Alterations and mutations of endo-lysosomal trafficking proteins have been associated with cancer progression. Identification and characterization of endo-lysosomal trafficking proteins in invasive cholangiocarcinoma (CCA) cells may benefit prognosis and drug design for CCA.
OBJECTIVE:
To identify and characterize endo-lysosomal trafficking proteins in invasive CCA.
METHODS:
A lysosomal-enriched fraction was isolated from a TNF- induced invasive CCA cell line (KKU-100) and uninduced control cells and protein identification was performed with nano-LC MS/MS. Novel lysosomal proteins that were upregulated in invasive CCA cells were validated by real-time RT-PCR. We selected Rab7 for further studies of protein level using western blotting and subcellular localization using immunofluorescence. The role of Rab7 in CCA invasion was determined by siRNA gene knockdown and matrigel transwell assay.
RESULTS:
Rab7 mRNA and protein were upregulated in invasive CCA cells compared with non-treated controls. Immunofluorescence studies demonstrated that Rab7 was expressed predominantly in invasive CCA cells and was localized in the cytoplasm and lysosomes. Suppression of Rab7 translation significantly inhibited TNF--induced cell invasion compared to non-treated control ( 0.044).
CONCLUSIONS:
Overexpression of Rab7 in CCA cells was associated with cell invasion, supporting Rab7 as a novel candidate for the development of diagnostic and therapeutic strategies for CCA.
Cholangiocarcinoma (CCA), a cancerous tumor of the bile duct, is a serious health problem in Southeast Asian countries including Thailand [1]. Low responsiveness of CCA to anticancer drugs in combination with a lack of reliable early diagnosis and prognosis are major obstacles to prevention and control [2]. In this regard, novel protein candidates have been identified and characterized for the development of reliable diagnostic strategies and anticancer chemotherapeutics. Endo-lysosomal associated proteins, especially endocytic trafficking proteins, are fascinating targets that are involved in cancer cell development, migration, invasion, and metastasis [3]. The Rab GTPase and associated proteins play important roles in the regulation of endocytic transport. Alteration or mutation of these proteins is associated with several human diseases, including the pathogenesis of cancer [4].
Increased expression of Rab5, Rab7, and Rab11 genes and proteins was detected in patients with oral squamous cell carcinoma (OSCC). Moreover, expression of these proteins was significantly elevated in cases of advanced OSCC with a poor survival rate [5]. Increased Rab25 mRNA level in ovarian and breast cancer patients correlated with a low survival rate, and overexpression of Rab25 in in vitro and in vivo models by transfection with pcDNA-Rab25 plasmid promoted proliferation, colony formation, and invasion of ovarian and breast cancer cells [6]. Conversely, down regulation of Rab38 was reported in melanoma primary tumors and metastases compared to melanocytes [7]. However, the functions of endocytic trafficking proteins including the Rab family have not been demonstrated in CCA and should be intensively explored.
We performed proteomics analysis of lysosomal proteins to identify significant targets involved in CCA cell invasion. Several Rab proteins were upregulated in TNF- induced invasive CCA cells, in particular Rab7, a member of the small GTPase superfamily that functions in trafficking of late endosomes to lysosomes [8]. Alterations of Rab7 have been introduced into several cancers to promote or inhibit functions such as cell proliferation, migration, invasion, and metastasis [9]. To validate Rab7 expression in CCA cell lines, mRNA and protein levels were analyzed with SYBR real-time RT-PCR and western blot analysis, respectively, and compared between TNF- induced CCA cells and non-treated controls. Subcellular localization of Rab7 and its co-localization with lysosomes was detected using immunofluorescence. Depletion of Rab7 expression using siRNA in combination with a matrigel transwell assay was performed to clarify the role of this protein in CCA cell invasion. Our findings suggest a role of Rab7 in the aggressive pathogenesis of CCA that might be a target in the development of effective strategies to prevent and control CCA.
Materials and methods
Induction of invasive CCA
KKU-100, a non-invasive cholangiocarcinoma cell line, was continuously cultured as described elsewhere [10]. In brief, KKU-100 was cultured in complete medium (Ham’s F12 (Gibco, Billing, MT) supplemented with 10% fetal bovine serum (FBS, Biowest, France) and 1 penicillin/streptomycin (Biowest) at 37C, 5% CO, and N balance. Invasiveness of KKU-100 cells was stimulated by treatment with 10 ng/ml TNF- (ProSpec, East Brunswick, NJ) as described previously [11]. The cells were harvested by the addition of cold EDTA solution (10 mM EDTA in 1 PBS) and gentle detachment using a cell scraper (SPL Life Science, Gyeonggi-do, Korea).
Isolation of lysosomes
Lysosomes of TNF- treated and non-treated KKU-100 cells were isolated using a Lysosome Enrichment Kit for Tissue and Cultured Cells according to the manufacturer’s instructions (Thermo Fisher Scientific, Waltham, MA). After removal of OptiPrep media by centrifugation, lysosomes were mixed with 2% CHAPS (Sigma, St. Louis, MO) in 1 PBS for 10 min and then sonicated using a sonicator (Heat System, Farmingdale, NY). Soluble lysosomal proteins were obtained by centrifugation at 18,000 g and 4C for 5 min and subsequently concentrated using an Amicon Ultra-0.5 centrifugal filter device (Merck, Darmstadt, Germany) according to the manufacturer’s instructions. Protein concentration was determined using Coomassie Plus Protein (Bradford) assay reagent (Thermo Fisher Scientific, Pittsburgh, PA). The lysosomal-enriched fraction (LYE) was confirmed by western blot analysis of lysosomal-associated membrane protein 1 (LAMP1) using anti-LAMP1 antibody (Millipore, Billerica, MA) [12].
Primers used in SYBR real-time RT-PCR for determination of gene expression profile
Gene
Accession No.
