Dkk3 protein attenuates the expression of Wnt3a, Wnt5a and LRP6, and their interaction, and interacts with TrCP to suppress wnt/-catenin pathway.
METHODS:
We performed a bioinformatics analysis of Dkk3 mRNA expression through Oncomine, TCGA and Kaplan-Meier plotter databases up to July 10, 2017.
RESULTS:
Up-regulated Dkk3 expression was higher in gastric, breast, and ovarian cancers than normal tissues ( 0.05). Bitter’s database showed a higher Dkk3 expression in ovarian cytoadenocarcinoma than clear cell adenocarcinoma ( 0.05). Dkk3 was more expressed in ductal breast cancer in situ than invasive ductal breast cancer ( 0.05), in mixed lobular and ductal cancer, and lobular cancer than ductal breast cancer ( 0.05). In TCGA data, Dkk3 expression was lower in gastric cancers with than without Barret’s esophagus ( 0.05), in intestinal-type than diffuse-type cancers ( 0.05), and in the cancers of elder than younger patients ( 0.05). Dkk3 expression was higher in squamous cell carcinoma than adenocarcinoma ( 0.05). Dkk3 expression was higher in ductal than lobular breast cancer, or in younger than elder patients with breast cancer ( 0.05). According to Kaplan-Meier plotter, Dkk3 expression was negatively correlated with overall, progression-free, relapse-free or distant-metastasis-free survival rate of gastric, breast or ovarian cancer patients, but versa for lung cancer patients ( 0.05).
CONCLUSION:
Dkk3 expression might be employed as a potential marker to indicate carcinogenesis and histogenesis, even prognosis.
REIC (Reduced Expression in Immortalized Cells) was identified by the representational difference analysis system because of its lower expression in immortalized than parental cells. A homology search demonstrates that REIC is identical with the human Dkk3 gene (1). Dkk3b transcript is recently found to originate from a second transcriptional start site located in intron 2 of Dkk3 gene [2]. Dkk3-encoding glycoprotein includes an N-terminal signal peptide, two cysteine-rich domains and two coiled-coil domain [3]. Dkk3 protein attenuates the expression of Wnt3a, Wnt5a and LRP6, and their interaction, thereby suppressing both canonical and non-canonical Wnt signal cascades [4]. Dkk3 can interact with TrCP to block the nuclear translocation of -catenin, down-regulating the transcription of its downstream VEGF and Cylcin D1 [4]. Dkk3b also interrupts nuclear translocation of -catenin by capturing cytoplasmic and unphosphorylated -catenin via an extra-nuclear complex with TrCP [2]. Dkk-3-induced JNK activation is largely mediated through endoplasmic reticulum stress and mitochondrial pathway [5, 6]. Dkk-3
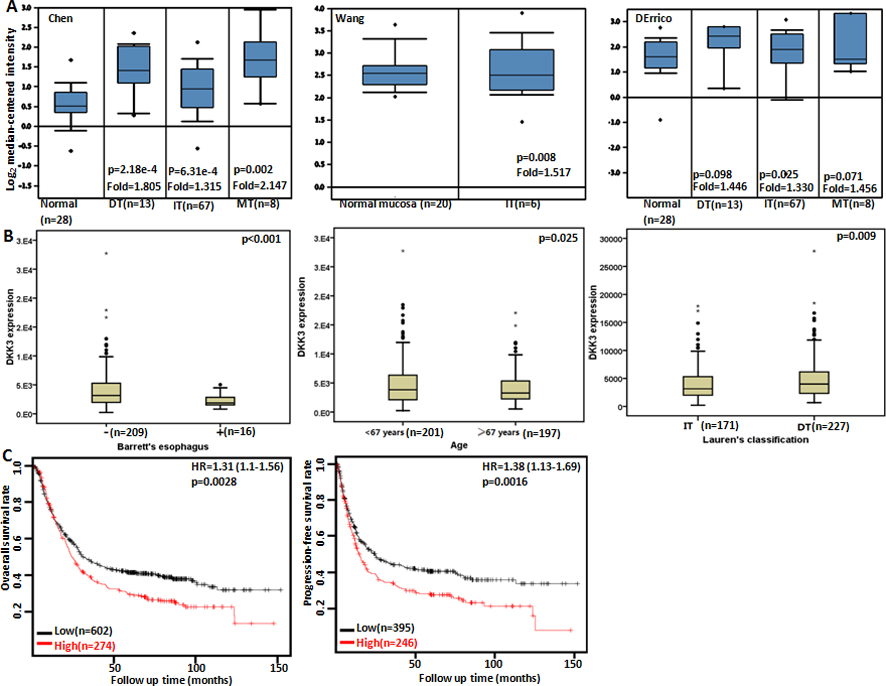
The clinicopathological and prognostic significances of Dkk3 mRNA expression in gastric cancer. According to Chen’s, Wang’s and DErrico’s database, a higher Dkk3 expression was detectable in gastric cancer than that in normal gastric mucosa (A, 0.05), even stratified into intestinal-type (IT), diffuse-type (DT) and mixed-type carcinomas. TCGA database showed that REIC expression was inversely linked to the existence of Barret’s esophagus, younger gage and differentiation (B, 0.05). According to the data from Kaplan-Meier plotter, Dkk3 expression was negatively related to both overall and progression-free survival rates of the patients with gastric cancer(C, 0.05). HR, hazard ratio.
interacts with SGTA and restores androgen receptor signaling via N-terminal regions of both proteins in androgen-independent prostate cancer cells [7, 8]. MiRNA-92b inhibitor promotes glioma cell apoptosis via targeting Dkk3 and blocking Wnt/ -catenin signaling pathway [9]. MiRNA-25 markedly increases invasion and proliferation of melanoma cells by targeting Dkk3 and through Wnt/-catenin pathway [10].
