Cervical cancer is the most common cancer in Indian women. Infection with a high-risk human papillomavirus (HR-HPV) is the greatest risk factor for developing cervical cancer. The genetic and epigenetic changes in the tumor suppressor p16 gene is play an important role in the development of cervical cancer.
OBJECTIVE:
To evaluate the expression and promoter methylation of p16 gene in HR-HPV infected squamous cell carcinoma of the uterine cervix.
METHODS:
To find out p16 expression and methylation status 105 squamous cell carcinoma of the uterine cervix were investigated by using immunohistochemistry and Methylation Specific PCR techniques.
RESULTS:
HPV16/18 was amplified in 83.8% cases of the cervix. 80% of them were positive for HPV type 16, while only 3.8% were positive for HPV type 18. Promoter CpG island hypermethylation of p16 gene was detected in 20.9% tissue samples of cervical carcinoma. Of these hypermethylated samples 90.9% cases showed nil/very low p16 expression ( 0.001). Overexpression of p16 was observed in 73.3% cases of HR-HPV infected squamous cell carcinoma of the cervix.
CONCLUSION:
An association between p16 methylation, expression, and HR-HPV infection suggested the compliance of HPV infection and aberration of p16 gene have a synergic effect on initiation and progression of cervical carcinoma.
Cervical cancer is the cancer of the uterine cervix. The most awful feature of cervical cancer is the number of deaths caused by this disease and the sufferers are the young age females often having minors. It is instigated by a virus (Human Papilloma Virus). Though, it is a curable disease and vaccines are also available against the causative agent, worldwide cervical cancer affects about 5,00,000 women and causes 2,75,000 deaths annually [1]. The burden of cervical cancer in India is enormous accounting for about 25 per cent of global cervical cancer and almost all cases harbor high risk HPV infection [2]. Previous epidemiological studies have also shown the presence of Human Papilloma Virus (HPV) infection in invasive cervical carcinoma and its precursor lesions [3]. HPV 16 and 18 are being the most commonly detected high risk HPV (HR-HPV) types in these lesions. However, HPV alone is not culpable for carcinogenesis, in addition to which, alterations in genetic makeup of the host is the major contributing factor involved in cervical carcinogenesis as most of the patients with HPV associated lesions do not progress to invasive cancer. Therefore, it is important to identify those genetic and epigenetic alterations occurring in the host which are involve in carcinogenesis, and obviously cell cycle regulatory genes plead more attention in this regard.
9p21 region of the human chromosome has apparent heterozygous and homozygous deletion in a variety of human cancers including malignant melanoma, glioma, pancreatic, renal, as well as gynecological cancers [4]. The p16 gene located in this region, is a tumor suppressor gene, and expected to be suppressed during development of cancer. However, published data signify over-expression of p16 protein in high-risk HPV infected cervical cancers [5]. It has been shown that HPV 16/18 E7 oncoprotein is responsible for the increased p16 levels [6]. Expression studies of p16 gene also demonstrated its inactivation either by homozygous deletion, mutation, or hypermethylation of the promoter region [7]. The extents of alterations in p16 gene vary in different populations and ethnic groups. Therefore we were interested to know the methylation and expression status of p16 gene in HPV 16/18 associated squamous cell carcinoma of the uterine cervix among the Indian population.
Materials and methods
Tissue specimens
In the present study, 105 female patients diagnosed with squamous cell carcinoma (SCC) of the uterine cervix were included. They were under treatment at Cancer Clinics of Lok Nayak Hospital, New Delhi. Thirty five control samples of cervical tissues obtained from female patients suffering from gynecological problems but not diagnosed of cervix cancer were included in the study. Cases of SCC were classified and graded according to the World Health Organization criteria and staged according to criteria of the International Federation of Gynecology and Obstetrics (FIGO). The Institutional ethical committee approved the study and informed consent was obtained from all patients.