Product size (bp)
Primer (5’-3’)
Primer length (nt)
Rab7
X93499.1
270
5’-AAGCCACAATAGGAGCTGAC-3’
20
5’-CAATCTTGTTTCCCAACACA-3’
20
Rab11B
NM_004218.3
260
5’-AACGAGTTCAACCTGGAGAG-3’
20
5’-ATGATGACGATGTTGCTGTC-3’
20
CATD
M11233.1
110
5’-GACCAGAACATCTTCTCCTTCTAC-3’
24
5’TAGGACAGAGAACCCTTGTAATACT-3’
25
GNA13
L22075.1
143
5’-AAGGGTTTTCTTACAATATCTTCCT-3’
25
5’-GTTCTCCAAGTTTATCCAAGTTATC-3’
25
Mass spectrometry
Extracted lysosomal proteins (1 g/l) from TNF- treated and non-treated KKU-100 cells were separated by 12% SDS-PAGE and stained with Coomassie brilliant blue G250 solution (BioRad, Berkeley, CA). Lanes of gels containing proteins were excised into equal small cubes (Fig. 1B) and subjected to downstream processing including alkylation, tryptic digestion, mass spectrometry analysis (MicroToFQ II, Bruker, Germany) coupled with an UltiMate 3000 nano-LC system (Dionex, Surrey, UK), and protein identification using bioinformatics as described previously [11, 13]. Novel lysosomal proteins that were overexpressed in TNF- induced invasive cells were chosen and computationally analyzed by protein – protein interaction analysis using STRING database version 10 pathway analysis [14] and the KEGG pathway website [15]. Selected candidates were further validated using molecular biology methods as described below.
SYBR real-time RT-PCR
Total RNA of TNF- treated and non-treated KKU-100 cells was isolated using Trizol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer’s instructions. Five micrograms of total RNA was incubated with DNase I (Thermo Fisher Science, Pittsburgh, PA) to remove genomic DNA contaminants and then converted to first-strand cDNA using RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific) according to the manufacturer’s instructions. Forward (Fwd) and reverse (Rev) primers were designed using Primer3 version 0.4.0 program [16] and are described in Table 1). To determine the level of gene expression, SYBR green real-time RT-PCR was performed by mixing 2 l of first-strand cDNA with 1 SsoAdvanced SYBR Green Supermix (BioRad, Hercules, CA) and 500 nM each of Fwd and Rev primers in a total volume of 15 l. Amplification was performed using a LightCycler 480 version 1.5 (Roche, Basel, Switzerland) with amplification conditions of 95C for 3 min, followed by 45 cycles of 95C for 30 s and 60C for 10 s. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as a housekeeping gene for normalization [17]. The relative fold change of gene targets in TNF--treated cells compared to non-treated controls was calculated using the formula of 2 [18]. The experiment was performed in duplicate in three independent experiments.
Western blot analysis
The results of SYBR real-time RT-PCR and mass spectrometry indicated that Rab7 was markedly overexpressed in TNF- induced invasive KKU-100 cells compared with other proteins. Therefore, Rab7 was selected for further confirmation of the level of protein expression using western blot analysis. TNF- treated and non-treated KKU-100 cells were mixed with RIPA buffer (50 mM Tris-HCl, 150 mM NaCl, 1% NP-40, 0.1% SDS, and 1 mM EDTA) and disrupted using a sonicator with three pulses of 9.0 s on and 9.0 s off. Protein concentration was determined with Coomassie Plus Protein assay reagent (Thermo Fisher Science) prior to size separation by 12% SDS-PAGE. The proteins in the gel were electrically transferred onto PVDF membranes (Pall, Washington, NY) and subjected to western blot analysis as described elsewhere [11, 13] using mouse anti-human Rab7 antibody (Abcam, Cambridge, MA) and mouse anti-human -actin antibody (Cell Signaling Technology, Danvers, MA) as a control. The signal was detected using ChemiDoc Imager (Syngene, Frederick, MD) and the intensity was calculated with ImageJ program (http://imagej.nih.gov/ij/). The experiment was performed in triplicate.
Immunolocalization
Detection of Rab7 in TNF- treated and non-treated KKU-100 cells was performed by an immunofluorescent technique. The co-localization of Rab7 with lysosomes (using LAMP1 as a marker) was also monitored. All procedures were performed as previously described with minor modification [11, 13]. Briefly, the cells were permeabilized by incubation with 0.25% Tween-20 for 10 min at RT and blocked with non-specific binding with blocking solution (3% FBS in 1 PBS) for 1 h. Mouse anti-human Rab7 antibody (1:500 dilution; Abcam) and rabbit anti-human LAMP1 antibody (1:500; Millipore, Temecula, CA) were added and incubated for 2 h at RT. FITC-conjugated goat anti-mouse IgG (1:500; Biolegend, San Diego, CA) and cy3-conjugated donkey anti-rabbit IgG (1:2,000; Biolegend) were then added and incubated with the cells for 1 h at RT in a dark moist chamber. Nuclei were counterstained with 1 g/ml 4’,6-diamidino-2-phenylinodole (DAPI; Sigma-Aldrich) for 5 min. Fluorescence imaging was performed using a LSM700 confocal microscope (Carl Zeiss, Oberkochen, Germany). The intensity of Rab7 expressed in the cells was calculated with ImageJ program (http://imagej.nih.gov/ij/).
siRNA knockdown
To investigate the role of Rab7 in CCA cell lines we performed gene knockdown using a small interference RNA (siRNA) approach. Sense [5’ GUC UAGUUCCC UUCUGUGU(dTdT) 3’] and antisense [5’ACACA GAAGGGAACUAGAC (dTdT) 3’] oligonucleotides specific for Rab7 were purchased from BIONEER Company (Bioneer, Daejeon, South Korea). Transfection procedures were performed according to the manufacturer’s instructions (Bioneer). Briefly, KKU-100 cells were grown until 60–80% confluent, seeded into 24-well plates at a density of 1 10 cells/well, and incubated at 37C, 5% CO for 24 h. The siRNA duplex-LipofectamineTM RNAimax complex was prepared by mixing Lipofectamine (Invitrogen) with 20 nM siRNA duplex (Rab7 or GAPDH as negative control) and added to KKU-100 cells. After incubation at 37C, 5% CO for 5–6 h, the medium was replaced with OptiMEM without antibiotics and the cells were incubated for a further 24 h. The cells were harvested by addition of 500 l Trizol reagent for isolation of total RNA and Rab7 expression level was determined using SYBR green real-time RT-PCR as described above. The experiment was performed in triplicate in two independent experiments.
