in carcinoma cells and stromal cells but none in normal cells [21]. Overexpression of FAP in gastric cancer tissues was always associated with worse prognosis and poor overall survival of patients [46]. Expression of FAP in CAFs from invasion front tissues was associated with tumor Lauren classification, differentiation, depth of tumor invasion and tumor progression [28]. Co-cultured with FAP-positive CAFs, gastric cancer cells had higher proliferative, metastatic and invasive abilities [27, 28, 47]. However, the role of FAP in gastric cancer epithelial cells has not been well elucidated. Here, we discovered that FAP was upregulated in gastric cancer tissues compared with nonmalignant tissues. FAP was targeted by miR-30a but not by other members of miR-30 family. Gain- and loss-of function proved that FAP acted as a potent oncogene in gastric cancer through regulating migration and proliferation of cancer cells. Meanwhile, overexpression of FAP in gastric cancer cells could effectively abrogate tumor-suppressor function of miR-30a, which indicated that FAP was involved in miR-30a-mediated suppression of tumor growth and metastasis.
In summary, the present study concluded that miR-30a acted as an important tumor suppressor through regulating FAP in gastric cancer development. Overexpression of miR-30a inhibited tumor metastasis both in vitro and in vivo. These findings provide a better understanding of molecular mechanisms of miR-30a in gastric cancer, and indicate that miR-30a and FAP may be new promising therapeutic targets for gastric cancer.
Footnotes
Acknowledgments
This study was supported by National Natural Science Foundation of China (81872392), Chongqing Youth Science and Technology Talent Training Project (cstc2014kjrc-qnrc10008), and Scientific Research Foundation for Introduced High-level Personnel In Chongqing Medical University (41021300160256).
Conflict of interest
All authors declare no conflicts of interest.
Supplementary data
The effects of miR-30 family except for miR-30a on the expression of FAP and survival analysis of FAP in gastric cancer. (A) The mRNA level of FAP in SGC-7901 cells transfected with miR-30b, miR-30c, miR-30d, miR-30e mimics and mimic NCs. (B) 354 cases from the TCGA database were divided into two groups by best separation (left) or median separation (right). (https://www.proteinatlas.org/)
References
1.
ChenW.ZhengR.BaadeP.D.ZhangS.ZengH.BrayF.JemalA.YuX.Q. and HeJ., Cancer statistics in China, 2015, CA Cancer J Clin66 (2016), 115–32.
2.
ThrumurthyS.G.ChaudryM.A.ChauI. and AllumW., Does surgery have a role in managing incurable gastric cancer?Nat Rev Clin Oncol12 (2015), 676–82.
3.
UedaT.VoliniaS.OkumuraH.ShimizuM.TaccioliC.RossiS.AlderH.LiuC.G.OueN.YasuiW.YoshidaK.SasakiH.NomuraS.SetoY.KaminishiM.CalinG.A. and CroceC.M., Relation between microRNA expression and progression and prognosis of gastric cancer: a microRNA expression analysis, Lancet Oncology11 (2010), 136–146.
4.
ZuoQ.F.CaoL.Y.YuT.GongL.WangL.N.ZhaoY.L.XiaoB. and ZouQ.M., MicroRNA-22 inhibits tumor growth and metastasis in gastric cancer by directly targeting MMP14 and Snail, Cell Death Dis6 (2015), e2000.
5.
LiB.S.ZuoQ.F.ZhaoY.L.XiaoB.ZhuangY.MaoX.H.WuC.YangS.M.ZengH.ZouQ.M. and GuoG., MicroRNA-25 promotes gastric cancer migration, invasion and proliferation by directly targeting transducer of ERBB2, 1 and correlates with poor survival, Oncogene34 (2015), 2556–65.
6.
ZuoQ.F.ZhangR.LiB.S.ZhaoY.L.ZhuangY.YuT.GongL.LiS.XiaoB. and ZouQ.M., MicroRNA-141 inhibits tumor growth and metastasis in gastric cancer by directly targeting transcriptional co-activator with PDZ-binding motif, TAZ, Cell Death Dis6 (2015), e1623.
7.
WangL.YuT.LiW.LiM.ZuoQ.ZouQ. and XiaoB., The miR-29c-KIAA1199 axis regulates gastric cancer migration by binding with WBP11 and PTP4A3, Oncogene38 (2019), 3134–3150.
8.
