We aimed to investigate the influence of miR-133b/fibrillin 1 (FBN1) on proliferation and invasion of human gastric cancer (GC) cells. Carcinomatous and adjacent tissues of 43 GC patients, normal gastric mucosa cell line GES-1 and GC cell lines including AGS, HGC-27, KATO III, NCI-N87, SGC-7901, MKN-45 and MGC-803 were collected. Then, the expressions of miR-133b and FBN1 were detected by qRT-PCR. The dual luciferase reporter gene assay was conducted to determine the targeting relationship between miR-133b and FBN1.The protein expression levels of FBN1, -catenin, Cyclin D1, C-myc and MMP-7 were detected by Western Blot. Furthermore, the cell viability, proliferation, migration and invasion ability were measured by CCK-8, colony formation assay, wound healing assay and Transwell assay, respectively. MiR-133b was down-regulated in GC tissues and cells compared with adjacent tissues and normal cells. Conversely, FBN1 was up-regulated in GC tissues and cells in contrast with adjacent tissues and normal cells. MGC-803 and MKN-45 cell lines were chosen to conduct the following assays. The luciferase reporter assay proved that miR-133b directly targeted FBN1. The overexpression of miR-133b and silence of FBN1 could inhibit the cell proliferative, migratory and invasive abilities of GC cells, while the influence of down-regulated miR-133b expression and up-regulated FBN1 expression were quite the contrary. Compared with NC group, in the miR-133b mimics group, the expression of -catenin, N-cadherin and Wnt1 of Wnt/-catenin signal pathway increased, while the expressions of E-cadherin decreased. MiR-133b inhibits the proliferative, migratory and invasive abilities of GC cells by increasing FBN1 expression.
Gastric cancer (GC), also called stomach cancer, is generally regarded as one of the most dreadful carcinomas [1]. The mortality of GC in African is 2.5 times as high as that in Caucasian and males are twice as likely as females to develop GC [2, 3]. GC is differentiated according to its anatomical location into non-cardia GC, which accounts for more than 70% of incidence, and cardia GC [4]. Different dietary patterns and life styles may affect the tumorigenesis of GC. Also, chronic infection of Helicobacter pylori is a significant risk factor for GC, and the cases of non-cardia gastric cancer by reason of H. pylori infection accounted of 6.2% of the total cancer cases in 2008 [5]. Although pathogenesis and therapies of GC have been studied a lot in past years, patients who were not diagnosed timely exhibited poor five-year survival rate (10%) [6]. In recent years, genetic factors have been linked with the development of GC [7] and researchers are eager to discover effective approaches that are able to improve the survival status of GC patients.
MicroRNAs (miRNAs, miRs) are endogenous noncoding RNAs consisting of about 20 nucleotides [8] and they play a significant role in mediating post-transcriptional processes including the degradation of mRNAs and the translation of proteins [9]. MiRNAs work by binding to the 3 untranslated regions (UTR) of target mRNAs [10, 11, 12]. Furthermore, growing evidences have suggested that miRs may act as oncogenic factors or tumor suppressors because of their influence on the initiation, proliferation and metastasis of human malignancies [1, 13, 14, 15]. For example, a series of experiments conducted by Zheng et al. validated that the reintroduction of miR-329 can remarkably repress the formation of GC [8]. MiR-140 was reported as a suppressor of GC cell line HGC-27 by inhibiting cell proliferation, while recent studies verified miR-935 as a promoting factor of GC cell progress by targeting SOX7 [16, 17]. MiR-133b is widely studied on its regulatory function in skeletal differentiation, cell proliferation and cardiogenesis in muscle cells [18]. It has also been studied as biomarker in several types of carcinoma. A study conducted by Bassampour et al. in 2015 indicated that the downregulation of miR-133b works as an efficient prognostic and diagnostic indicator for patients suffering from osteosarcoma [19]. Moreover, miR-133b was indicated as a regulating factor, inhibiting epithelial-mesenchymal progression in lung cancer cells and the Warburg effect in colorectal tumors respectively [20, 21]. As for GC, several studies have revealed that miR-133b acts as a suppressor in biological progress including proliferation, metastasis and apoptosis by targeting Gli1, FSCN1 and FGFR1 [1, 13, 22]. The previous studies suggested that miR-133b may be a potential target for treatment of GC, whereas detailed functional mechanism and more potential target of miR-133b in GC cells remains undiscovered.
FBN1 (fibrillin 1) is a gene located on chromosome 15, and it is responsible for encoding a kind of protein named fibrillin 1 [23, 24]. Fibrillin 1 protein is a large extracellular matrix glycoprotein that can form threadlike filaments, microfibrils, which can provide structural support for tissues, form elastic fibers in skins and vessels and store a protein called transforming growth factor beta (TGF-) [25, 26, 27, 28, 29]. A recent study suggested that FBN1 was remarkably overexpressed in testicular germ cell tumors and was involved in the progress of in situ development of the tumors [30]. In addition, FBN1 has been reported as a promoter of ovarian cancer metastasis, which may be helpful in therapy of ovarian cancer [31]. As for the regulation of its expression, previous studies have found that FBN1 could be regulated by miRNA-mRNA regulation network, and several miRNAs, such as miR29b and miR-486-5p, have been verified as the regulator of it [32, 33]. However, neither the expression level of FBN1 nor its regulation in GC has been studied before.
The Wnt/-catenin signaling pathway is involved in the development and progression of a significant proportion of gastric cancer cases [34]. Activation of the Wnt/-catenin signaling was found in about 30% to 50% of gastric cancer tissues and in many kinds of gastric cancer cell lines [35, 36, 37]. Meanwhile, changes of the -catenin expression were observed in breast, colorectal, melanoma, prostate and other cancers [38, 39, 40, 41].
In order to verify whether miR-133b and FBN1 are potential candidates for developing target therapies of GC, we explored the relationship between miR-133b and FBN1. We also verified whether miR-133b is able to control the proliferation, metastatic and invasive ability of GC.