Cell invasion assay
The matrigel invasion assay was modified from the science advisory board protocol (http//www.scienceboard.net/resources/protocols) to investigate the effect of Rab7 protein knockdown on the invasiveness of KKU-100 cells. The transwells were filled with matrix gel at 1 mg/ml and incubated for 4–5 hours at 37C. KKU-100 cells (1 10 cells/well) were seeded on top of the matrix gel under four conditions: without TNF- as a cell control, with 10 ng/ml TNF- and without siRNA to Rab7 as a positive control, with 10 ng/ml TNF- and 20 nM siRNA to Rab7; and with 10 ng/ml TNF- and Lipofectamine only as a negative control. Cells were incubated at 37C with or without siRNA to Rab7 and then treated with or without 10 ng/ml TNF- for 1 h depending on the treatment group. After treatment, 650 l of OptiMEM medium without antibiotics was added to all transwells, and the cells were incubated at 37C, 5% CO for 24 h. The transwells were washed with 1 PBS (pH 7.4) two times and the cells were fixed with 3.7% formaldehyde for 2 min and permeabilized by incubation with 100% methanol for 20 min at room temperature. Cells were washed with 1 PBS (pH 7.4), stained with Giemsa (Sigma) for 15 min in the dark, washed again with 1 PBS (pH 7.4), and the non-invading cells were gently removed with a cotton swab. The plates were left at room temperature until dry and then the transwell membrane was removed and placed upside down on 90% glycerol before monitoring of KKU-100 cell invasion under the microscope (Olympus, Bulverde, TX).
Data analysis
Mann-Whitney test was used for analysis of the fluorescence intensity of Rab7 expressed in cells and paired t-test was used for analysis of the invasion inhibition assay. Both analyses were significant at a -value 0.05.
Results
Isolation of lysosomal enriched fraction from CCA cell line
KKU-100 cells were incubated with 10 ng/ml TNF- for 24 h to induce invasive characteristics as previously described by our group [11]. Lysosomes from TNF- treated and non-treated KKU-100 cells were isolated using a gradient centrifugation technique [19]. Western blot analysis of the lysosomal marker LAMP-1 confirmed that LAMP-1 was predominantly detected in the LYE of both TNF- treated and non-treated KKU-100 cells at a molecular weight of 95 kDa (Fig. 1A). LAMP-1 was also detected in other subcellular fractions and the wash supernatant but at a lower level compared to the LYE. The LYE from TNF- treated and non-treated KKU-100 cells was separated by 12% SDS-PAGE and the lanes of the gel were cut into 10 equal small cubes prior to protein identification with MicroToFQ II (Fig. 1B).
Analysis of lysosomal enriched proteins. (A) Western blot analysis of isolated lysosomal enriched fraction using anti-LAMP1 antibody. LYE: Lysosome enriched, OSC: Other sub-cellular, WSP: Wash supernatant. LAMP1 was detected at 95 kDa (arrow head). (B) The lysosomal enriched fraction was separated by 12% SDS-PAGE and the lane of the gel was excised into 10 equal sized.
Lysosomal protein profiles of TNF- induced invasive CCA cell line
Mass spectrometry combined with bioinformatics analysis identified a total of 78 proteins from the lysosomal enriched fraction of both TNF- treated and non-treated KKU-100 cells. Thirty-five novel upregulated proteins were detected in TNF- treated KKU-100 cells. Analysis of molecular function, biological process, cellular component, and protein class using the PANTHERclassification system suggested that 13 different proteins were associated with the lysosome and/or endosome, as presented in Table 2. Mean emPAI value and ratio with standard deviation (S.D) were calculated to compare the expression level of endosomal-lysosomal proteins. The interactive protein network expressed in TNF- induced invasive CCA cells was presented through a string pathway (Fig. 2). Several proteins belonging to the Rab family were identified: Rab7, Rab9, Rab11, Rab13, Rab35, RAP1A, RRAS, GNA13, and CFL1. These proteins were shown to interact with other protein groups; for example, proteins related to cytoskeleton organization such as CDC42, CD44, TUBA1A, and ACTB were demonstrated to have a close relationship with the Rab-family.
Upregulated endo-lysosomal proteins in TNF- induced invasive CCA cells compared with non-treated controls
Accession no.
Prot. name
Subcellular location
Mass
emPAI value
emPAI ratio
Control
TNF-
1. gi809185
Annexin V
Early and late endosome
35,783
0.22 0.05
2.01 0.25
9.14
2. gi4757756
Annexin A2 isoform2
Lysosome membrane
38,580
0.59 0.14
3.02 0.31
5.12
Early/late endosome
3. gi4503143
Cathepsin D
Lysosome
44,524
0.08 0.02
0.18 0.04
2.25
4. gi291360740
MHC class I Ag
Early endosome membrane
21,021
0.18 0.08
0.4 0.08
2.22
5. gi1174149
Small GTP binding protein Rab7
Lysosome membrane
23,447
0.16 0.04
0.35 0.07
2.19
Late endosome
6. gi763130
Ras-related protein Rab11B
Endosome
24,559
0.16 0.04
0.33 0.04
2.06
7. gi334849548
MHC class I Ag
Early endosome membrane
39,295
0.1 0.02
0.2 0.01
2
8. gi374843690
MHC class I Ag, partial
Early endosome membrane
35,565
0.35 0.09
0.57 0.11
1.63
9. gi410110995
MHC class I Ag, partial
Early endosome membrane
31,571
0.25 0.12
0.4 0.06
1.60
10. gi231367
HLA class I Ag A-31 chain
Early endosome membrane
40,978
ND*
0.42 0.05
NC*
11. gi4759012
Ras-related protein Rab9A
Lysosome/late endosome
22,823
ND*
0.17 0.02
NC*
Phagosome membrane
12. gi5803135
Ras-related protein Rab35
Endosome membrane
23,011
ND*
0.17 0.07
NC*
13. gi404722
Guanine nucleotide regulatory protein
Lysosome membrane
44,036
ND*
0.08 0.00
NC*
emPAI exponentially modified protein abundance index. ND Non detectable, NC Not calculated.
Functional protein association by string network. Balls show protein alterations; dark grey balls show members of the Rab protein family. RAB7A: Ras-related protein Rab7, RAB9A: Ras-related protein Rab9, RAB11B: Ras-related protein Rab-11B, RAB13: Ras-related protein Rab13, RAB35: Ras-related protein Rab35, RAP1A: Ras-related protein Rap-1A, GNA13: Guanine nucleotide-binding protein subunit alpha-13, CFL1: Cofilin-1, RRAS: Ras-related protein R-Ras.