The prognostic significance of Dkk3 mRNA expression in gastric cancer
Clinicopathological features
Overall survival
Progression-free survival
Hazard ratio
Hazard ratio
Sex
Female
1.7 (1.2–2.41)
0.0025
1.92 (1.3–2.85)
0.00092
Male
1.33 (1.06–1.66)
0.015
1.33 (1.04–1.71)
0.022
T
2
2.44 (1.59–3.73)
2.3e-05
2.29 (1.38–3.81)
0.00095
3
1.32 (0.92–1.89)
0.13
1.24 (0.87–1.75)
0.23
4
3.59 (1.44–8.96)
0.0036
3.76 (1.56–9.03)
0.0017
N
0
3.63 (1.23–10.68)
0.012
3.84 (1.3–11.32)
0.0086
1–3
2.02 (1.53–2.66)
3.4e-07
1.9 (1.45–2.48)
2.1e-06
1
1.9 (1.26–2.87)
0.002
1.75 (1.16–2.63)
0.0063
2
1.73 (1.07–2.8)
0.024
1.86 (1.17–2.96)
0.0079
3
2.36 (1.36–4.11)
0.0017
1.95 (1.13–3.37)
0.014
M
0
1.89 (1.43–2.49)
4.8e-06
1.78 (1.34–2.35)
4.2e-05
1
1.51 (0.81–2.81)
0.19
0.69 (0.38–1.25)
0.22
TNM staging
I
0.46 (0.13–1.63)
0.22
0.54 (0.15–1.97)
0.34
II
2.02 (1.11–3.67)
0.019
2.62 (1.17–5.9)
0.015
III
1.15 (0.83–1.59)
0.41
1.82 (1.22–2.7)
0.0026
IV
1.66 (1.13–2.45)
0.0097
1.39 (0.94–2.05)
0.093
Differentiation
Well-differentiated
2.76 (0.81–9.41)
0.091
–
–
Moderately-differentiated
1.72 (0.9–3.29)
0.097
1.86 (0.86–4.06)
0.11
Poorly-differentiated
1.54 (1.04–2.3)
0.032
2.17 (1.17–4.04)
0.012
Lauren’s classification
Intestinal-type
1.22 (0.86–1.72)
0.27
1.45 (1–2.1)
0.049
Diffuse-type
1.98 (1.4–2.8)
7.6e-05
2.08 (1.47–2.94)
2.5e-05
Mixed-type
6.8 (0.89–52.04)
0.033
7.1 (0.93–53.9)
0.027
Her2 positivity
1.45 (1.15–1.81)
0.0012
1.51 (1.15–2)
0.0031
1.49 (1.15–1.93)
0.0024
1.57 (1.11–2.22)
0.0094
Perforation
1.94 (1.29–2.91)
0.0011
2.32 (1.39–3.86)
0.00089
Treatment
Surgery alone
1.84 (1.38–2.46)
2.5e-05
1.88 (1.35–2.62)
0.00016
5-FU-based adjuvant
0.77 (0.52–1.14)
0.19
0.83 (0.57–1.22)
0.34
Other adjuvant
2.43 (0.99–5.96)
0.044
2.07 (0.93–4.62)
0.069
Although Dkk3 protein exists in various normal human tissues, such as brain, heart, liver, pancreas, kidney, mammary gland and lymph node, Dkk3 expression is down-regulated not only in the tumor-derived cell lines, but also in different types of human cancers, including cervical cancer, seminoma, renal clear cell carcinoma, prostate cancer and non-small cell lung cancers due to its promoter methylation [1, 4, 6, 11, 12, 13, 14, 15, 16]. Most noteworthy, Dkk3 methylation was correlated significantly with a poor prognosis of hepatocellular, gastric or breast cancers [17, 18, 19]. Our group also found that Dkk3 expression was negatively associated with tumor size, lymph node metastasis, dedifferentiation or poor prognosis of gastric cancer. The serum Dkk3 level was significantly higher in healthy individuals than gastric cancer patients and inversely linked to tumor size [20]. In the present study, we performed a bioinformatics analysis to clarify the clinicopathological and prognostic significances of Dkk3 mRNA expression in various cancers.
Materials and methods
Oncomine database analysis
The individual gene expression level of Dkk3 was analyzed using Oncomine (www.oncomine.org), a cancer microarray database and web-based data mining platform for a new discovery from genome-wide expression analyses. We compared the differences in Dkk3 mRNA level between normal and cancer tissues. All data were log-transformed, median centered per array, and standard deviation normalized to one per array.