Primer sequences for qRT-PCR
cDNA
Forward
Reverse
GADPH
5-GCACCGTCAAGGCTGAGAAC-3
5-TGGTGAAGACGCCAGTGGA-3
U6
5-CGCTTCGGCAGCACATATAC-3
5-TTCACGAATTTGCGTGTCAT-3
C-myc
5-GCAGCTGCTTAGACGCGCTGGA-3
5-CGCAGTAGAAATACGGCTGCAC-3
Cyclin D1
5-ATGTTCGTGGCCTCTAAGATGA-3
5-CAGGTTCCACTTGAGCTTGTTC-3
MMP-7
5-GTATGGGACATTCCTCTGATCC-3
5-CCAATGAATGAATGAATGGATG-3
miR-133b
5-CTGGAGTTTGGTCCCCTTCAAC-3
5-GTGCAGGGTCCGAGGT-3
FBN1
5- GAGGCTGGGAACGTGAAG-3’
5-AGTGCTGCTGTGATGCCG-3
Materials and methods
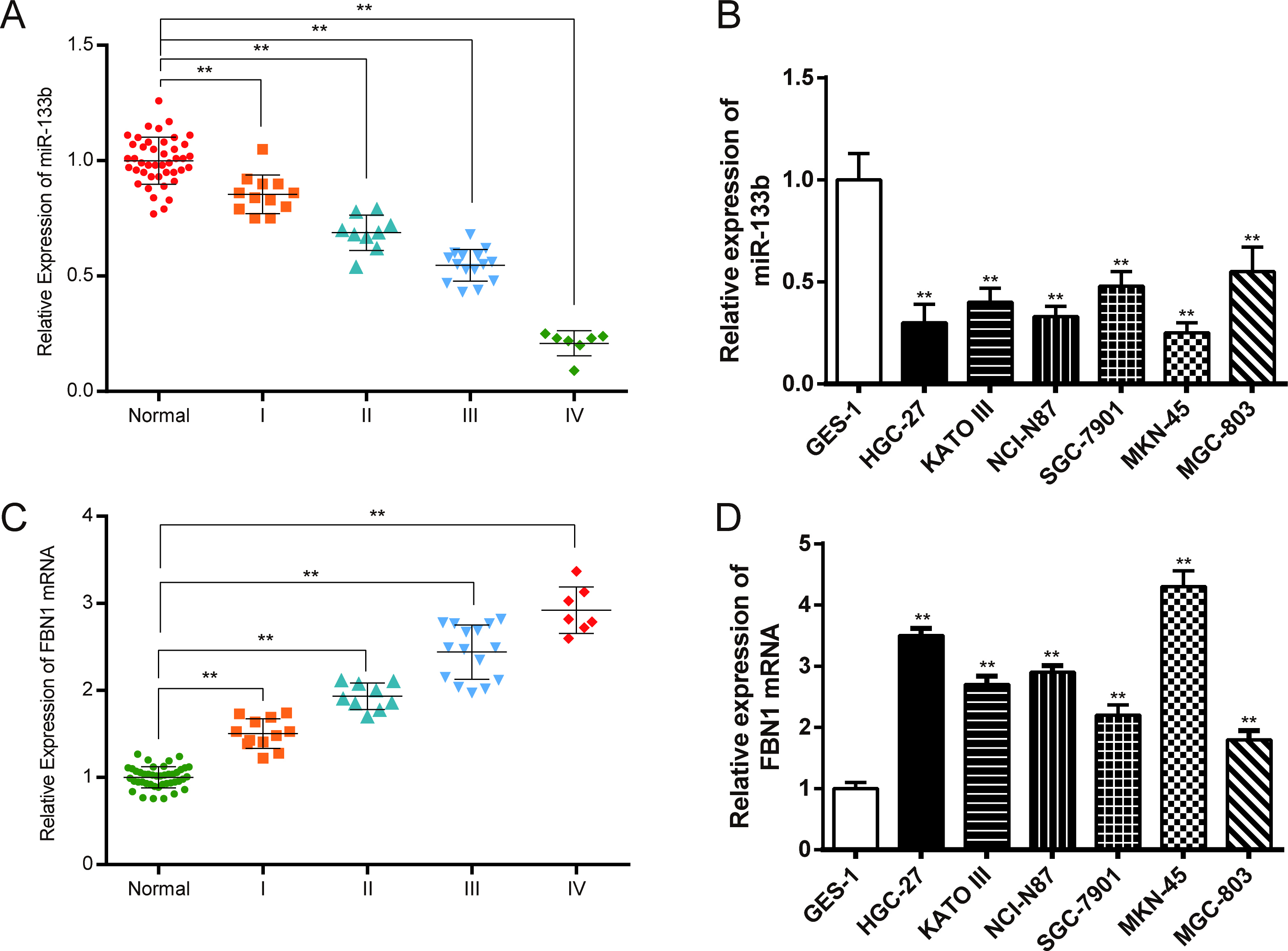
Relative expression levels of miR-133b and FBN1 in tissues and cells. (A) Expression levels of miR-133b in normal tissues and GC tissues with different stages detected by qRT-PCR; (B) Expression levels of miR-133b in normal and GC cells detected by qRT-PCR; (C) Expression levels of FBN1 in normal tissues and GC tissues with different stages detected by qRT-PCR; (D) Expression levels of FBN1 in normal and GC cells detected by qRT-PCR. Data is expressed as mean SD. , 0.01 compared with normal tissues or normal cells.
Clinical samples
The gastric cancer (GC) tissues and adjacent tissues (located 5 cm from the tumors) were collected from 43 patients who received the surgical treatment in the Linyi People’s Hospital and Chinese Medicine Hospital of Linyi City, from June, 2015 to June, 2016. The frequency-matched controls from 46 patients to these cases by age ( 5 years) and sex were collected at the same period and regions of gastric ulcer. Tissue samples were collected and immediately snap frozen in liquid nitrogen. All samples were obtained with informed consent and approved by the institutional review board of the Linyi People’s Hospital and Chinese Medicine Hospital of Linyi City.
Cell lines and cell culture
GC cell lines AGS, HGC-27, KATO III, HGC-27, and NCI-N87 were purchased from BNCC (Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences). GC cell lines SGC-7901, MGC-803 and MKN-45 and normal gastric mucosa cell line GES-1 were purchased from ATCC (American Type Culture Collection, Manassas, VA, USA). All cells were cultured in RPMI-1640 medium supplemented with 10% fetal-bovine serum in a 37C, 5% CO incubator.
Cell transfection
The GC cells were divided into six groups randomly. Cells in the six groups were transfected with non-sense sequences (NC), Lipofectamine2000 reagent (Con), miR-133b mimics (miR-133b), FBN1-siRNA (FBN1-siRNA), FBN1-cDNA (FBN1-cDNA) or co-transfected with miR-133b mimics and FBN1-cDNA (miR-133b/FBN1-cDNA), respectively (Supplemen- tary Fig. 1). MiR-133b mimics, FBN1-siRNA and FBN1-cDNA were purchased from Shanghai JiMa Pharmaceutical Co., Ltd. The manufacturer’s protocols were strictly followed. Lipofectamine2000 was used as the transfection reagent, and all the transfection materials were purchased from Invitrogen (Carlsbad, CA, USA).