Upregulation of lysosomal proteins in TNF- induced CCA cell line
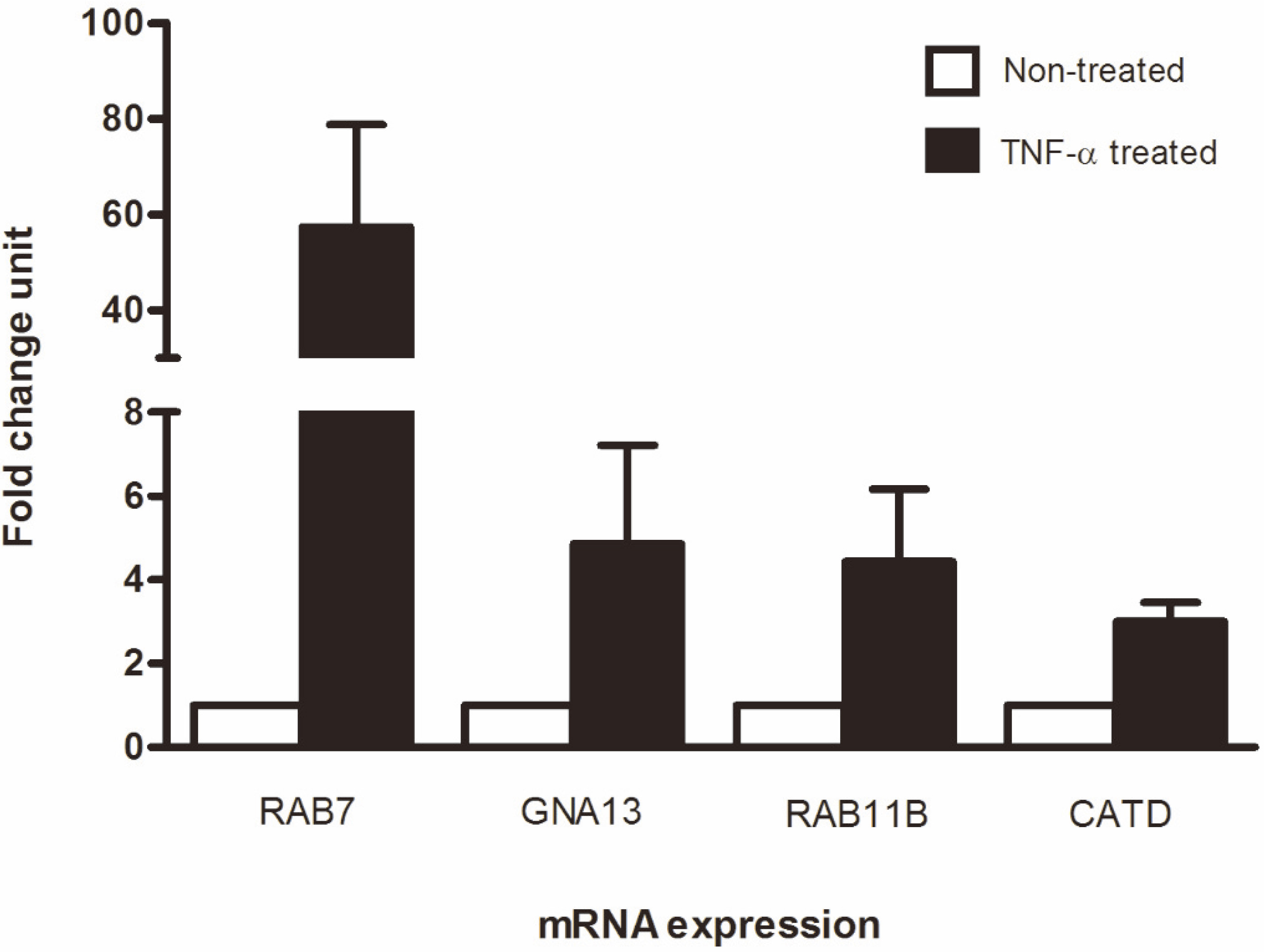
Among 13 lysosomal protein candidates, six proteins – Ras-related protein Rab-9A (Rab9A), Ras-related protein Rab-35 (Rab35), small GTP binding protein Rab7 (Rab7), Ras-related Rab11B (Rab11B), cathepsin D (CATD), and guanine nucleotide regulatory protein (GNA13) – were selected for further validation using SYBR real-time RT-PCR based on the following criteria: (1) More than 2-fold up regulation in TNF- treated KKU-100 cells compared to non-treated control, (2) present in TNF- treated KKU-100 cells but not controls, (3) no or little available information regarding a role in CCA pathogenesis. However, SYBR real-time PCR of Rab9A and Rab35 gave negative results, in contrast to the results of mass spectrometry. As a result, only four proteins (Rab7, Rab11B, CATD, and GNA13) that showed consistent findings between mass spectrometry and SYBR real-time RT-PCR were selected as candidates. Interestingly, Rab7 mRNA was markedly expressed in TNF- treated KKU-100 cells with a fold change of 57.43 compared with non-treated controls. GNA13, Rab11B, and CATD were also upregulated in TNF- treated KKU-100 with fold change units of 4.86, 4.43, and 3.01, respectively (Fig. 3). Rab7 therefore showed massive upregulation more than 10 times greater than that of the other candidates, and was selected for further investigation of the protein expression level by western blotting. Comparison of Rab7 expression in TNF- treated and non-treated KKU-100 cells suggested that protein expression was increased approximately 5-fold during TNF- induced cell invasiveness relative to controls (Fig. 4A and B). These data suggest a pivotal role of Rab7 in invasiveness and/or other pathogenesis-related processes of CCA.
SYBR real-time RT-PCR for validation of mRNA expression. Fold change unit was calculated by comparison of TNF- treated and non-treated cells. GAPDH was used as a control to normalize the expression level of gene targets. RAB7: Ras-related protein Rab7, GNA13: guanine nucleotide regulatory protein, RAB11B: Ras-related protein Rab11B, CATD: Cathepsin D.