PetroccaF.VisoneR.OnelliM.R.ShahM.H.NicolosoM.S.de MartinoI.IliopoulosD.PilozziE.LiuC.G.NegriniM.CavazziniL.VoliniaS.AlderH.RucoL.P.BaldassarreG.CroceC.M. and VecchioneA., E2F1-regulated microRNAs impair TGFbeta-dependent cell-cycle arrest and apoptosis in gastric cancer, Cancer Cell13 (2008), 272–86.
9.
DingL.XuY.ZhangW.DengY.SiM.DuY.YaoH.LiuX.KeY.SiJ. and ZhouT., MiR-375 frequently downregulated in gastric cancer inhibits cell proliferation by targeting JAK2, Cell Res20 (2010), 784–93.
10.
ZhuE.D.LiN.LiB.S.LiW.ZhangW.J.MaoX.H.GuoG.ZouQ.M. and XiaoB., miR-30b, down-regulated in gastric cancer, promotes apoptosis and suppresses tumor growth by targeting plasminogen activator inhibitor-1, PLoS One9 (2014), e106049.
11.
BaraniskinA.Birkenkamp-DemtroderK.MaghnoujA.ZollnerH.MundingJ.Klein-ScoryS.Reinacher-SchickA.Schwarte-WaldhoffI.SchmiegelW. and HahnS.A., MiR-30a-5p suppresses tumor growth in colon carcinoma by targeting DTL, Carcinogenesis33 (2012), 732–9.
12.
LiuM.HuangF.ZhangD.JuJ.WuX.B.WangY.WangY.WuY.NieM.LiZ.MaC.ChenX.ZhouJ.Y.TanR.YangB.L.ZenK.ZhangC.Y.ChenY.G. and ZhaoQ., Heterochromatin protein HP1gamma promotes colorectal cancer progression and is regulated by miR-30a, Cancer Res75 (2015), 4593–604.
13.
ShenH.XingC.CuiK.LiY.ZhangJ.DuR.ZhangX. and LiY., MicroRNA-30a attenuates mutant KRAS-driven colorectal tumorigenesis via direct suppression of ME1, Cell Death Differ24 (2017), 1253–1262.
14.
ZhangN.WangX.HuoQ.SunM.CaiC.LiuZ.HuG. and YangQ., MicroRNA-30a suppresses breast tumor growth and metastasis by targeting metadherin, Oncogene33 (2014), 3119–28.
15.
KumarswamyR.MudduluruG.CeppiP.MuppalaS.KozlowskiM.NiklinskiJ.PapottiM. and AllgayerH., MicroRNA-30a inhibits epithelial-to-mesenchymal transition by targeting Snai1 and is downregulated in non-small cell lung cancer, Int J Cancer130 (2012), 2044–53.
16.
JiangB.Y.ZhangX.C.SuJ.MengW.YangX.N.YangJ.J.ZhouQ.ChenZ.Y.ChenZ.H.XieZ.ChenS.L. and WuY.L., BCL11A overexpression predicts survival and relapse in non-small cell lung cancer and is modulated by microRNA-30a and gene amplification, Molecular Cancer12 (2013), 61.
17.
ZhuJ.ZengY.LiW.QinH.LeiZ.ShenD.GuD.HuangJ.A. and LiuZ., CD73/NT5E is a target of miR-30a-5p and plays an important role in the pathogenesis of non-small cell lung cancer, Mol Cancer16 (2017), 34.
18.
SousaJ.F.NamK.T.PetersenC.P.LeeH.J.YangH.K.KimW.H. and GoldenringJ.R., miR-30-HNF4gamma and miR-194-NR2F2 regulatory networks contribute to the upregulation of metaplasia markers in the stomach, Gut65 (2016), 914–24.
19.
LiuZ.ChenL.ZhangX.XuX.XingH.ZhangY.LiW.YuH.ZengJ. and JiaJ., RUNX3 regulates vimentin expression via miR-30a during epithelial-mesenchymal transition in gastric cancer cells, J Cell Mol Med18 (2014), 610–23.
20.
ParkJ.E.LenterM.C.ZimmermannR.N.Garin-ChesaP.OldL.J. and RettigW.J., Fibroblast activation protein, a dual specificity serine protease expressed in reactive human tumor stromal fibroblasts, J Biol Chem274 (1999), 36505–12.
21.