TCGA database analysis
The expression data (RNA-seqV2) and clinicopathological data of gastric ( 392), lung ( 865), breast ( 1093) and ovarian ( 304) cancer patients were downloaded from the Cancer Genome Atlas (TCGA) database by TCGA-assembler in R software. We integrated the raw data, analyzed Dkk3 mRNA expression in these cancers, and compared it with clinicopathological and prognostic data of the cancer patients. The means was compared with student t test. Kaplan-Meier survival plots were generated with survival curves compared by log-rank statistic. Cox’s proportional hazards model was employed for multivariate analysis. Two-sided 0.05 was considered as statistically significant. SPSS 17.0 software was employed to analyze all data.
Kaplan-Meier plotter analysis
The prognostic significance of Dkk3 mRNA was also analyzed in gastric, lung, breast and ovarian cancers using Kaplan-Meier plotter (http://kmplot.com).
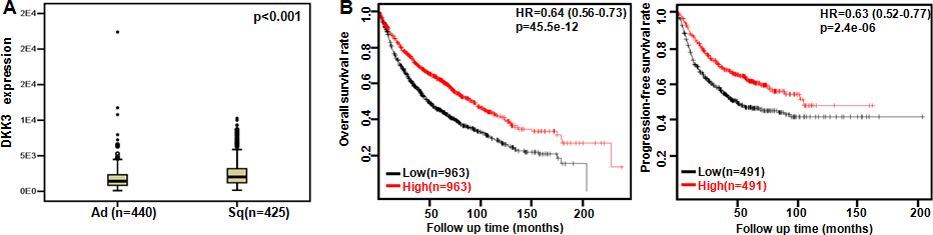
The clinicopathological and prognostic significances of Dkk3 mRNA expression in lung cancer. TCGA database showed that Dkk3 expression was lower in adenocarcinoma (Ad) than squamous cell carcinoma (Sq, A, 0.05). A positive correlation between Dkk3 expression and overall or progression-free survival rate of the patients with lung cancer was found according to Kaplan-Meier plotter (B, 0.05). HR, hazard ratio.
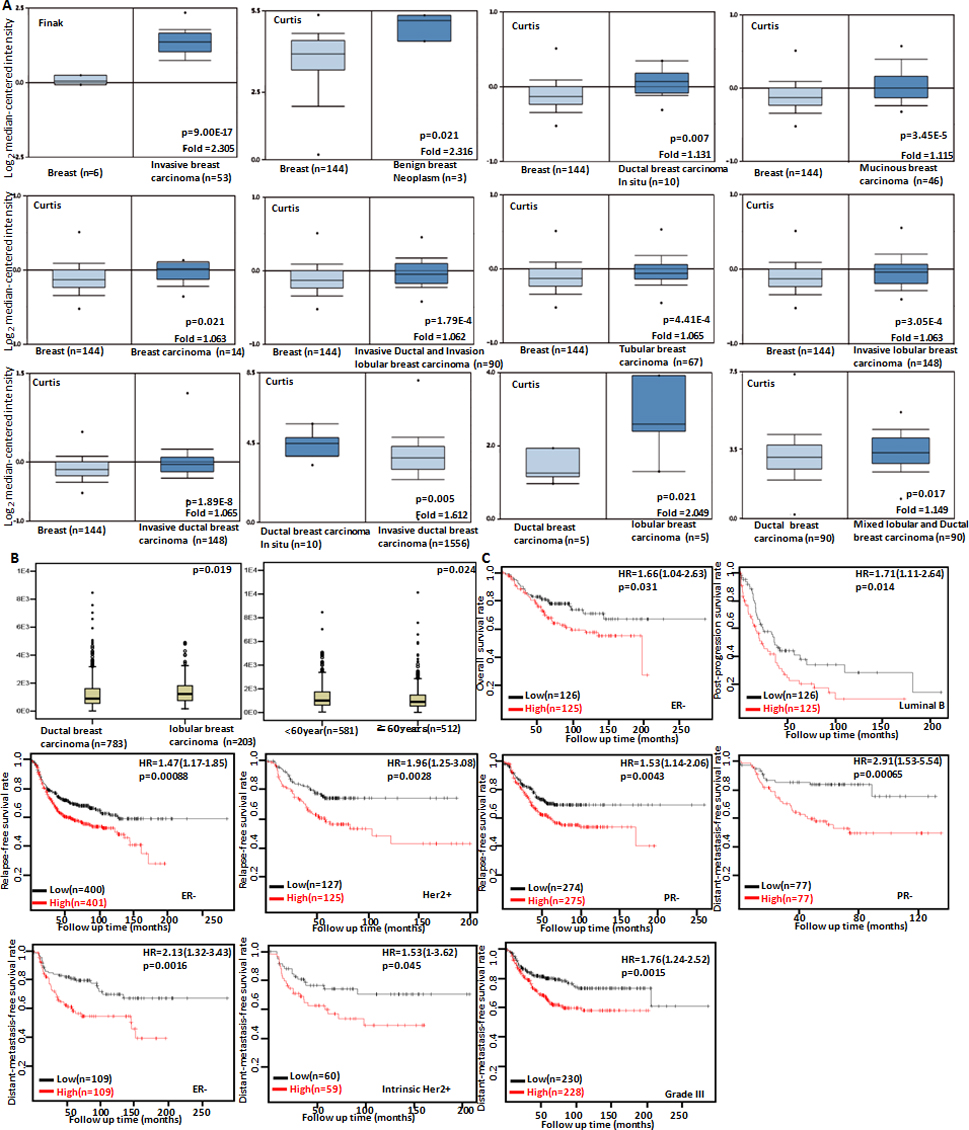
The clinicopathological and prognostic significances of Dkk3 mRNA expression in breast cancer. According to Finak’s and Curtis’s data, Dkk3 hyoexpression was detectable in normal breast tissue, compared with breast benign tumors, breast cancer in situ, invasive breast cancer, mucinous and tubular breast cancers (A, 0.05). Dkk3 expression was inversely linked to invasion of breast cancer (A, 0.05). It was higher in mixed lobular and ductal cancer, and lobular cancer than ductal breast cancer (A, 0.05). TCGA database indicated that Dkk3 expression was higher in ductal than lobular breast cancer, or in younger than elder patients with breast cancer (B, 0.05). The correlation between Dkk3 expression and overall, post-progression or distant-metastasis-free or relapse-free survival rate of the patients with breast cancer was analyzed according to Kaplan-Meier plotter, even stratified by different clinicopathological parameters (C, 0.05). HR, hazard ratio.