Quantitative RT-PCR
TRIzol™ reagent (Invitrogen, Carlsbad, CA, USA) was used to extract total RNA from frozen tissues and cell samples. All reagents used for the stem-loop RT-PCR were purchased from Applied Biosystems (CA, USA). The PCR primer sequences were shown in Table 1. QRT-PCR was conducted by SYBR Premix Ex Taq II (TaKaRa, DALIAN, China) and measured by a LightCycler 480 system (Roche, Basel, Switzerland). GAPDH and U6 were used as the internal control. The relative expression of the RNA and miRNA was calculated by the method.
Western blot analysis
The total cell protein was extracted using RIPA lysis buffer and measured by BCA (bicinchonininc acid) method. Proteins were separated by SDS-PAGE gel and transferred to polyvinylidene difluoride (PVDF) membrane (Millipore, MA, USA). The membranes were probed with primary antibodies specific to FBN1 (1:1000, Abcam, Cambridge, UK), -catenin (1:2000, Abcam), N-cadherin (1:1000, Abcam), Wnt1 (1:1000, Abcam), E-cadherin (1:1000, Abcam) and -actin (1:1000, Abcam). The membranes were then incubated with HRP-conjugated secondary antibody goat anti-rabbit IgG (1:900, Abcam). Finally, the membranes were visualized using WesternBright™ ECL developing liquid (Pierce, IL, USA). -actin was used as internal control here.
CCK-8 assay
Transfected cells were seeded in 96-well plates (in triplicate) and cultured for 24 h. Then OD at 450 nm was measured by microplate computer software according to the protocol of the Cell Counting Kit-8 (CCK-8; Dojindo Laboratories, Japan) and proliferation curves were plotted.
Colony formation assay
Transfected cells were trypsinized and seeded into 96-well plates (in triplicate). Seven days after seeding, cells were fixed with methanol and stained with 1% crystal violet for 15 min. Then the plates were read by automated enzyme-linked spot image analyzer (Elisporeader 400 PRO-X, BIOSYS) to count the number of colonies that contained more than 50 cells.
Wound healing assay
Transfected cells were seeded into 6-well plates after trypsinization (in triplicate). After the cells grew to monolayers, a sterile 200-lL pipette tip was used to scratch a straight line through the culture surface. The open wound rate at the initial time and after 24 h was measured using a microscope.
Transwell invasion assay
The assay was performed using 8 m pore size 96-well MIC Transwell plates (Millipore) coated with matrigel (BD Biosciences, Bedford, MA, USA). Cells were resuspended and then 2.5 10 cells in 300 l medium were added to the upper chamber while the lower chamber was filled with DMEM 10% FBS as a chemoattractant. After 24 hs’ incubation, cells remained on the matrigel and upper surface of the chamber were cleaned by a cotton swab. The invaded cells were fixed in methanol, stained with gentian violet and then directly counted under a microscope in three fields (200 ).
Dual luciferase reporter gene assay
The wild type or a mutant 3UTR sequence of FBN1 were inserted into the pGL4 luciferase reporter vector (Promega, USA) to construct pGL4 -FBN1-wt and pGL4 -FBN1-mut vectors. After cultured in 96-well plates, MGC-803 and MKN-45 cells were co-transfected with miR-133b mimics or NC-mimics, pGL4 -FBN1-wt or pGL4 -FBN1-mut plasmid. Transfection was performed using Lipofectamine™ 2000 reagent (Invitrogen, USA). Cells were collected 48 h after transfection and then Dual-Luciferase Reporter Assay System (Promega, WI, USA) was used to detect renilla and firefly luciferase activity according to protocols.
Statistical analysis
All statistical analyses were performed by SPSS 21.0 (SPSS Inc., Chicago, IL, USA) and GraphPad Prism 6.0. All data were represented as mean standard deviation (SD). Comparisons between two groups were analyzed using Student’s t-test while analysis of variance (one-way ANOVA) was applied for multiple comparisons. All experiments were conducted in triplicate. A value of 0.05 was considered statistically significant.
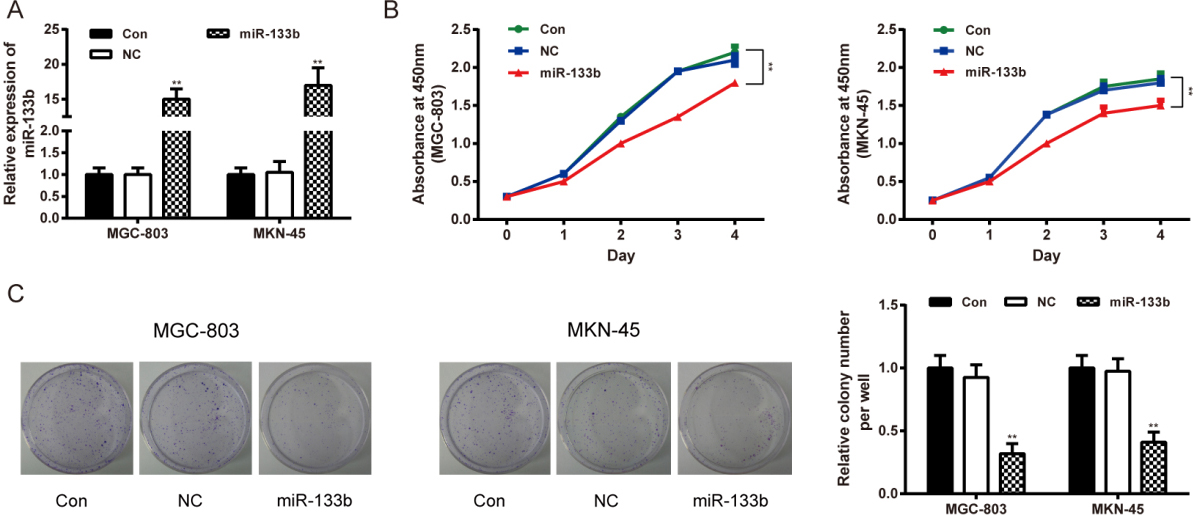
MiR-133b regulated GC cell viability and proliferative abilities. (A) qRT-PCR confirmed the transfection. Data is expressed as mean SD. , 0.01 compared with the Con group. (B) Cell viability of MGC-803 and MKN-45 cells was tested by CCK-8 assay. Data is expressed as mean SD. , 0.01 compared with the Con group. (C) Proliferation of MGC-803 and MKN-45 cells was tested by colony formation assay. Data is expressed as mean SD. , 0.01 compared with the Con group. Con: control group, cells transfected with Lipofectamine2000 reagent. NC: negative control group, cells transfected with non-sense sequences. miR-133b: miR-133b mimics group, cells transfected with miR-133b mimics.
Demographic and clinical characteristics of patients
Characteristics
Control ( 46)
Cases ( 43)
Age (Mean SD)
50.15 12.77
52.66 13.15
0.3636
Sex
0.2788
Female
14
18
Male
32
25
Smoking
0.0011
Non-smokers
33
16
Smokers
13
27
Alcoholism
0.0084
Non-alcoholic
31
17
Alcoholic
15
26
Tumor sites
0.8895
Non-cardia
25
24
Cardia
21
19
TNM stages
I
/
12
II
/
9
III
/
15
IV
/
7
applies t-test, applies Chi-square test.