MoriY.KonoK.MatsumotoY.FujiiH.YamaneT.MitsumataM. and ChenW.T., The expression of a type II transmembrane serine protease (Seprase) in human gastric carcinoma, Oncology67 (2004), 411–9.
22.
IwasaS.OkadaK.ChenW.-T.JinX.YamaneT.OoiA. and MitsumataM., Increased expression of seprase, a membrane-type serine protease, is associated with lymph node metastasis in human colorectal cancer, Cancer Letters227 (2005), 229–236.
23.
GoscinskiM.A.SuoZ.FlorenesV.A.VlatkovicL.NeslandJ.M. and GierckskyK.E., FAP-alpha and uPA show different expression patterns in premalignant and malignant esophageal lesions, Ultrastruct Pathol32 (2008), 89–96.
24.
ShiM.YuD.H.ChenY.ZhaoC.Y.ZhangJ.LiuQ.H.NiC.R. and ZhuM.H., Expression of fibroblast activation protein in human pancreatic adenocarcinoma and its clinicopathological significance, World J Gastroenterol18 (2012), 840–6.
25.
Mhawech-FaucegliaP.WangD.SamraoD.KimG.LawrensonK.MenesesT.LiuS.YessaianA. and PejovicT., Clinical Implications of Marker Expression of Carcinoma-Associated Fibroblasts (CAFs) in Patients with Epithelial Ovarian Carcinoma After Treatment with Neoadjuvant Chemotherapy, Cancer Microenviron7 (2014), 33–9.
26.
WangH.WuQ.LiuZ.LuoX.FanY.LiuY.ZhangY.HuaS.FuQ.ZhaoM.ChenY.FangW. and LvX., Downregulation of FAP suppresses cell proliferation and metastasis through PTEN/PI3K/AKT and Ras-ERK signaling in oral squamous cell carcinoma, Cell Death Dis5 (2014), e1155.
27.
WangR.F.ZhangL.H.ShanL.H.SunW.G.ChaiC.C.WuH.M.IblaJ.C.WangL.F. and LiuJ.R., Effects of the fibroblast activation protein on the invasion and migration of gastric cancer, Exp Mol Pathol95 (2013), 350–356.
28.
ShanL.H.SunW.G.HanW.QiL.YangC.ChaiC.C.YaoK.ZhouQ.F.WuH.M.WangL.F. and LiuJ.R., Roles of fibroblasts from the interface zone in invasion, migration, proliferation and apoptosis of gastric adenocarcinoma, J Clin Pathol65 (2012), 888–95.
29.
YuT.ZuoQ.F.GongL.WangL.N.ZouQ.M. and XiaoB., MicroRNA-491 regulates the proliferation and apoptosis of CD8(+) T cells, Sci Rep6 (2016), 30923.
30.
ZhangQ.TangQ.QinD.YuL.HuangR.LvG.ZouZ.JiangX.C.ZouC.LiuW.LuoJ.ZhaoZ.MuhammadS.WangG.ChenY.G. and WangX., Role of microRNA 30a targeting insulin receptor substrate 2 in colorectal tumorigenesis, Mol Cell Biol35 (2015), 988–1000.
31.
ZouZ.NiM.ZhangJ.ChenY.MaH.QianS.TangL.TangJ.YaoH.ZhaoC.LuX.SunH.QianJ.MaoX.LuX.LiuQ.ZenJ.WuH.BaoZ.LinS.ShengH.LiY.LiangY.ChenZ. and ZongD., miR-30a can inhibit DNA replication by targeting RPA1 thus slowing cancer cell proliferation, Biochem J473 (2016), 2131–9.
32.
ChenZ.ZhangJ.ZhangZ.FengZ.WeiJ.LuJ.FangY.LiangY.CenJ.PanY.HuangY.ZhouF.ChenW. and LuoJ., The putative tumor suppressor microRNA-30a-5p modulates clear cell renal cell carcinoma aggressiveness through repression of ZEB2, Cell Death Dis8 (2017), e2859.
33.
ChangC.W.YuJ.C.HsiehY.H.YaoC.C.ChaoJ.I.ChenP.M.HsiehH.Y.HsiungC.N.ChuH.W.ShenC.Y. and ChengC.W., MicroRNA-30a increases tight junction protein expression to suppress the epithelial-mesenchymal transition and metastasis by targeting Slug in breast cancer, Oncotarget7 (2016), 16462–78.