Western blot analysis of Rab7 using anti-human Rab7 antibody shows an increase in Rab7 protein in TNF- induced CCA cells (A) with calculated fold change compared with non-treated control (B). Immunolocalization of Rab7 in CCA cells demonstrated that Rab7 was overexpressed in TNF- induced invasive CCA cells and was localized in the cytoplasm and lysosomes (C). Co-localization of Rab7 (green) with LAMP1 (red) was indicated in the merged image (yellow). Green, anti-human Rab7; red, anti-human LAMP1; blue, DAPI bound to DNA. Mean fluorescence intensity of Rab7 expressed in CCA cells; p-value 0.05; 0.0119, 0.0119, NS Not significant (D).
Rab7 localized with lysosomes
The intracellular localization of Rab7 was examined using immunofluorescence to determine the location of this protein in subcellular organelles. We found that the increased Rab7 expression in TNF- treated KKU-100 cells was predominantly localized in the cytoplasm. In previous studies, Rab7 was found on the endosomal/lysosomal membrane where it facilitates trafficking of endosomes to lysosomes [20]. To investigate the lysosomal localization of Rab7, we examined co-immunofluorescence staining of Rab7 and LAMP-1 and found that Rab7 was co-localized with LAMP-1, supporting its lysosomal association (Fig. 4C). An expression level of Rab7 in KKU-100 cells was determined by detection of the fluorescence intensity using ImageJ program. The result demonstrated that the intensity of Rab7 expressed in TNF- treated KKU-100 cells was extremely high but rarely observed in non-treated control (-value 0.0119; Fig. 4D).
Rab7 knockdown affected invasion of TNF- induced CCA cells
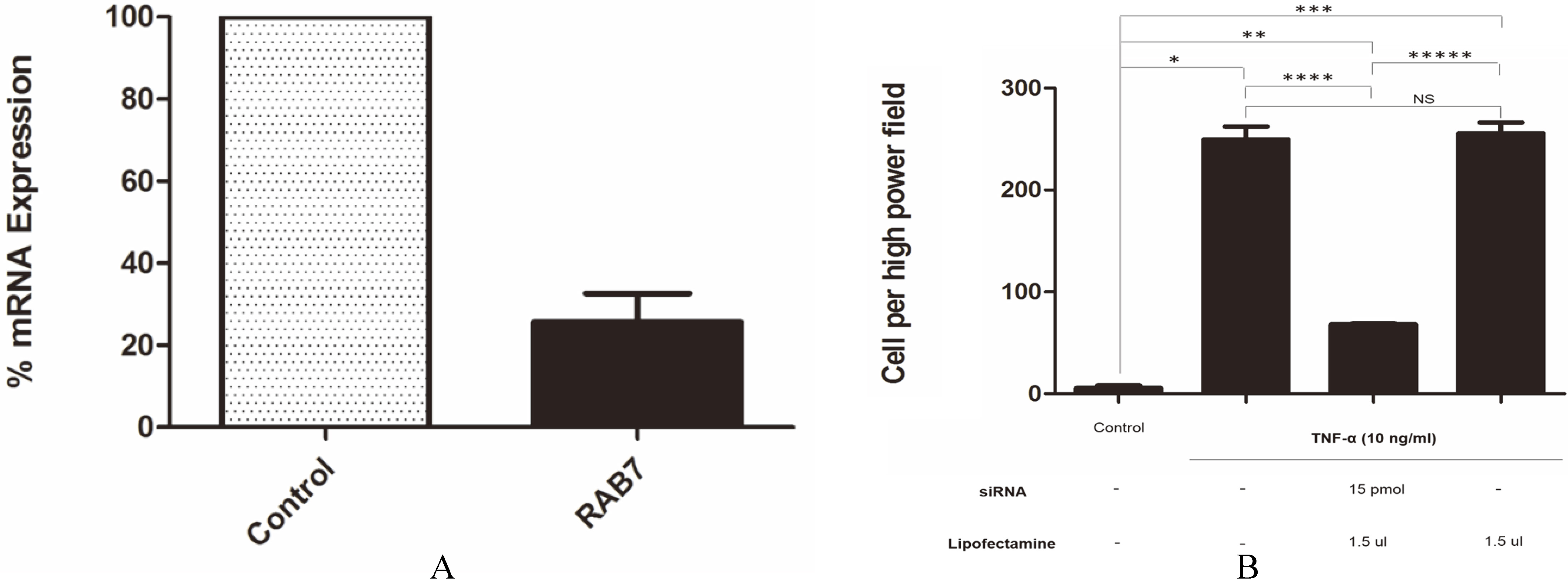
The role of Rab7 in cell migration and invasion of CCA has not been previously described. Therefore, specific siRNA against Rab7 was introduced into CCA cells to inhibit gene expression. TNF- induced KKU-100 cells transfected with siRNA against Rab7 exhibited an approximately 80% decrease in Rab7 mRNA expression level compared to cells transfected with siRNA control (Fig. 5A). To prove the hypothesis that Rab7 is associated with invasion of KKU-100 cells, we performed a matrigel invasion assay in combination with Rab7 knockdown. siRNA against Rab7 inhibited cell invasion after TNF- treatment compared with siRNA control (-value 0.044; Fig. 5B).
Knockdown of Rab7 expression using siRNA. (A) Approximately 80% suppression of Rab7 expression was achieved with Rab7 siRNA compared with control siRNA. (B) Rab7 siRNA impaired CCA cell invasion as analyzed by matrigel invasion assay; -value 0.05; 0.026, 0.025, 0.033, 0.044 and 0.036, NS Not significant. The experiment was performed in triplicate.
Discussion
Treatment of CCA has recently become an area of concern because of its low responsiveness to available anticancer drugs [21]. Trials of alternative drugs and supportive therapies, as well as identification of novel drug targets, may provide novel insights into this cancer. Endo-lysosomal associated proteins, especially lysosomal trafficking proteins and proteases, are fascinating candidates for the development of anticancer therapeutics [22]. In prostate cancer, blocking anterograde lysosome trafficking using niclosamide, a human anti-helminthic drug, resulted in a significant decrease in tumor cell invasion [23]. Treatment of glioma with the selective lysosome lysing drug glycyl-L-phenylalanine-ß-naphthylamide (GPN) or the lysosome exocytosis inhibitor vacuolin-1 significantly inhibited cancer cell migration and invasion [24].