Results
Characteristics of the patients
Clinico-pathological characteristics of patients and controls are presented in Table 2. This study included 89 subjects: 46 healthy individuals and 43 GC patients. As shown in the Table, the age, sex and tumor sites of the patients in the two groups did not demonstrate a significant difference ( 0.05). The difference was remarkable in smoking and alcoholism between normal patients and GC patients ( 0.01).
Expression levels of miR-133b and FBN1 mRNA in GC tissues and cells
The results of qRT-PCR suggested that miR-133b expression in cancer tissues were lower than those in normal tissues ( 0.01) (Fig. 1A). Also, miR-133b expression levels in selected GC cells were lower than those in normal gastric mucosa cell GES-1 ( 0.01) (Fig. 1B). FBN1 mRNA was high-expressed in GC cells and tissues as shown in Figs 1C and D. As shown in Fig. 1, the expressions of miR-133b and FBN1 were correlated with the different stages of GC. With the progression of GC, the expression of miR-133b decreased, while the FBN1 expression increased. MKN-45 and MGC-803 cells were selected to conduct the following assays because the miR-133b expression levels were the lowest in MKN-45 cells and the highest in MGC-803 cells among all the cancer cells.
MiR-133b regulated GC cell viability and proliferative ability
The transfection efficiency of miR-133b mimics was validated by qRT-PCR. The expression of miR-133b in the miR-133b mimics group was significantly higher than that in the control and NC groups ( 0.01). No notable difference was observed between the control and NC groups ( 0.05, Fig. 2A). In CCK-8 assay, the OD value of MGC-803 and MKN-45 cells transfected with MiR-133b mimics had been significantly smaller than those treated with reagents (Con) on the 2, 3 and 4 day after transfection ( 0.01, MGC-803; 0.01, MKN-45) (Fig. 2B). Meanwhile, colony formation assay confirmed that miR-133b over-expression inhibited the proliferative activities of MGC-803 and MKN-45 cells, compared with those treated with reagents (Con) ( 0.01, MGC-803; 0.01, MKN-45) (Fig. 2C). There was no significant difference between the control and NC groups ( 0.05).
MiR-133b inhibited GC cell migration and invasion. (A) Migratory ability of MGC-803 and MKN-45 cells was tested by wound healing assay. (B) Invasive ability of MGC-803 and MKN-45 cells was tested by Transwell assay. Data is expressed as mean SD. , 0.01 compared with the Con group. Con: control group, cells transfected with Lipofectamine2000 reagent. NC: negative control group, cells transfected with non-sense sequences. miR-133b: miR-133b mimics group, cells transfected with miR-133b mimics.
MiR-133b directly targeted FBN1. (A) Scheme and sequence of the wild-type miR-133b binding site (wt) and the corresponding mutated-type (mut) of FBN1 3UTR; (B) The FBN1 protein expression was tested by Western Blot assay; (C) The relative luciferase activity of MGC-803 and MKN-45 cells transfected with wild-type and mutated FBN1 3UTR in different subgroups. Data is expressed as mean SD. , 0.01 compared with the NC group. Con: control group, cells transfected with Lipofectamine2000 reagent. NC: negative control group, cells transfected with non-sense sequences. miR-133b: miR-133b mimics group, cells transfected with miR-133b mimics.
MiR-133b attenuated GC cells viability through targeting FBN1. (A) The FBN1 protein expression was tested by Western Blot assay; (B) Cell viability of MGC-803 and MKN-45 cells was tested by CCK-8 assay. Data is expressed as mean SD. , 0.01 compared with the Con group. (C) Proliferation of MGC-803 and MKN-45 cells was tested by colony formation assay. Data is expressed as mean SD. , 0.01 compared with the Con group. Data is expressed as mean SD. , 0.05, , 0.01 compared with the Con group. Con: control group, cells transfected with Lipofectamine2000 reagent. NC: negative control group, cells transfected with non-sense sequences. FBN1-siRNA: FBN1 siRNA, cells transfected with FBN1 siRNA. FBN1-cDNA: FBN1 cDNA, cells transfected with FBN1 cDNA. miR-133b/FBN1-cDNA: miR-133b mimics FBN1-cDNA group, cells co-transfected with miR-133b mimics and FBN1 cDNA.
MiR-133b inhibited GC cell migrating and invasive abilities
In wound healing assay, both of the GC cell lines transfected with miR-133b mimics showed significantly lower wound healing rate compared with those transfected with reagents (Con) 24 h ( 0.01, MGC-803; 0.01, MKN-45) (Fig. 3A). Meanwhile, Transwell invasion assay confirmed that the number of invaded cells dramatically decreased in groups transfected with miR-133b mimics compared with those treated with reagents (Con) ( 0.01, MGC-803; 0.01, MKN-45) (Fig. 3B). Above all, enforced expression of miR-133b inhibited GC cell migration and invasion.
MiR-133b directly targeted FBN1
MiR-133b was predicted to influence the expression of FBN1 through binding to the 3 UTR of FBN1 by TargetScan. To further confirm miR-133b’s negative regulation effect on FBN1, we investigated the FBN1 expression levels of MGC-803 and MKN-45 cells transfected with miR-133b mimics or NC-mimics (NC) using Western Blot assay. The results initially indicated that over-expression of miR-133b suppressed the protein expression of FBN1 (Fig. 4B).
In order to further detect whether miR-133b directly targeted FBN1, pGL4-FBN1-wt and pGL4-FBN1-mut (Fig. 4A) were co-transfected with miR-133b mimics or mimics-NC into MGC-803 and MKN-45 cells. As shown in Fig. 4C, the luciferase activity of FBN1 3 UTR-wt dramatically dropped down while co-transfected with miR-133b mimics compared with the NC control ( 0.01, MGC-803; 0.01, MKN-45), but not in those with respective mutant 3 UTRs. Above all, miR-133b directly targeted FBN1 and the suppression of miR-133b on FBN1 was dependent on the binding site on 3 UTR.