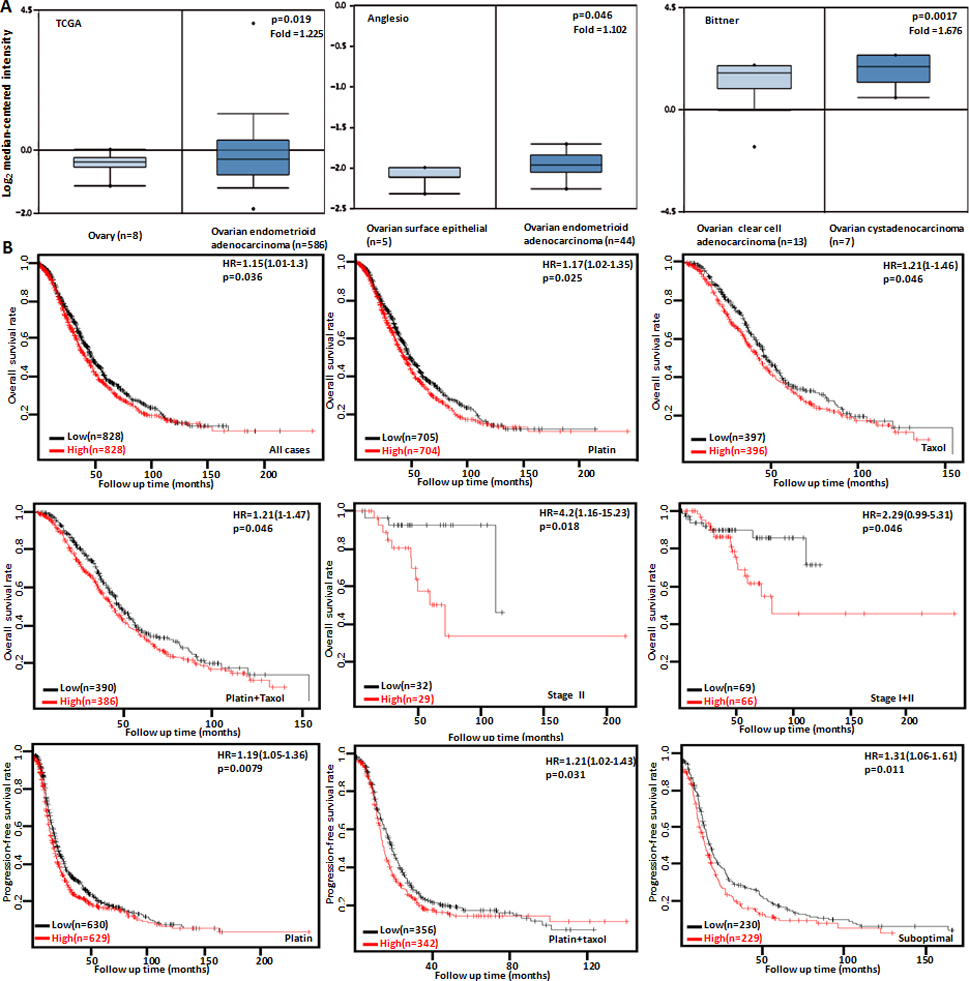
The clinicopathological and prognostic significances of Dkk3 mRNA expression in ovarian cancer. TCGA’s and Anglesio’s datasets were employed for bioinformatics analysis to observe Dkk3 expression in ovarian cancer. A lower Dkk3 expression was detectable in ovary or ovarian surface epithelium than that in endometrioid cancer (A, 0.05). Bitter’s database showed a higher Dkk3 expression in ovarian cytoadenocarcinoma than clear cell adenocarcinoma (A, 0.05). The correlation between Dkk3 expression and overall, or progression-free survival rate of the patients with breast cancer was analyzed according to Kaplan-Meier analysis, even stratified by different clinicopathological parameters (B, 0.05). HR, hazard ratio.