Lysosomal proteins associated with the invasion of CCA were identified in this study using a proteomics approach. LYE was isolated from TNF- induced invasive KKU-100 cells and the controls using a gradient centrifugation technique and confirmed by detection of LAMP1. After analysis by mass spectrometry, six lysosomal proteins – Rab7, Rab9A, Rab11B, Rab35, CATD, and GNA13 – were selected for further validation using SYBR real-time PCR. Annexin V and A2 isoform 2 were remarkably upregulated with the highest emPAI ratios of 9.14 and 5.12, respectively; however, these two annexins were not selected for further study because they were already known to be associated with CCA development [25, 26]. The MHC class I family is another group that was omitted from this study because of available data on its role in cancer [27].
Small GTP binding proteins Rab7, Rab9A, Rab11B, and Rab35 play an important role in endocytosis and intracellular movement of proteins; Rab7 is localized on lysosomal membranes and late endosomes and drives vesicle fusion to the lysosome [28], whereas Rab9A, Rab11B, and Rab35 are presented on endosomes and play a role in recycling proteins to the trans golgi network and the plasma membrane [29, 30]. CATD is a lysosomal aspartic protease that is involved in protein turnover and activation of hormones and growth factors [31]. Lastly, GNA13 plays important roles in transduction and modulation in various transmembrane signaling systems as well as cell division [32].
The transcription level of all six selected candidates was compared between TNF- treated KKU-100 cells and non-treated controls using SYBR real time RT-PCR. Rab9A and Rab35 genes could not be detected despite changing the primer sets and several repeats of the experiment. Therefore, Rab9A and Rab35 were discarded from this study. The transcription levels of Rab7, Rab11B, CATD, and GNA13 were upregulated during CCA cell invasion. In thyroid adenomas, overexpression of Rab5A and Rab7 was observed in cancerous tissue with a 6-fold increase compared with surrounding quiescent tissue [33]. Comparison of gene expression signatures between ovarian/primary peritoneal serous carcinoma (OC/PPC) and diffuse peritoneal malignant mesothelioma (DMPM) showed that Rab7 was upregulated in OC/PPC whereas Rab25 was upregulated in DMPM [34]. In colorectal cancer, an increase in Rab11B expression level was associated with cancer cell migration through regulation of E-cadherin [35]. An increase in secreted CATD was observed in most metastatic breast cancer cell lines in response to estrogen, growth factors, or unknown mechanisms [36]. An important role of GNA13 in inducing invasive characteristics in prostate and breast cancers was also reported [37, 38]. Among the four proteins, Rab7 was predominantly transcribed in a TNF- induced invasive CCA cell line compared with the control. Moreover, upregulation of Rab7 transcript in the TNF- treated CCA cell line was approximately 10 times greater than that of other candidate genes under the same conditions. These findings imply an important role of Rab7 in the invasiveness of CCA cell lines after TNF- induction. In this regard, Rab7 was selected for further analysis and its expression was confirmed at the protein level using western blot analysis. The high expression level of Rab7 protein in the TNF- induced CCA cell line seems to emphasize its association with CCA cell invasion. Rab7 is a small GTP binding protein that is stimulated by catalysis of the inactive-GDP bound form to the active-GTP bound form. GTP-bound Rab7 can recruit several downstream effectors to facilitate different functions of membrane transportation [28], including a role in endo-lysosomal trafficking that mediates endosomal maturation, transportation of vesicles from a late endosome to a lysosome via the cytoskeleton (plus or minus end), and biogenesis [8, 39].
In the CCA cell line, Rab7 protein was located in the cytoplasm and was also co-localized with lysosomes, consistent with reports in other cancers [40]. Overexpression of Rab7 in the cytoplasm and lysosomes of TNF- treated KKU-100 cells supports its role in cell invasion as well as other processes associated with cancer progression. In lung cancer, up-regulation of Rab7 was detected in invasive and metastatic tumors compared to normal lung tissue [41]. Moreover, recent follow-up data of patients with metastatic oral squamous cell carcinoma (OSCC) over 10 years suggested predominant amplification of Rab gene family including Rab7 in chromosomal regions [5]. However, up regulation of Rab7 in progressive clinical CCA samples has not yet been investigated and this should be a priority of future studies.
As mentioned above, Rab7 was associated with invasive and progressive characteristics of CCA and other cancers. Blockage of Rab7 and other Rab family members in a variety of cancers interfered with cell migration, invasion, and metastatic characteristics [42]. Knockdown of Rab7 expression in a CCA cell line through siRNA silencing resulted in a significant decrease in cell invasion after induction with TNF-. Our results were consistent with those of previous studies regarding the role of Rab7 in tumor invasion and metastasis. It has been shown that suppression of Rab7 expression in skin cancer (A431) and breast cancer (MCF7) cell lines significantly down regulated EGFR and HER2 levels, which may affect Akt signaling pathway-mediated cell survival and invasion [43]. However, a controversial outcome of Rab7 down regulation on cancer invasion has been reported in several cancers. For example, depletion of Rab7 expression in melanoma and prostate cancer using shRNA induced cancer cell invasion [44, 45]. Such discrepancies in the apparent role of Rab7 in cancer cell invasion may reflect the origin of the cancerous tissue. The mechanism of action of Rab7 in CCA cell invasion was addressed by modeling in the string pathway, which demonstrated that a partner protein predominantly involved in cytoskeleton organization interacts with Rab7 and other Rab proteins in the TNF- induced invasive CCA cell line. These findings may provide a good explanation for Rab7 function in migration and invasion of the CCA cell line. In this regard, cofilin-1 (CFL1) is an interesting protein that was upregulated during TNF- induced CCA cell invasion. It belongs to a family of small actin binding proteins that have an important role in depolymerization, cell morphology, and cytoskeletal management [46]. Major expression of the cofilin pathway has been described in invasive and metastatic breast cancer [47]. In addition to CFL1, cell division control protein 42 (CDC42) was also identified in the invasive CCA cell line. This protein associates with CFL1 to activate actin cytoskeleton motility in the cell [48] and promotes cell movement via filopodia cell formation. Moreover, overexpression of CDC42 has been reported in some breast cancers [49]. CD44 is another fascinating protein that was present in our string pathway and correlates with tumor cell invasiveness and metastasis through the phenomenon of lamellipodia cell transformation [50]. Based on the information above, most of the proteins that interplay with Rab7 in invasive CCA cells were related to cancer cell migration via transformation of cell cytoskeleton. Nonetheless, the interaction of lysosomal trafficking proteins such as Rab7 with other migration and invasion associated proteins in vitro and in vivo should be investigated in the near future.