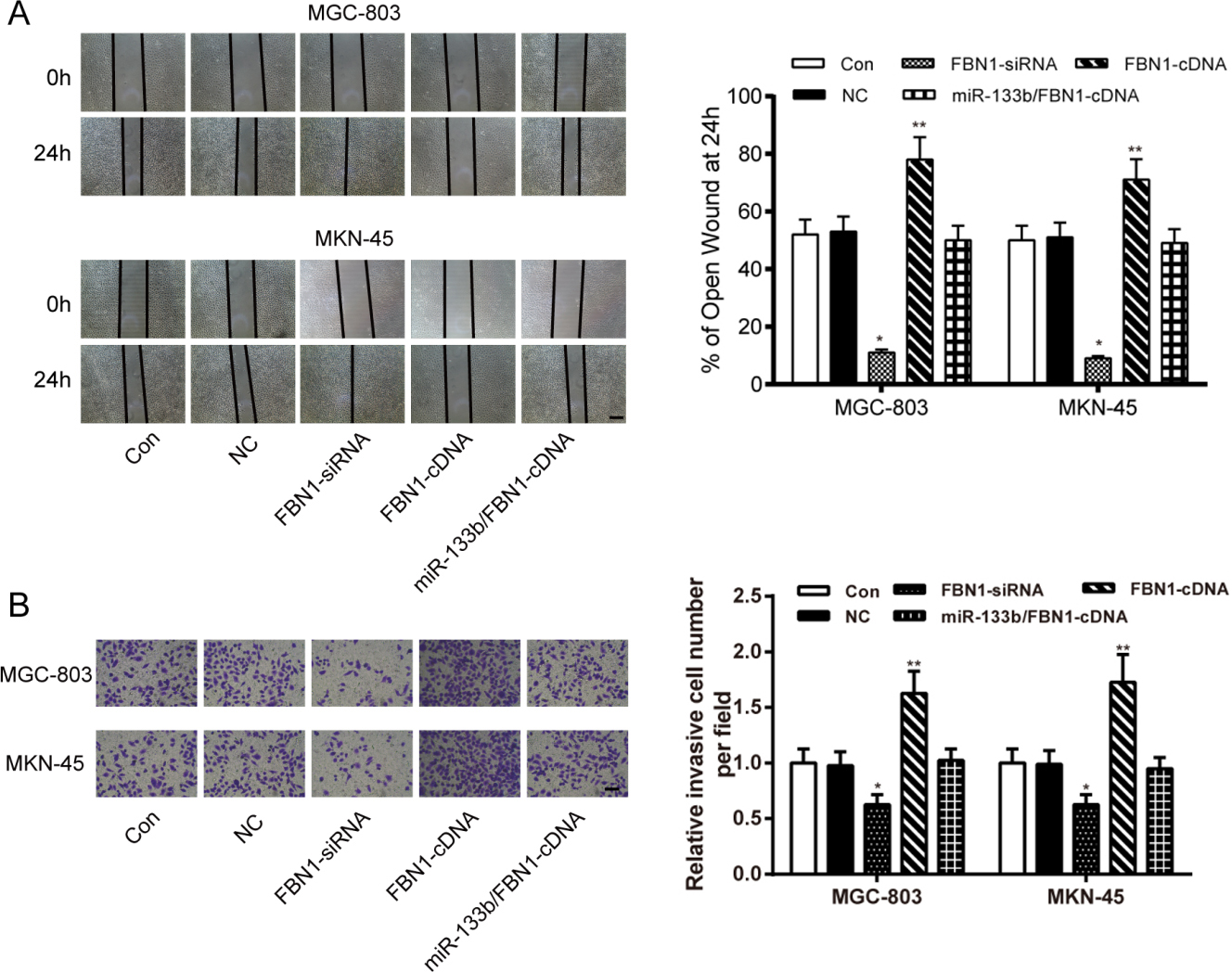
MiR-133b attenuated GC cells migration and invasion through targeting FBN1. (A) Migratory ability of MGC-803 and MKN-45 cells was tested by wound healing assay. (B) Invasive ability of MGC-803 and MKN-45 cells was tested by Transwell assay. Data is expressed as mean SD. , 0.05, , 0.01 compared with the Con group. Con: control group, cells transfected with Lipofectamine2000 reagent. NC: negative control group, cells transfected with non-sense sequences. FBN1-siRNA: FBN1 siRNA, cells transfected with FBN1 siRNA. FBN1-cDNA: FBN1 cDNA, cells transfected with FBN1 cDNA. miR-133b/FBN1-cDNA: miR-133b mimics FBN1-cDNA group, cells co-transfected with miR-133b mimics and FBN1 cDNA.
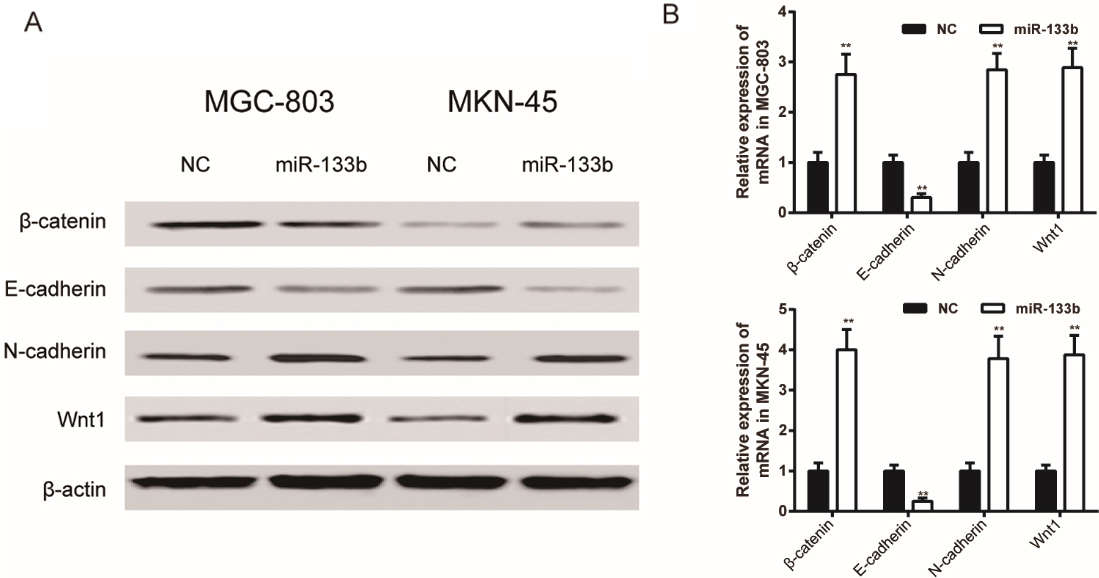
MiR-133b regulated Wnt/-catenin signal pathway through down-regulated FBN1 expression. (A) Protein expression of -catenin, E-cadherin, N-cadherin and Wnt1 of MGC-803 and MKN-45 cells was tested by Western Blot assay; (B) mRNA expression of -catenin, E-cadherin, N-cadherin and Wnt1 of MGC-803 and MKN-45 cells was tested by qRT-PCR assay. Data is expressed as mean SD. , 0.01 compared with the NC group. NC: negative control group, cells transfected with non-sense sequences. miR-133b: miR-133b mimics group, cells transfected with miR-133b mimics.