34.
ZhuH.WuH.LiuX.LiB.ChenY.RenX.LiuC.G. and YangJ.M., Regulation of autophagy by a beclin 1-targeted microRNA, miR-30a, in cancer cells, Autophagy5 (2009), 816–23.
35.
SestitoR.CianfroccaR.RosanoL.TocciP.SemprucciE.Di CastroV.CapraraV.FerrandinaG.SacconiA.BlandinoG. and BagnatoA., miR-30a inhibits endothelin A receptor and chemoresistance in ovarian carcinoma, Oncotarget7 (2016), 4009–23.
36.
LiuX.JiQ.ZhangC.LiuX.LiuY.LiuN.SuiH.ZhouL.WangS. and LiQ., miR-30a acts as a tumor suppressor by double-targeting COX-2 and BCL9 in H. pylori gastric cancer models, Sci Rep7 (2017), 7113.
37.
LiX.ZhangY.ZhangY.DingJ.WuK. and FanD., Survival prediction of gastric cancer by a seven-microRNA signature, Gut59 (2010), 579–85.
38.
KandaM. and KoderaY., Molecular mechanisms of peritoneal dissemination in gastric cancer, World J Gastroenterol22 (2016), 6829–40.
39.
KoczorowskaM.M.TholenS.BucherF.LutzL.KizhakkedathuJ.N.De WeverO.WellnerU.F.BiniossekM.L.StahlA.LassmannS. and SchillingO., Fibroblast activation protein-alpha, a stromal cell surface protease, shapes key features of cancer associated fibroblasts through proteome and degradome alterations, Mol Oncol10 (2016), 40–58.
40.
ZhangY.TangH.CaiJ.ZhangT.GuoJ.FengD. and WangZ., Ovarian cancer-associated fibroblasts contribute to epithelial ovarian carcinoma metastasis by promoting angiogenesis, lymphangiogenesis and tumor cell invasion, Cancer Lett303 (2011), 47–55.
41.
JacksonK.W.ChristiansenV.J.YadavV.R.Silasi-MansatR.LupuF.AwasthiV.ZhangR.R. and McKeeP.A., Suppression of tumor growth in mice by rationally designed pseudopeptide inhibitors of fibroblast activation protein and prolyl oligopeptidase, Neoplasia17 (2015), 43–54.
42.
FangJ.XiaoL.JooK.I.LiuY.ZhangC.LiuS.ContiP.S.LiZ. and WangP., A potent immunotoxin targeting fibroblast activation protein for treatment of breast cancer in mice, Int J Cancer138 (2016), 1013–23.
43.
ChristiansenV.J.JacksonK.W.LeeK.N.DownsT.D. and McKeeP.A., Targeting inhibition of fibroblast activation protein-alpha and prolyl oligopeptidase activities on cells common to metastatic tumor microenvironments, Neoplasia15 (2013), 348–58.
44.
LoA.WangL.C.SchollerJ.MonslowJ.AveryD.NewickK.O’BrienS.EvansR.A.BajorD.J.ClendeninC.DurhamA.C.BuzaE.L.VonderheideR.H.JuneC.H.AlbeldaS.M. and PureE., Tumor-Promoting Desmoplasia Is Disrupted by Depleting FAP-Expressing Stromal Cells, Cancer Res75 (2015), 2800–10.
45.
SaadiA.ShannonN.B.Lao-SirieixP.O’DonovanM.WalkerE.ClemonsN.J.HardwickJ.S.ZhangC.DasM.SaveV.NovelliM.BalkwillF. and FitzgeraldR.C., Stromal genes discriminate preinvasive from invasive disease, predict outcome, and highlight inflammatory pathways in digestive cancers, Proc Natl Acad Sci U S A107 (2010), 2177–82.
46.
LiuF.QiL.LiuB.LiuJ.ZhangH.CheD.CaoJ.ShenJ.GengJ.BiY.YeL.PanB. and YuY., Fibroblast activation protein overexpression and clinical implications in solid tumors: a meta-analysis, PLoS One10 (2015), e0116683.
47.
WuX.ChenX.ZhouQ.LiP.YuB.LiJ.QuY.YanJ.YuY.YanM.ZhuZ.LiuB. and SuL., Hepatocyte growth factor activates tumor stromal fibroblasts to promote tumorigenesis in gastric cancer, Cancer Lett335 (2013), 128–35.