In summary, overexpression of the endo-lysosomal trafficking protein Rab7 was identified in invasive CCA cell lines using a proteomics approach. Silencing of this gene suggested its important role in CCA cell invasion. Our findings may advance the development of anticancer drugs and prognostic markers for CCA.
Conflict of interest
The authors declare no conflict of interest.
Footnotes
Acknowledgments
This work is supported by a grant from the Thailand Research Fund (TRF) in the Program of Initiative Career Development to Assistant Professor Dr. Poom Adisakwattana (MRG5480127) and a RA scholarship from the Faculty of Graduate Studies and Faculty of Tropical Medicine, Mahidol University, Thailand, to Ms. Nantana Suwandittakul. We thank the Faculty of Tropical Medicine for supporting the budget for an English editing service.
References
1.
SripaB. and PairojkulC., Cholangiocarcinoma: lessons from Thailand, Curr Opin Gastroenterol24 (2008), 349–56.
MosessonY.MillsG.B. and YardenY., Derailed endocytosis: an emerging feature of cancer, Nat Rev Cancer8 (2008).
4.
SteinM.P.DongJ. and Wandinger-NessA., Rab proteins and endocytic trafficking: potential targets for therapeutic intervention, Adv Drug Deliv Rev55 (2003), 1421–1437.
5.
da SilvaS.D.MarchiF.A.XuB.BijianK.AlobaidF.MlynarekA.RogattoS.R.HierM.KowalskiL.P. and Alaoui-JamaliM.A., Predominant Rab-GTPase amplicons contributing to oral squamous cell carcinoma progression to metastasis, Oncotarget6 (2015), 21950–21963.
6.
ChengK.W.LahadJ.P.KuoW.L.LapukA.YamadaK.AuerspergN.LiuJ.Smith-McCuneK.LuK.H.FishmanD.GrayJ.W. and MillsG.B., The RAB25 small GTPase determines aggressiveness of ovarian and breast cancers, Nat Med10 (2004).
7.
MuellerD.W.RehliM. and BosserhoffA.K., miRNA expression profiling in melanocytes and melanoma cell lines reveals miRNAs associated with formation and progression of malignant melanoma, J Invest Dermatol129 (2009), 1740–1751.
8.
FengY.PressB. and Wandinger-NessA., Rab 7: an important regulator of late endocytic membrane traffic, J Cell Biol131 (1995), 1435–1452.
9.
RecchiC. and SeabraM.C., Novel functions for Rab GTPases in multiple aspects of tumour progression, Biochem Soc Trans40 (2012), 1398–403.
10.
SripaB.LeungwattanawanitS.NittaT.WongkhamC.BhudhisawasdiV.PuapairojA.SripaC. and MiwaM., Establishment and characterization of an opisthorchiasis-associated cholangiocarcinoma cell line (KKU-100), World J Gastroenterol11 (2005), 3392–3397.
11.
AdisakwattanaP.SuwandittakulN.PetmitrS.WongkhamS.SangvanichP. and ReamtongO., ALCAM is a novel cytoplasmic membrane protein in TNF-α stimulated invasive cholangiocarcinoma cells, Asian Pac J Cancer Prev16 (2015), 3849–3856.
12.
KornfeldS. and MellmanI., The biogenesis of lysosomes, Annu Rev Cell Biol5 (1989), 483–525.
13.
MoleeP.AdisakwattanaP.ReamtongO.PetmitrS.SricharunratT.SuwandittakulN. and ChaisriU., Up-regulation of AKAP13 and MAGT1 on cytoplasmic membrane in progressive hepatocellular carcinoma: a novel target for prognosis, Int J Clin Exp Pathol8 (2015), 9796–9811.
14.
SzklarczykD.FranceschiniA.WyderS.ForslundK.HellerD.Huerta-CepasJ.SimonovicM.RothA.SantosA.TsafouK.P.KuhnM.BorkP.JensenL.J. and von MeringC., STRING v10: protein-protein interaction networks, integrated over the tree of life, Nucleic Acids Res43 (2015), D447–D452.
15.
KanehisaM., Pathway databases and higher order function, Adv Protein Chem54 (2000), 381–408.
16.
UntergasserA.NijveenH.RaoX.BisselingT.GeurtsR. and LeunissenJ.A., Primer3Plus, an enhanced web interface to Primer3, Nucleic Acids Res35 (2007), W71–W74.
17.
DhedaK.HuggettJ.F.BustinS.A.JohnsonM.A.RookG. and ZumlaA., Validation of housekeeping genes for normalizing RNA expression in real-time PCR, Biotechniques37 (2004), 112–114, 116, 118–119.
18.
LivakK.J. and SchmittgenT.D., Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method, Methods25 (2001), 402–408.
ZhangM.ChenL.WangS. and WangT., Rab7: Roles in membrane trafficking and disease, Biosci Rep29 (2009), 193–209.
21.
Ramírez-MerinoN.AixS.P. and Cort’es-FunesH., Chemotherapy for cholangiocarcinoma: An update, World Journal of Gastrointestinal Oncology5 (2013), 171–176.
22.
FehrenbacherN. and JaattelaM., Lysosomes as targets for cancer therapy, Cancer Res65 (2005), 2993–2995.
23.
CircuM.L.DykesS.S.CarrollJ.KellyK.GalianoF.GreerA.CardelliJ. and El-OstaH., A novel high content imaging-based screen identifies the anti-helminthic niclosamide as an inhibitor of lysosome anterograde trafficking and prostate cancer cell invasion, PLoS One11 (2016), e0146931.
24.
LiuY.ZhouY. and ZhuK., Inhibition of glioma cell lysosome exocytosis inhibits glioma invasion, PLoS One7 (2012), e45910.
25.
SrisomsapC.SawangareetrakulP.SubhasitanontP.ChokchaichamnankitD.ChiablaemK.BhudhisawasdiV.WongkhamS. and SvastiJ., Proteomic studies of cholangiocarcinoma and hepatocellular carcinoma cell secretomes, J Biomed Biotechnol2010 (2010), 437143.
26.