MiR-133b attenuated GC cells viability, migration and invasion through binding to FBN1
The FBN1 expression levels of MGC-803 and MKN-45 cells transfected with FBN1-cDNA, FBN1-siRNA or miR-133b/FBN1-cDNA was detected by Western Blot assay. The results showed that the expression of FBN1 in the FBN1-cDNA group increased, while the FBN1 protein expression was decreased in the FBN1-siRNA group (Fig. 5A). In CCK-8 assay, compared with the NC group, the OD value of MGC-803 and MKN-45 cells transfected with FBN1-siRNA were significantly lower than those treated with reagents (Con) ( 0.01, MGC-803; 0.01, MKN-45) while those transfected FBN1-cDNA were significantly higher ( 0.01, MGC-803; 0.01, MKN-45). Meanwhile, the GC cells co-transfected with miR-133b mimics and FBN1-cDNA showed no obvious difference compared with Control Group in average OD level (Fig. 5B). Colony formation assay confirmed that FBN1-siRNA inhibited the proliferative activities of MGC-803 and MKN-45 cells, while FBN1-cDNA facilitated the the proliferative activities compared with those treated with reagents (Con) ( 0.01, MGC-803; 0.01, MKN-45). There was no significant difference among the control, the NC and the miR-133b/FBN1-cDNA groups ( 0.05, Fig. 5C).
In Transwell invasion assay, GC cells transfected with FBN1-siRNA presented impaired migratory and invasive abilities than those treated with reagents (Con) ( 0.05, MGC-803; 0.05, MKN-45) while those transfected FBN1-cDNA showed improved ones ( 0.01, MGC-803; 0.01, MKN-45). In addition, transfection of FBN1-cDNA restored the miR-133b-mediated inhibition on the migration and invasion of MGC-803 and MKN-45 cells (Fig. 6). These results indicated that miR-133b remarkably attenuated the viability, migration and invasion of GC cells through targeting FBN1.
MiR-133b regulated Wnt/-catenin signal pathway through down-regulating FBN1 expression
In Western Blot assays, transfection of miR-133b mimics promoted the protein expression of -catenin, N-cadherin and Wnt1 and suppressed the protein expression of E-cadherin compared with the NC group (Fig. 7A). Meanwhile, it was detected by qRT-PCR assays that transfection of miR-133b mimics promoted the mRNA expression of -catenin, N-cadherin and Wnt1 ( 0.01, MGC-803; 0.01, MKN-45) and suppressed the mRNA expression of E-cadherin ( 0.01, MGC-803; 0.01, MKN-45) compared with the NC group (Fig. 7B).
Discussion
Our findings led to the conclusion that by repressing the expression of FBN1 and regulating Wnt/-catenin signal pathway, miR-133b can significantly suppress the proliferation, metastasis and invasion of stomach cancer cells.
The prognosis of curve of miR-133b and FBN1 was evaluated by Kaplan-Meier survival analysis and the choice of the targets of miR-133b was used by microarray data and qRT-PCR. (A) The prognosis of curve of miR-133b ( 0.01). (B) The prognosis of curve of FBN1 ( 0.05). (C) The differential expressed target mRNAs of miR-133b from the heat map of microarray data. (D) The expressions of the top-ten differential expressed mRNAs have been detected by qRT-PCR.
For the time being, the treatments for GC are limited to the combination of surgery, chemotherapy, radiation therapy and targeted therapy, mainly based on general clinical data and histological features rather than individual features [42]. In spite of advances in modern medical technology, serviceable and effective treatments for GC are yet to be developed, thus the overall outcome of GC currently remains poor [6, 43]. Molecular markers carrying individual features such as miRNAs have been identified to be therapy targets in GC, but very few has come to the clinical practice due to the controversial role of the markers [42]. Consequently, there is an urge for identifying specific biomarkers in the clinical management of GC [43]. So far there have been no reliable biomarkers that could help with predicting the prognosis and the response to chemotherapy individually. The challenge of identifying a novel and reliable biomarker mainly lies in the technical shortage of assessing molecular alterations quantitatively [42]. Also, the varied responses and drug resistance of patients are still huge hinders lying in front of us. Although a few successful markers have been discovered, the treatments are only confined to individual features. Additionally, the sensitivity of targeted therapy is still questionable. To make it worse, at the moment the published conclusions are suspiciously incompatible from time to time [42, 43]. It is undeniable that current targeted therapy is flawed and immature, but considering the trend that emphases over miRs are slowly shifting from basic molecular exploration to clinical application, miRs might act as potential diagnostic biomarkers or treatments for clinical management of GC [43].
Various in vitro and in vivo studies demonstrated that GC cells presented disordered miRNA expression patterns compared with normal gastric cells [44, 45, 46, 47, 48]. MiR-133b was first discovered to be dysregulated in GC tissues in 3 patients [49]. Wang Z et al. claimed that -catenin could be blocked by FBN1 [31]. The down-regulation of Wnt/-catenin signaling pathway was reported to be constantly observed in various human cancers, and the pathway was proven to be involved in the development, differentiation and metastasis of GC [34, 50]. Our results revealed that miR-133b was down-regulated in GC tissues, while FBN1’s expression turned out otherwise compared with adjacent non-tumor tissues, and we also confirmed the targeting relationship between miR-133b and FBN1 in the present study. We thus speculated that miR-133b could affect GC by regulating FBN1.
Wnt signaling has been considered to be associated with cancer since its initial discovery. Mao et al. demonstrated that activation of Wnt1 signaling accelerates the proliferation of gastric cancer stem cells [51]. With further efforts, Liu et al. confirmed that the activity of the Wnt/-catenin signaling pathways was suppressed with the expression of E-cadherin increased and the expression of N-cadherin decreased in GC [52]. In our study, the expression levels of -catenin, N-cadherin and Wnt1 in miR-133b mimics group was significantly higher than that in NC group, while the expression level of E-cadherin was remarkably lower in the miR-133b mimics group compared with the NC group. Our results suggested that the overexpression of miR-133b may regulate Wnt/-catenin signal pathway.
To sum up, our study provides a firm support to the assertion that miR-133b is able to control the proliferation, metastasis and invasion of stomach cancer cells by targeting FBN1. The regulatory function of miR-133b is realized by mediating Wnt/-catenin signaling pathway, a pathway assumed to be involved in various cancers. The abnormal expression of miR-133b in GC may serve as a candidate biomarker for prognosis and diagnosis of GC and may further contribute much more to the exploration of therapeutic treatment of GC.
Footnotes
Conflict of interest
The authors declare that they have no conflicts of interest concerning this article.
References
1.