Results
The clinicopathological and prognostic significances of Dkk3 mRNA expression in gastric cancer
We used Chen’s, Wang’s and DErrico’s datasets to perform bioinformatics analysis and found that Dkk3 expression was higher in gastric cancer than normal tissues, even stratified into intestinal-, diffuse- and mixed-type carcinomas (Fig. 1A, 0.05). In TCGA data, Dkk3 expression was lower in gastric cancers with than without Barret’s esophagus (Fig. 1B, 0.05), and in the cancers of elder than younger patients (Fig. 1B, 0.05). There was a lower Dkk3 expression in intestinal-type than diffuse-type cancers (Fig. 1B, 0.05). According to Kaplan-Meier plotter, we found that a higher Dkk3 expression was negatively correlated with overall and progression-free survival rates of all cancer patients, even stratified by gender, lymph node status, and Her2 positivity (Fig. 1C and Table 1, 0.05). It was the same for the patients with T2, T4, M0, stage II, poorly-differentiated, diffuse-type or mixed-type cancer, or the patients receiving surgical treatment alone (Table 1, 0.05). The overall survival rate of the patient with stage-IV cancers or receiving other adjuvant was lower in the group of high Dkk3 expression than its low expression (Table 1, 0.05). T4, stage III or intestinal-type cancer patients with high Dkk3 expression showed a shorter progression- free survival time than those with its low expression (Table 1, 0.05).
The clinicopathological and prognostic signifi- cances of Dkk3 mRNA expression in lung cancer
In TCGA data, Dkk3 expression was higher in squamous cell carcinoma than adenocarcinoma (Fig. 2A, 0.05). According to Kaplan-Meier plotter, we found that a higher Dkk3 expression was positively correlated with overall and progression-free survival rates of all, male, female, N1, smoking, nonsmoking, Grade II, cancer patients, and the cancer patients with surgical margin negative (Fig. 2B, 0.05). N0, T1, T2, M0, or stage I cancer patients with high Dkk3 expression showed a long progression-free survival time than those with its low expression ( 0.05, data not shown). There appeared a positive relationship between Dkk3 expression and the post-progression survival rate of female or non-smoking cancer patients ( 0.05, data not shown).
The clinicopathological and prognostic significances of Dkk3 mRNA expression in breast cancer
We used Finak’s and Curtis’s datasets to perform bioinformatics analysis and found that Dkk3 mRNA expression was lower in normal breast tissue than breast benign tumors, invasive or ductal breast cancer in situ, invasive ductal or lobular breast cancer, mucinous and tubular breast cancers (Fig. 3A, 0.05). Dkk3 was more expressed in ductal breast cancer in situ than invasive ductal breast cancer (Fig. 3A, 0.05). There was a higher Dkk3 expression in in mixed lobular and ductal cancer, and lobular cancer than ductal breast cancer (Fig. 3A, 0.05). TCGA database indicated that Dkk3 expression was higher in ductal than lobular breast cancer, or in younger than elder patients with breast cancer (Fig. 3B, 0.05). According to Kaplan-Meier plotter, we found that a higher Dkk3 expression was negatively correlated with overall survival rate of ER-negative cancer patients, and post-progression survival rate of Luminal-B cancer patients (Fig. 3C, 0.05). ER-negative, Her2-positive or PR-negative cancer patients with high Dkk3 expression showed a shorter relapse-free survival time than those with its low expression (Fig. 3C, 0.05). There appeared a negative relationship between Dkk3 expression and the distant-metastasis-free survival rate of PR-negative, ER-negative, intrinsic-type and Her-2-positive, or Grade-III carcinoma patients ( 0.05).
The clinicopathological and prognostic significances of Dkk3 mRNA expression in ovarian cancer
According to TCGA’s and Anglesio’s datasets, we found a lower Dkk3 expression in ovary or ovarian surface epithelium than that in endometrioid cancer (Fig. 4A, 0.05). Bitter’s database showed a higher Dkk3 expression in ovarian cytoadenocarcinoma than clear cell adenocarcinoma (Fig. 4A, 0.05). In terms of Kaplan-Meier plotter, we found that a higher Dkk3 expression was negatively correlated with overall survival rate of all cancer patients, the patients receiving the treatment of platin, taxol or platin taxol and the stage-II or stage-II+III cancer patients (Fig. 4B, 0.05). There appeared a positive relationship between Dkk3 expression and the progression-free survival rate of the suboptimal-type cancer patients or the patients receiving the treatment of platin or platin taxol (Fig. 4B, 0.05).
Discussion
Dkk3 was reported to suppress cellular proliferation, invasion, migration, and tumor cell survival in head and neck squamous cell carcinoma cells via PI3K-Akt inactivation [21]. Dkk3 overexpression resulted in slower cell growth by an extension of G phase, promoted survival of microcolonies, and resulted in significant impairment of migratory and invasive behaviors, largely attributable to modified cell adhesions and adhesion kinetics in adrenal cortex carcinoma cells [22]. Ad-SGE-REIC reduced cell viability and tumor growth, induced apoptosis in hepatocellular carcinoma cells [23]. It reduced the tumor growth of lung cancer, malignant mesothelioma and pancreatic cancer cells not only of directly treated tumors but also of distant untreated tumors [24, 25]. Its intraperitoneal administration suppressed peritoneal dissemination of scirrhous gastric cancer [26]. Edamura et al. [27] demonstrated that the Ad-REIC treatment inhibited prostate cancer growth and lymph node metastasis, and prolonged mice survival in the model by intratumoral apoptotic induction and in vitro suppression of cell invasion/migration with reduced MMP-2 activity. These findings indicate that Dkk3 may be considered as a molecule target of gene therapy for cancers.