YonglitthipagonP.PairojkulC.ChamgramolY.MulvennaJ. and SripaB., Up-regulation of annexin A2 in cholangiocarcinoma caused by Opisthorchis viverrini and its implication as a prognostic marker, Int J Parasitol40 (2010), 1203–1212.
27.
GoeppertB.FrauenschuhL.ZucknickM.RoesslerS.MehrabiA.HafeziM.StenzingerA.WarthA.PathilA.RennerM.SchirmacherP. and WeichertW., Major histocompatibility complex class I expression impacts on patient survival and type and density of immune cells in biliary tract cancer, Br J Cancer113 (2015), 1343–1349.
28.
HutagalungA.H. and NovickP.J., Role of Rab GTPases in membrane traffic and cell physiology, Physiol Rev91 (2011), 119–149.
29.
BarberoP.BittovaL. and PfefferS.R., Visualization of Rab9-mediated vesicle transport from endosomes to the trans-Golgi in living cells, J Cell Biol156 (2002), 511–518.
30.
JingJ. and PrekerisR., Polarized endocytic transport: the roles of Rab11 and Rab11-FIPs in regulating cell polarity, Histol Histopathol24 (2009), 1171–1180.
31.
WoessnerJ.F., Jr. and ShambergerR.J., Jr., Purification and properties of cathepsin D from bovine utrus, J Biol Chem246 (1971), 1951–1960.
32.
SimonM.I.StrathmannM.P. and GautamN., Diversity of G proteins in signal transduction, Science252 (1991), 802–808.
33.
Croizet-BergerK.DaumerieC.CouvreurM.CourtoyP.J. and van den HoveM.F., The endocytic catalysts, Rab5a and Rab7, are tandem regulators of thyroid hormone production, Proc Natl Acad Sci U S A99 (2002), 8277–8282.
34.
DavidsonB.ZhangZ.KleinbergL.LiM.FlorenesV.A.WangT.L. and Shih IeM., Gene expression signatures differentiate ovarian/peritoneal serous carcinoma from diffuse malignant peritoneal mesothelioma, Clin Cancer Res12 (2006), 5944–5950.
35.
ChungY.C.WeiW.C.HuangS.H.ShihC.M.HsuC.P.ChangK.J. and ChaoW.T., Rab11 regulates E-cadherin expression and induces cell transformation in colorectal carcinoma, BMC Cancer14 (2014), 587.
36.
GarciaM.PlatetN.LiaudetE.LaurentV.DerocqD.BrouilletJ.P. and RochefortH., Biological and clinical significance of cathepsin D in breast cancer metastasis, Stem Cells14 (1996), 642–650.
37.
RasheedS.A.TeoC.R.BeillardE.J.VoorhoeveP.M. and CaseyP.J., MicroRNA-182 and microRNA-200a control G-protein subunit alpha-13 (GNA13) expression and cell invasion synergistically in prostate cancer cells, J Biol Chem288 (2013), 7986–7995.
38.
RasheedS.A.TeoC.R.BeillardE.J.VoorhoeveP.M.ZhouW.GhoshS. and CaseyP.J., MicroRNA-31 controls G protein alpha-13 (GNA13) expression and cell invasion in breast cancer cells, Mol Cancer14 (2015), 67.
39.
VitelliR.SantilloM.LatteroD.ChiarielloM.BifulcoM.BruniC.B. and BucciC., Role of the small GTPase Rab7 in the late endocytic pathway, J Biol Chem272 (1997), 4391–4397.
40.
StenmarkH., Rab GTPases as coordinators of vesicle traffic, Nat Rev Mol Cell Biol10 (2009), 513–525.
41.
NakanoT.ShimizuK.KawashimaO.KamiyoshiharaM.KakegawaS.SuganoM.IbeT.NagashimaT.KairaK.SunagaN.OhtakiY.AtsumiJ. and TakeyoshiI., Establishment of a human lung cancer cell line with high metastatic potential to multiple organs: gene expression associated with metastatic potential in human lung cancer, Oncol Rep28 (2012), 1727–1735.
42.
SubramaniD. and AlahariS.K., Integrin-mediated function of Rab GTPases in cancer progression, Mol Cancer9 (2010), 312.
43.
WangT.ZhangM.MaZ.GuoK.TergaonkarV.ZengQ. and HongW., A role of Rab7 in stabilizing EGFR-Her2 and in sustaining Akt survival signal, J Cell Physiol227 (2012), 2788–2797.
44.
Alonso-CurbeloD.Riveiro-FalkenbachE.Perez-GuijarroE.CifdalozM.KarrasP.OsterlohL.MegiasD.CanonE.CalvoT.G.OlmedaD.Gomez-LopezG.GranaO.Sanchez-Arevalo LoboV.J.PisanoD.G.WangH.W.Ortiz-RomeroP.TormoD.HoekK.Rodriguez-PeraltoJ.L.JoyceJ.A. and SoengasM.S., RAB7 controls melanoma progression by exploiting a lineage-specific wiring of the endolysosomal pathway, Cancer Cell26 (2014), 61–76.
45.
SteffanJ.J.DykesS.S.ColemanD.T.AdamsL.K.RogersD.CarrollJ.L.WilliamsB.J. and CardelliJ.A., Supporting a role for the GTPase Rab7 in prostate cancer progression, PLoS One9 (2014), e87882.
46.
GohlaA.BirkenfeldJ. and BokochG.M., Chronophin, a novel HAD-type serine protein phosphatase, regulates cofilin-dependent actin dynamics, Nat Cell Biol7 (2005), 21–29.
47.
WangW.EddyR. and CondeelisJ., The cofilin pathway in breast cancer invasion and metastasis, Nat Rev Cancer7 (2007), 429–440.
48.
KohC.-G. and LimL., Cdc42 and Rac control of the actin cytoskeleton, in: RHO Family GTPasesManserE., eds., Springer Netherlands, 2005, pp. 137–156.
49.
FritzG.JustI. and KainaB., Rho GTPases are over-expressed in human tumors, Int J Cancer81 (1999), 682–687.
50.
FriedlP.MaaserK.KleinC.E.NiggemannB.KrohneG. and ZankerK.S., Migration of highly aggressive MV3 melanoma cells in 3-dimensional collagen lattices results in local matrix reorganization and shedding of alpha2 and beta1 integrins and CD44, Cancer Res57 (1997), 2061–2070.