DNA extraction
Genomic DNA was extracted from tumor and normal tissue samples by phenol/chloroform method as described by Sambrook et al. [8]. Briefly, about 50 mg tissue was gently minced and suspended in 1 ml TE buffer (10 mM Tris-HCL, 1 mM EDTA, pH 8.0). To this suspension 0.5 ml lyses buffer (3% SDS, 2X TE, pH 8.0) and Proteinase K (final concentration of 100 g/ml) were added and incubated overnight at 37C. Equal volume of phenol/chloroform was added and further processed to isolate the DNA from the aqueous phase of the solution. DNA was precipitated with 3M sodium acetate and absolute ethanol. Finally, DNA was dissolved and stored in 1X TE buffer at 20C.
HPV detection and typing
Presence of High-risk HPV-16/18 infections in cervical DNA samples were detected by Polymerase Chain Reaction (PCR) using type specific primers: 5’ AAG GCC AAC TAA ATG TCA C-3’ (sense), 5’ CGT CTT TTA TAC TAA CCG G-3’ (antisense) for HPV 16; 5’ ACC TTA ATG AAA AAC CAC GA-3’ (sense), 5’ CGT CGT TTA GAG TCG TTC CTG-3’ (antisense) for HPV-18. PCR was carried out in 25 l master mixture with 35 cycles of amplification. Each cycle comprises of denaturation at 94C for 45 sec, annealing at 56C for 45 sec, and extension at 72C for 1 min. -globin gene primers were used as an internal control of the PCR reaction. Amplified PCR products were electrophoresed on 3% agarose gel. HPV presence were confirmed by the presence of 269 bp band for HPV 16 and 100 bp band for HPV 18 along with the 312 bp band of -globin gene. Absence of HPV in the samples were assured by presence of only -globin band and absence of HPV 16/18 specific bands.
Frequency of methylation in different clinical stages of the HPV infected squamous cell carcinoma
Clinical stage
No. of patients (n)
Methylation positivity
HPV status
Positive
Negative
I
12
2
2
0
II
33
5
3
2
III
57
15
12
3
IV
03
00
0
0
I II III IV
105
22 (20.9%)
17 (77.3%)
5 (22.7%)
Bisulfite modification of DNA and Methylation Specific PCR (MSP)
The genomic DNA was treated with sodium bisulfite to convert unmethylated cytosine to uracil. The methylated cytosine remains unaffected during this process. Modified DNA was then purified using Wizard DNA clean-up Resin (Promega, Wisconsin USA). Desulfonation of the uracil ring was done by treatment with 3M NaOH solution. Alkaline samples were neutralization by 3M ammonium acetate and modified DNA was precipitated by ethanol and resuspended in TE buffer. Methylation status of the p16 gene promoter was determined by MSP method as described by Herman et al. [9]. The primer sequences specific for methylated (sense; 5’ TTA TTA GAG GGT GGG GCG GAT CGC, antisense; 5’ GAC CCC GAA CCG CGA CCG TAA) and unmethylated cytocine (sense; 5’ TTA TTA GAG GGT GGG GTG GAT TGT, antisense; 5’ CAA CCC CAA ACC ACA ACC ATA A) of the promoter region of p16 gene were taken from Herman et al. [9]. PCR amplification was performed in 25 l of reaction mixture contained 150–200 ng of modified DNA as template along with other PCR reagents. The temperature profile of the 40 cycles of the PCR were 95C for 30 sec (Denaturation), 60C (methylated primers) and 65C (unmethylated primers) for 30 sec (Annealing), and 72C for 30 sec (extension), and final extension at 72C for 2 min. Amplified PCR products were visualized on 3% ethidium bromide stained agarose gels.