Dkk3 staining was decreased from epidermis in normal skin, actinic keratosis, squamous cell carcinoma in situ, to keratoacanthoma and squamous cell carcinoma [28]. A low expression of Dkk3 was found in pancreatic cancer, renal cell carcinoma, thyroid cancer, breast cancer, and colorectal cancer [16, 29, 30, 31, 32, 33]. Dkk3 mRNA was also down-regulated in most cancer samples including the kidney cancer, urinary bladder cancer, prostate cancer, pancreatic cancer, hepatocellular cancer and lung cancer [3, 34, 35]. In contrast, we found up-regulated Dkk3 mRNA expression in gastric cancer, breast cancer and ovarian cancer. The paradoxical results might be due to different methods: previous studies are performed by immunostaining or real-time PCR; our bioinformatics analysis is based on cDNA microarray. Additionally, we found that Dkk3 was more expressed in ductal breast cancer in situ than invasive ductal breast cancer, in line with other reports [13, 30, 34]. Shin et al. [28] found that Dkk-3 significantly decreased the proliferation, migration and tumor growth of cutaneous squamous cell carcinoma cells with Cyclin D1 and CDK4/6 hypoexpression and p21 hyperexpression. Dkk3 also blocked translocation of -catenin/EMT (epithelial-mesenchymal transition) induced by hypoxia and improved gemcitabine therapeutic effect in pancreatic cancer cells [29]. In our work, a higher Dkk3 expression was in diffuse-type than intestinal-type cancers in ovarian cytoadenocarcinoma than clear cell adenocarcinoma, in ductal than lobular breast cancer, and in squamous cell carcinoma than adenocarcinoma, suggesting that Dkk3 has distinct expression levels in different histological subtypings, and underlies the molecular mechanisms of cancer histogenesis.
Dkk3 promoter methylation was a poor prognostic factor for the overall and disease-free survival of breast cancer patients [35], while cervical cancer patients with low Dkk3 expression had a significantly lower 5-year disease-free survival rate than those with high Dkk3 expression [13]. Dkk3 expression was negatively correlated with overall poor prognosis of patients with gastric cancer [20], pancreatic cancer [36] or head and neck squamous cell carcinoma [37]. Yang et al. [17] found that that the progression-free survival time of hepatocellular patients with a higher Dkk3 methylation level was significantly shorter than that of those with a lower Dkk3 methylation level as an independent factor. According to Kaplan-Meier plotter, we found that Dkk3 mRNA expression was negatively correlated with overall, progression-free, relapse-free or distant- metastasis-free survival rate of gastric, breast and ovarian cancer patients, but versa for lung cancer patients. This result may be attributable to differences in tissue specificity and detection methods.
In conclusion, Dkk3 mRNA expression underwent an up-regulation during gastric, breast and ovarian carcinogenesis. It was negatively correlated with the invasion of breast cancer and underlay the molecular mechanism of the histogenesis of lung, breast and ovarian cancers. Dkk3 expression might be employed as a potential marker for the prognosis of the cancer patients.
Footnotes
Acknowledgments
This study was supported by Liaoning BaiQianWan Talents Program, Outstanding Scientific Fund of Shengjing Hospital, Award for Liaoning Distinguished Professor, Shenyang Science and Technology Grand (18-013-0-59); A Key Scientific and Technological Project of Liaoning Province (2015408001) and National Natural Scientific Foundation of China (81472544; 81672700).
Conflict of interest
The authors have declared that no competing interests exist.
References
1.
TsujiT.MiyazakiM.SakaguchiM.InoueY. and NambaM., A REIC gene shows down-regulation in human immortalized cells and human tumor-derived cell lines, Biochem Biophys Res Commun268 (2000), 20–24.
2.
LeonardJ.L.LeonardD.M.WolfeS.A.LiuJ.RiveraJ.YangM.LeonardR.T.JohnsonJ.P.S.KumarP.LiebmannK.L.TuttoA.A.MouZ. and SiminK.J., The Dkk3 gene encodes a vital intracellular regulator of cell proliferation, PLoS One12 (2017), e0181724.
3.
HsiehS.Y.HsiehP.S.ChiuC.T. and ChenW.Y., Dickkopf-3/REIC functions as a suppressor gene of tumor growth, Oncogene23 (2004), 9183–9189.
4.
LeeE.J.JoM.RhoS.B.ParkK.YooY.N.ParkJ.ChaeM.ZhangW. and LeeJ.H., Dkk3, downregulated in cervical cancer, functions as a negative regulator of beta-catenin, Int J Cancer124 (2009), 287–297.
5.
HaraK.KagejiT.MizobuchiY.KitazatoK.T.OkazakiT.FujiharaT.NakajimaK.MureH.KuwayamaK.HaraT. and NagahiroS., Blocking of the interaction between Wnt proteins and their co-receptors contributes to the anti-tumor effects of adenovirus-mediated DKK3 in glioblastoma, Cancer Lett356 (2015), 496–505.
6.