GuoL.BaiH.ZouD.HongT.LiuJ.HuangJ.HeP.ZhouQ. and HeJ., The role of microrna-133b and its target gene fscn1 in gastric cancer, J Exp Clin Cancer Res33 (2014), 99.
2.
SiegelR.L.MillerK.D. and JemalA., Cancer statistics, 2016, CA Cancer J Clin66 (2016), 7–30.
3.
TorreL.A.BrayF.SiegelR.L.FerlayJ.Lortet-TieulentJ. and JemalA., Global cancer statistics, 2012, CA Cancer J Clin65 (2015), 87–108.
4.
Van CutsemE.SagaertX.TopalB.HaustermansK. and PrenenH., Gastric cancer, Lancet388 (2016), 2654–2664.
5.
PlummerM.FranceschiS.VignatJ.FormanD. and de MartelC., Global burden of gastric cancer attributable to helicobacter pylori, International Journal of Cancer136 (2015), 487–490.
6.
OrdituraM.GaliziaG.SforzaV.GambardellaV.FabozziA.LaterzaM.M.AndreozziF.VentrigliaJ.SavastanoB.MabiliaA.LietoE.CiardielloF. and De VitaF., Treatment of gastric cancer, World Journal of Gastroenterology20 (2014), 1635–1649.
7.
SchraderK. and HuntsmanD., Hereditary diffuse gastric cancer, Cancer Treat Res155 (2010), 33–63.
8.
LiZ.YuX.WangY.ShenJ.WuW.K.LiangJ. and FengF., By downregulating tiam1 expression, microrna-329 suppresses gastric cancer invasion and growth, Oncotarget6 (2015), 17559–17569.
9.
ShiY.WangJ.XinZ.DuanZ.WangG. and LiF., Transcription factors and microrna-co-regulated genes in gastric cancer invasion in ex vivo, PLoS One10 (2015), e0122882.
10.
OhdairaH.SekiguchiM.MiyataK. and YoshidaK., Microrna-494 suppresses cell proliferation and induces senescence in a549 lung cancer cells, Cell Proliferation45 (2012), 32–38.
11.
SchirmerU.DobersteinK.RuppA.K.BretzN.P.WuttigD.KiefelH.BreunigC.FieglH.Muller-HolznerE.ZeillingerR.SchusterE.ZeimetA.G.SultmannH. and AltevogtP., Role of mir-34a as a suppressor of l1cam in endometrial carcinoma, Oncotarget5 (2014), 462–472.
12.
CodoP.WellerM.MeisterG.SzaboE.SteinleA.WolterM.ReifenbergerG. and RothP., Microrna-mediated down-regulation of nkg2d ligands contributes to glioma immune escape, Oncotarget5 (2014), 7651–7662.
13.
WenD.LiS.JiF.CaoH.JiangW.ZhuJ. and FangX., Mir-133b acts as a tumor suppressor and negatively regulates fgfr1 in gastric cancer, Tumour Biol34 (2013), 793–803.
14.
ZhangY.ZhangZ.LiZ.GongD.ZhanB.ManX. and KongC., Microrna-497 inhibits the proliferation, migration and invasion of human bladder transitional cell carcinoma cells by targeting e2f3, Oncology Reports36 (2016), 1293–1300.
15.
ZengL.P.HuZ.M.LiK. and XiaK., Mir-222 attenuates cisplatin-induced cell death by targeting the ppp2r2a/akt/mtor axis in bladder cancer cells, Journal of Cellular and Molecular Medicine20 (2016), 559–567.
16.
ZouJ. and XuY., Microrna-140 inhibits cell proliferation in gastric cancer cell line hgc-27 by suppressing sox4, Med Sci Monit22 (2016), 2243–2252.
17.
YangM.CuiG.DingM.YangW.LiuY.DaiD. and ChenL., Mir-935 promotes gastric cancer cell proliferation by targeting sox7, Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie79 (2016), 153–158.
18.
YuH.LuY.LiZ. and WangQ., Microrna-133: Expression, function and therapeutic potential in muscle diseases and cancer, Curr Drug Targets15 (2014), 817–828.
19.
BassampourS.A.AbdiR.BahadorR.ShakeriM.TorkamanA.YahaghiE. and TaheriazamA., Retracted article: Downregulation of mir-133b/mir-503 acts as efficient prognostic and diagnostic factors in patients with osteosarcoma and these predictor biomarkers are correlated with overall survival, Tumour Biology: The Journal of the International Society for Oncodevelopmental Biology and Medicine (2015).
20.
XiaoB.LiuH.GuZ. and JiC., Expression of microrna-133 inhibits epithelial-mesenchymal transition in lung cancer cells by directly targeting foxq1, Arch Bronconeumol (2016).
21.
TaniguchiK.SakaiM.SugitoN.KumazakiM.ShinoharaH.YamadaN.NakayamaT.UedaH.NakagawaY.ItoY.FutamuraM.UnoB.OtsukiY.YoshidaK.UchiyamaK. and AkaoY., Ptbp1-associated microrna-1 and -133b suppress the warburg effect in colorectal tumors, Oncotarget (2016).
22.
ZhaoY.HuangJ.ZhangL.QuY.LiJ.YuB.YanM.YuY.LiuB. and ZhuZ., Mir-133b is frequently decreased in gastric cancer and its overexpression reduces the metastatic potential of gastric cancer cells, BMC Cancer14 (2014), 34.
23.
BieryN.J.EldadahZ.A.MooreC.S.StettenG.SpencerF. and DietzH.C., Revised genomic organization of fbn1 and significance for regulated gene expression, Genomics56 (1999), 70–77.
24.
FaivreL.GorlinR.J.WirtzM.K.GodfreyM.DagoneauN.SamplesJ.R.Le MerrerM.Collod-BeroudG.BoileauC.MunnichA. and Cormier-DaireV., In frame fibrillin-1 gene deletion in autosomal dominant weill-marchesani syndrome, J Med Genet40 (2003), 34–36.
25.
DavisM.R.AnderssonR.SeverinJ.de HoonM.BertinN.BaillieJ.K.KawajiH.SandelinA.ForrestA.R.SummersK.M. and ConsortiumF., Transcriptional profiling of the human fibrillin/ltbp gene family, key regulators of mesenchymal cell functions, Molecular Genetics and Metabolism112 (2014), 73–83.
26.
SakaiL.Y.KeeneD.R. and EngvallE., Fibrillin, a new 350-kd glycoprotein, is a component of extracellular microfibrils, The Journal of Cell Biology103 (1986), 2499–2509.
27.
IsogaiZ.OnoR.N.UshiroS.KeeneD.R.ChenY.MazzieriR.CharbonneauN.L.ReinhardtD.P.RifkinD.B. and SakaiL.Y., Latent transforming growth factor beta-binding protein 1 interacts with fibrillin and is a microfibril-associated protein, The Journal of Biological Chemistry278 (2003), 2750–2757.