Immunohistochemical detection of p16 expression
The p16 protein expression was determined by Immunohistochemistry (IHC) method using a monoclonal antibody raised against full length human p16 protein (Lab Vision Corporation, USA). Histologically prepared sections on poly-L-lysine coated glass slides of the cervix samples were first deparaffinized and subsequently rehydrated. The endogenous peroxidase activity was quenched by treating the samples with 0.3% HO/methanol. For antigen retrieval the slides were heated at 90C for 15 min in 0.01M sodium phosphate/citrate buffers (pH 8.0). After bringing to normal temperature slides were rinsed in 0.01M phosphate-buffered saline (PBS, pH 7.4). Sections were incubated with a primary monoclonal antibody against p16 (1:500 dilution) and placed overnight at 4C. After three washing with PBS, the slides were incubated with biotinylated anti-mouse secondary IgG (1:100 dilution) for 30 min at room temperature. To this added Avidin-biotin-peroxidase complex for an additional 30 min. Peroxidase activity was then visualized by adding 100–200 l 0.02% 3–3’-diaminobenzidine tetrahydrochloride substrate containing 0.005% HO just for 5 min. Slides were then rinsed with water and counterstained with hematoxylin for 1 min. Slides were dehydrated with alcohol grading and mounted with Canada Balsam. The presence of p16 protein was scored according to Geradts et al. [10].
Statistical analysis
The association of expression and methylation of p16 gene with different clinico-pathological variables and HPV infection was determined by Chi-square analysis. Probability values of 0.05 were regarded as statistically significant.
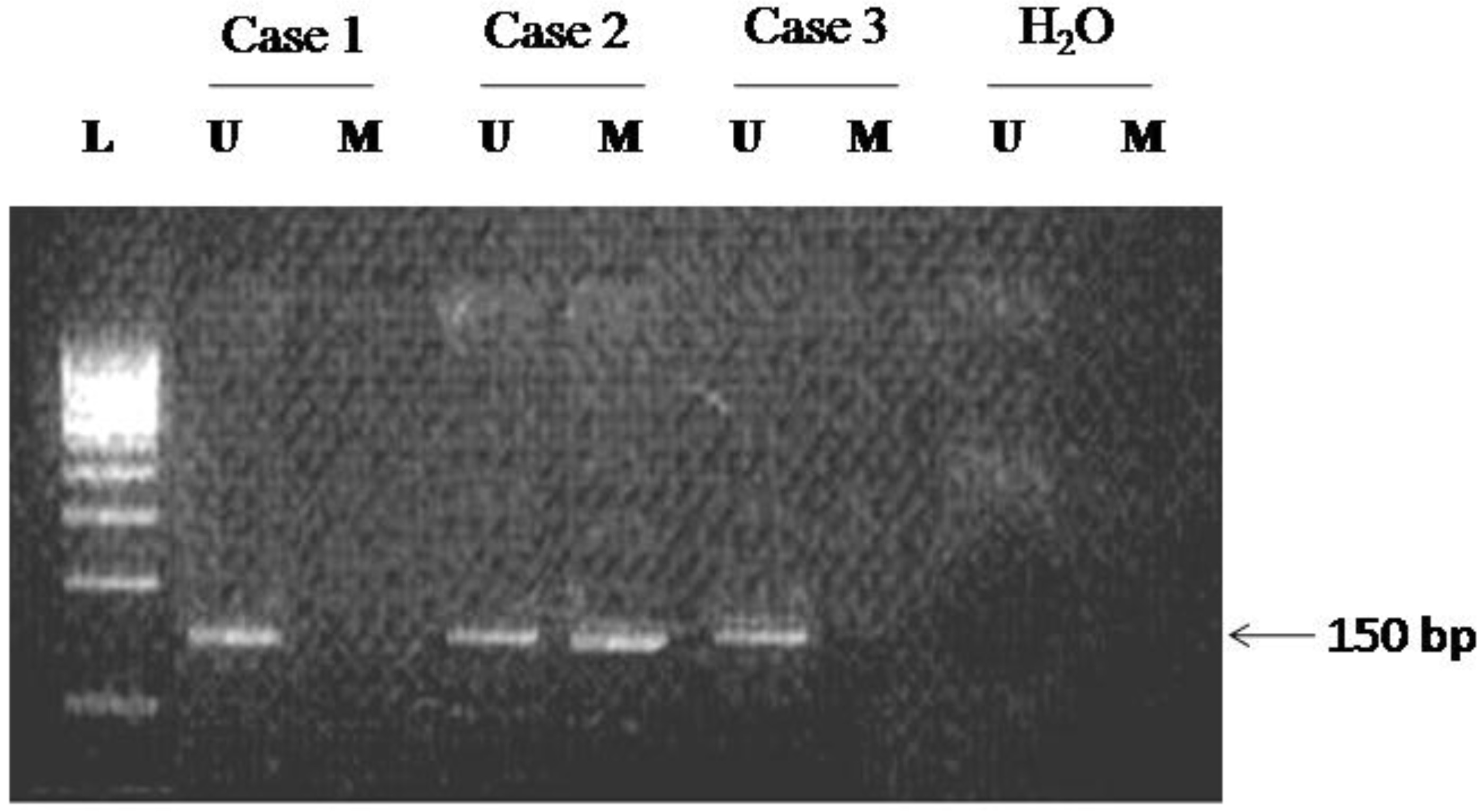
Methylation-specific PCR of the promoter region of p16 gene in cervical carcinomas. Both the methylated (M) and unmethylated (U) PCR products are shown for each sample. The presence of PCR product (151 bp) in lane U indicates the presence of unmethylated genes; the presence of product (150 bp) in lane M indicates the presence of methylated genes. HO; water blank for negative PCR control.