AbarzuaF.SakaguchiM.TakaishiM.NasuY.KuroseK.EbaraS.MiyazakiM.NambaM.KumonH. and HuhN.H., Adenovirus-mediated overexpression of REIC/Dkk-3 selectively induces apoptosis in human prostate cancer cells through activation of c-Jun-NH2-kinase, Cancer Res65 (2005), 9617–9622.
7.
KatoY.OchiaiK.KawakamiS.NakaoN.AzakamiD.BonkobaraM.MichishitaM.MorimatsuM.WatanabeM. and OmiT., Canine REIC/Dkk-3 interacts with SGTA and restores androgen receptor signalling in androgen-independent prostate cancer cell lines, BMC Vet Res13 (2017), 170.
8.
OchiaiK.MorimatsuM.KatoY.Ishiguro-OonumaT.UdagawaC.RungsuriyawiboonO.AzakamiD.MichishitaM.AriyoshiY.UekiH.NasuY.KumonH.WatanabeM. and OmiT., Tumor suppressor REIC/DKK-3 and co-chaperone SGTA: Their interaction and roles in the androgen sensitivity, Oncotarget7 (2016), 3283–3296.
9.
LiQ.ShenK.ZhaoY.MaC.LiuJ. and MaJ., MiR-92b inhibitor promoted glioma cell apoptosis via targeting DKK3 and blocking the Wnt/beta-catenin signaling pathway, J Transl Med11 (2013), 302.
10.
HuoJ.ZhangY.LiR.WangY.WuJ. and ZhangD., Upregulated MicroRNA-25 mediates the migration of melanoma cells by targeting DKK3 through the WNT/β-Catenin pathway, Int J Mol Sci17 (2016), E1124.
11.
KobayashiK.OuchidaM.TsujiT.HanafusaH.MiyazakiM.NambaM.ShimizuN. and ShimizuK., Reduced expression of the REIC/Dkk-3 gene by promoter-hypermethylation in human tumor cells, Gene282 (2002), 151–158.
12.
HayashiT.AsanoH.ToyookaS.TsukudaK.SohJ.ShienT.TairaN.MakiY.TanakaN.DoiharaH.NasuY.HuhN.H. and MiyoshiS., DNA methylation status of REIC/Dkk-3 gene in human malignancies, J Cancer Res Clin Oncol138 (2012), 799–809.
13.
RyuS.W.KimJ.H.KimM.K.LeeY.J.ParkJ.S.ParkH.M.KimD.H.LeeS.H. and LeeE.J., Reduced expression of DKK3 is associated with adverse clinical outcomes of uterine cervical squamous cell carcinoma, Int J Gynecol Cancer23 (2013), 134–140.
14.
NozakiI.TsujiT.IijimaO.OhmuraY.AndouA.MiyazakiM.ShimizuN. and NambaM., Reduced expression of REIC/Dkk-3 gene in non-small cell lung cancer, Int J Oncol19 (2001), 117–121.
15.
KuroseK.SakaguchiM.NasuY.EbaraS.KakuH.KariyamaR.AraoY.MiyazakiM.TsushimaT.NambaM.KumonH. and HuhN.H., Decreased expression of REIC/Dkk-3 in human renal clear cell carcinoma, J Urol171 (2004), 1314–1318.
16.
TanimotoR.AbarzuaF.SakaguchiM.TakaishiM.NasuY.KumonH. and HuhN.H., REIC/Dkk-3 as a potential gene therapeutic agent against human testicular cancer, Int J Mol Med19 (2007), 363–368.
17.
YangB.DuZ.GaoY.T.LouC.ZhangS.G.BaiT.WangY.J. and SongW.Q., Methylation of Dickkopf-3 as a prognostic factor in cirrhosis-related hepatocellular carcinoma, World J Gastroenterol16 (2010), 755–763.
18.
YuJ.TaoQ.ChengY.Y.LeeK.Y.NgS.S.CheungK.F.TianL.RhaS.Y.NeumannU.RöckenC.EbertM.P.ChanF.K. and SungJ.J., Promoter methylation of the Wnt/beta-catenin signaling antagonist Dkk-3 is associated with poor survival in gastric cancer, Cancer115 (2009), 49–60.
19.
VeeckJ.WildP.J.FuchsT.SchüfflerP.J.HartmannA.KnüchelR. and DahlE., Prognostic relevance of Wnt-inhibitory factor-1 (WIF1) and Dickkopf-3 (DKK3) promoter methylation in human breast cancer, BMC Cancer9 (2009), 217.
20.
XuX.Y.XiaP.YuM.NieX.C.YangX.XingY.N.LiuY.P.TakanoY. and ZhengH.C., The roles of REIC gene and its encoding product in gastric carcinoma, Cell Cycle11 (2012), 1414–1431.
21.
KataseN.NishimatsuS.I.YamauchiA.YamamuraM.TeradaK.ItadaniM.OkadaN.HassanN.M.M.NagatsukaH.IkedaT.NohnoT. and FujitaS., DKK3 overexpression increases malignant properties of head and neck squamous cell carcinoma cells, Oncol Res in press.
22.
ChengJ.Y.BrownT.C.MurthaT.D.StenmanA.JuhlinC.C.LarssonC.HealyJ.M.PrasadM.L.KnoefelW.T.KriegA.SchollU.I.KorahR. and CarlingT., A novel FOXO1-mediated dedifferentiation blocking role for DKK3 in adrenocortical carcinogenesis, BMC Cancer17 (2017), 164.