28.
Massam-WuT.ChiuM.ChoudhuryR.ChaudhryS.S.BaldwinA.K.McGovernA.BaldockC.ShuttleworthC.A. and KieltyC.M., Assembly of fibrillin microfibrils governs extracellular deposition of latent tgf beta, Journal of Cell Science123 (2010), 3006–3018.
29.
ZilberbergL.TodorovicV.DabovicB.HoriguchiM.CourousseT.SakaiL.Y. and RifkinD.B., Specificity of latent tgf-beta binding protein (ltbp) incorporation into matrix: Role of fibrillins and fibronectin, Journal of Cellular Physiology227 (2012), 3828–3836.
30.
CiernaZ.MegoM.JurisicaI.MachalekovaK.ChovanecM.MiskovskaV.SvetlovskaD.KalavskaK.RejlekovaK.KajoK.MardiakJ. and BabalP., Fibrillin-1 (fbn-1) a new marker of germ cell neoplasia in situ, BMC Cancer16 (2016), 597.
31.
WangZ.LiuY.LuL.YangL.YinS.WangY.QiZ.MengJ.ZangR. and YangG., Fibrillin-1, induced by aurora-a but inhibited by brca2, promotes ovarian cancer metastasis, Oncotarget6 (2015), 6670–6683.
32.
MerkD.R.ChinJ.T.DakeB.A.MaegdefesselL.MillerM.O.KimuraN.TsaoP.S.IosefC.BerryG.J.MohrF.W.SpinJ.M.AlviraC.M.RobbinsR.C. and FischbeinM.P., Mir-29b participates in early aneurysm development in marfan syndrome, Circulation Research110 (2012), 312–324.
33.
MaX.WeiJ.ZhangL.DengD.LiuL.MeiX.HeX. and TianJ., Mir-486-5p inhibits cell growth of papillary thyroid carcinoma by targeting fibrillin-1, Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie80 (2016), 220–226.
34.
ChiurilloM.A., Role of the wnt/beta-catenin pathway in gastric cancer: An in-depth literature review, World J Exp Med5 (2015), 84–102.
35.
OoiC.H.IvanovaT.WuJ.LeeM.TanI.B.TaoJ.WardL.KooJ.H.GopalakrishnanV.ZhuY.ChengL.L.LeeJ.RhaS.Y.ChungH.C.GanesanK.SoJ.SooK.C.LimD.ChanW.H.WongW.K.BowtellD.YeohK.G.GrabschH.BoussioutasA. and TanP., Oncogenic pathway combinations predict clinical prognosis in gastric cancer, PLoS Genetics5 (2009), e1000676.
36.
IkenoueT.IjichiH.KatoN.KanaiF.MasakiT.RengifoW.OkamotoM.MatsumuraM.KawabeT.ShiratoriY. and OmataM., Analysis of the beta-catenin/t cell factor signaling pathway in 36 gastrointestinal and liver cancer cells, Japanese Journal of Cancer Research: Gann93 (2002), 1213–1220.
37.
ClementsW.M.WangJ.SarnaikA.KimO.J.MacDonaldJ.Fenoglio-PreiserC.GrodenJ. and LowyA.M., Beta-catenin mutation is a frequent cause of wnt pathway activation in gastric cancer, Cancer Research62 (2002), 3503–3506.
38.
NusseR., Wnt signaling in disease and in development, Cell Research15 (2005), 28–32.
39.
RakowskiW., Personal health practices, health status, and expected control over future health, J Community Health11 (1986), 189–203.
40.
HoweL.R. and BrownA.M., Wnt signaling and breast cancer, Cancer Biology & Therapy3 (2004), 36–41.
41.
AnastasJ.N. and MoonR.T., Wnt signalling pathways as therapeutic targets in cancer, Nature Reviews Cancer13 (2013), 11–26.
42.
PietrantonioF.De BraudF.Da PratV.PerroneF.PierottiM.A.GariboldiM.FanettiG.BiondaniP.PellegrinelliA.BossiI. and Di BartolomeoM., A review on biomarkers for prediction of treatment outcome in gastric cancer, Anticancer Research33 (2013), 1257–1266.
43.
WangF.SunG.P.ZouY.F.HaoJ.Q.ZhongF. and RenW.J., Micrornas as promising biomarkers for gastric cancer, Cancer Biomarkers: Section A of Disease Markers11 (2012), 259–267.
44.
ZhangX.LiX.TanZ.LiuX.YangC.DingX.HuX.ZhouJ.XiangS.ZhouC. and ZhangJ., Microrna-373 is upregulated and targets tnfaip1 in human gastric cancer, contributing to tumorigenesis, Oncol Lett6 (2013), 1427–1434.
45.
GuoL.H.LiH.WangF.YuJ. and HeJ.S., The tumor suppressor roles of mir-433 and mir-127 in gastric cancer, International Journal of Molecular Sciences14 (2013), 14171–14184.
46.
SongY.X.YueZ.Y.WangZ.N.XuY.Y.LuoY.XuH.M.ZhangX.JiangL.XingC.Z. and ZhangY., Microrna-148b is frequently down-regulated in gastric cancer and acts as a tumor suppressor by inhibiting cell proliferation, Mol Cancer10 (2011), 1.
47.
ChenG.ShenZ.L.WangL.LvC.Y.HuangX.E. and ZhouR.P., Hsa-mir-181a-5p expression and effects on cell proliferation in gastric cancer, Asian Pac J Cancer Prev14 (2013), 3871–3875.
48.
XuX.D.HeX.J.TaoH.Q.ZhangW.WangY.Y.YeZ.Y. and ZhaoZ.S., Abnormal expression of mir-301a in gastric cancer associated with progression and poor prognosis, Journal of Surgical Oncology108 (2013), 197–202.
49.
GuoJ.MiaoY.XiaoB.HuanR.JiangZ.MengD. and WangY., Differential expression of microrna species in human gastric cancer versus non-tumorous tissues, Journal of Gastroenterology and Hepatology24 (2009), 652–657.
50.
ZhangH. and XueY., Wnt pathway is involved in advanced gastric carcinoma, Hepato-gastroenterology55 (2008), 1126–1130.
51.
MaoJ.FanS.MaW.FanP.WangB.ZhangJ.WangH.TangB.ZhangQ.YuX.WangL.SongB. and LiL., Roles of wnt/beta-catenin signaling in the gastric cancer stem cells proliferation and salinomycin treatment, Cell Death Dis5 (2014), e1039.
52.
LiuX.LiZ.SongY.WangR.HanL.WangQ.JiangK.KangC. and ZhangQ., Aurka induces emt by regulating histone modification through wnt/beta-catenin and pi3k/akt signaling pathway in gastric cancer, Oncotarget (2016).