Expression level of p16 protein in different clinical stages
Clinical stages
No. of patients ( n)
Expression level of p16 protein
Nil ()
Weak ()
Medium ()
Strong ()
I
12
1
(8.3)
0
(00)
3
(25.0)
8
(66.7)
II
33
3
(9.1)
2
(6.1)
4
(12.1)
24
(72.7)
III
57
8
(14.0)
6
(10.5)
00
(0.0)
43
(75.4)
IV
03
00
(0.0)
1
(33.0)
00
(0.0)
2
(66.6)
Total
105
12
(11.4)
09
(8.6)
07
(6.7)
77
(73.3)
Figures in parenthesis are percentages.
Correlation of expression and methylation in HPV infected squamous cell carcinoma
Expression level of p16 protein
No. of cases
Methylation status
HPV status
Positive
Negative
Nil ()
12
12
9
3
Weak ()
9
8
5
4
Medium ()
7
0
2
5
Strong ()
77
2
72
5
Total
105
22
88
17
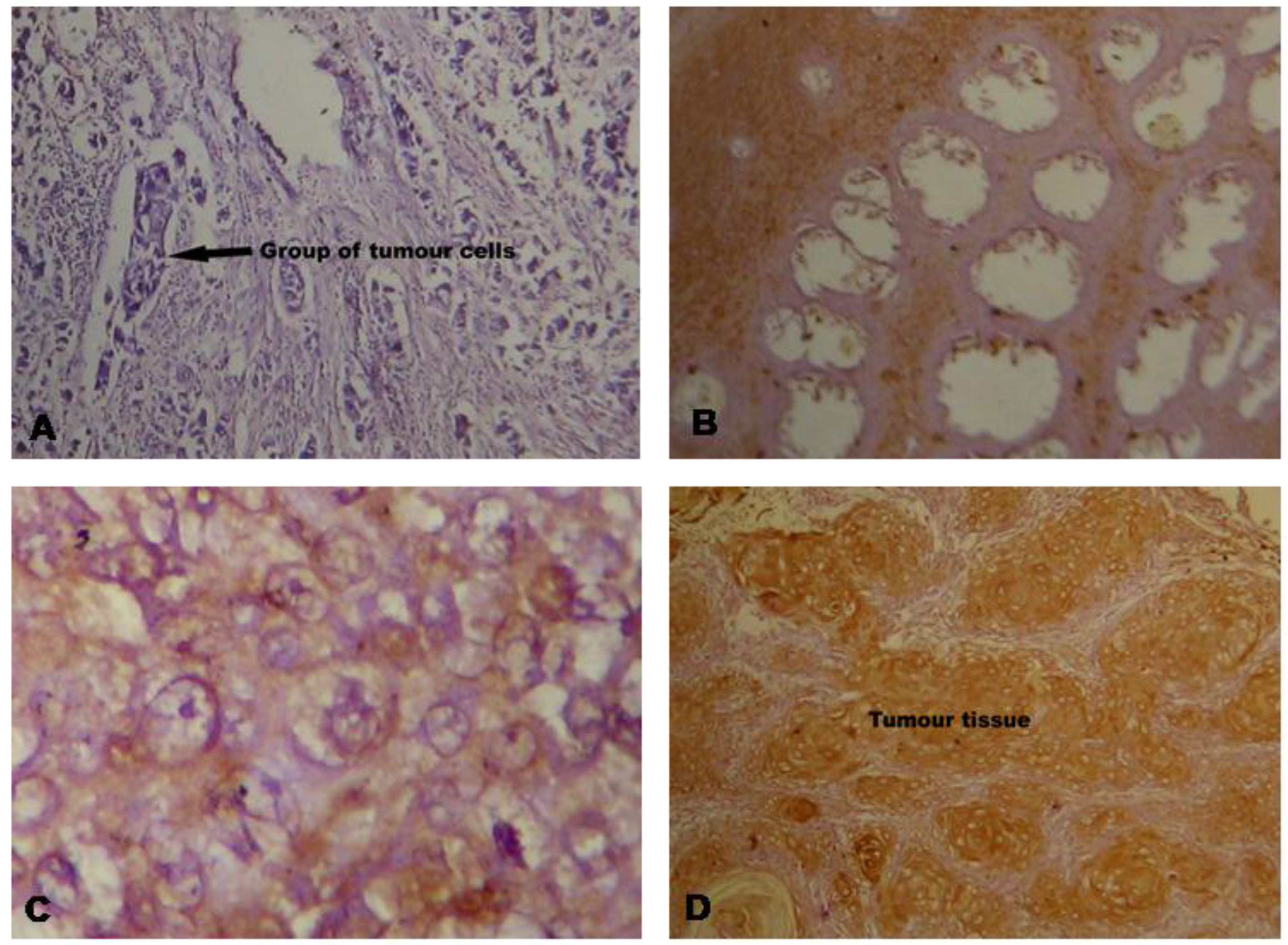
Immunohistochemical staining of the p16 protein in the tissue of squamous cell carcinoma of the cervix: (A) No expression; (B) Under Expression; (C) Normal Expression; (D) Over Expression. (Magnification: (A) & (B) at 100x and (C) & (D) at 600x).
Results
Prevalence of high risk HPV 16/18 infection in cervical cancer
High-risk HPV (HPV 16/18) infection was detected in 83.8% cases of cervical carcinoma. 80% of them were positive for HPV 16, while only 3.8% were positive for HPV 18. None was found to be positive for both HPV 16 and 18. In control samples, only 17.5% were observed positive for HPV 16 and none for HPV 18.
Methylation status of the p16 gene