23.
SawaharaH.ShirahaH.UchidaD.KatoH.NagaharaT.IwamuroM.KataokaJ.HoriguchiS.WatanabeM.SakaguchiM.TakakiA.NousoK.NasuY.KumonH. and OkadaH., Novel REIC/Dkk-3 encoding adenoviral vector as a promising therapeutic agent for pancreatic cancer, Cancer Gene Ther23 (2016), 278–283.
24.
SuzawaK.ShienK.PengH.SakaguchiM.WatanabeM.HashidaS.MakiY.YamamotoH.TomidaS.SohJ.AsanoH.TsukudaK.NasuY.KumonH.MiyoshiS. and ToyookaS., Distant bystander effect of REIC/DKK3 gene therapy through immune system stimulation in thoracic malignancies, Anticancer Res37 (2017), 301–307.
25.
UchidaD.ShirahaH.KatoH.NagaharaT.IwamuroM.KataokaJ.HoriguchiS.WatanabeM.TakakiA.NousoK.NasuY.YagiT.KumonH. and YamamotoK., Potential of adenovirus-mediated REIC/Dkk-3 gene therapy for use in the treatment of pancreatic cancer, J Gastroenterol Hepatol29 (2014), 973–983.
26.
ThanS.S.KataokaK.SakaguchiM.MurataH.AbarzuaF.TaketaC.DuG.YashiroM.YanagiharaK.NasuY.KumonH. and HuhN.H., Intraperitoneal administration of an adenovirus vector carrying REIC/Dkk-3 suppresses peritoneal dissemination of scirrhous gastric carcinoma, Oncol Rep25 (2011), 989–995.
27.
EdamuraK.NasuY.TakaishiM.KobayashiT.AbarzuaF.SakaguchiM.KashiwakuraY.EbaraS.SaikaT.WatanabeM.HuhN.H. and KumonH., Adenovirus-mediated REIC/Dkk-3 gene transfer inhibits tumor growth and metastasis in an orthotopic prostate cancer model, Cancer Gene Ther14 (2007), 765–772.
28.
ShinJ.M.ChoiD.K.KangH.Y.SohnK.C.LeeY.KimC.D.LeeJ.H. and ParkB.C., The expression pattern and functional role of REIC/Dkk-3 in the development of cutaneous squamous cell carcinoma, J Dermatol Sci84 (2016), 88–96.
29.
GuoQ. and QinW., DKK3 blocked translocation of β-catenin/EMT induced by hypoxia and improved gemcitabine therapeutic effect in pancreatic cancer Bxpc-3 cell, J Cell Mol Med19 (2015), 2832–2841.
30.
YinD.T.WuW.LiM.WangQ.E.LiH.WangY.TangY. and XingM., DKK3 is a potential tumor suppressor gene in papillary thyroid carcinoma, Endocr Relat Cancer20 (2013), 507–514.
31.
GuoC.C.ZhangX.L.YangB.GengJ.PengB. and ZhengJ.H., Decreased expression of Dkk1 and Dkk3 in human clear cell renal cell carcinoma, Mol Med Rep9 (2014), 2367–2373.
32.
VeeckJ.BektasN.HartmannA.KristiansenG.HeindrichsU.KnüchelR. and DahlE., Wnt signalling in human breast cancer: expression of the putative Wnt inhibitor Dickkopf-3 (DKK3) is frequently suppressed by promoter hypermethylation in mammary tumours, Breast Cancer Res10 (2008), R82.
33.
WangW.ZhuW.XuX.Y.NieX.C.YangX.XingY.N.YuM.LiuY.P.TakanoY. and ZhengH.C., The clinicopathological significance of REIC expression in colorectal carcinomas, Histol Histopathol27 (2012), 735–743.
34.
DingZ.QianY.B.ZhuL.X. and XiongQ.R., Promoter methylation and mRNA expression of DKK-3 and WIF-1 in hepatocellular carcinoma, World J Gastroenterol15 (2009), 2595–2601.
35.
KashiwakuraY.OchiaiK.WatanabeM.AbarzuaF.SakaguchiM.TakaokaM.TanimotoR.NasuY.HuhN.H. and KumonH., Down-regulation of inhibition of differentiation-1 via activation of activating transcription factor 3 and Smad regulates REIC/Dickkopf-3-induced apoptosis, Cancer Res68 (2008), 8333–8341.
36.
FongD.HermannM.UntergasserG.PirkebnerD.DraxlA.HeitzM.MoserP.MargreiterR.HengsterP. and AmbergerA., Dkk-3 expression in the tumor endothelium: A novel prognostic marker of pancreatic adenocarcinomas, Cancer Sci100 (2009), 1414–1420.
37.
KataseN.GunduzM.BederL.GunduzE.LefeuvreM.HatipogluO.F.BorkoskyS.S.TamamuraR.TominagaS.YamanakaN.ShimizuK.NagaiN. and NagatsukaH., Deletion at Dickkopf (dkk)-3 locus (11p15.2) is related with lower lymph node metastasis and better prognosis in head and neck squamous cell carcinomas, Oncol Res17 (2008), 273–282.