Promoter CpG island hypermethylation of p16 gene was detected in 20.9% tissue samples of cervical carcinoma. All the normal control samples were negative for hypermethylation of p16 gene (Fig. 1). In 90.9% samples of hypermethylated promoter, expression of p16 protein was either nil or very low. Apparently, a negative relationship between CpG promoter hypermethylation and expression of p16 gene is obvious in squamous cell carcinoma of cervix ( 0.001). There was no statistically significant correlation in the frequencies of p16 methylation of early (I & II) and advanced stages (III & IV) of the disease ( 0.4708), and presence of HPV 16/18 in squamous cell carcinoma of the cervix (Table 1).
Expression analysis of p16 by immunohistochemistry
Immunohistochemical (IHC) analysis was per- formed to analyze the expression pattern of p16 protein in the squamous cell carcinoma of cervix. Overexpression of p16 was observed in 73.3% cases of squamous cell carcinoma of the cervix (Table 2). Comparatively, higher expression was detected in advanced stages rather in early stages of carcinomas. Representative photographs of the IHC staining of the p16 are shown in Fig. 2. Immunohistochemistry data also reveals a relationship with HPV infection and methylation of p16 gene (Table 3). Non-neoplastic cervical cells showed no expression for p16 protein except the basal cells which normally expressed p16 protein. Similarly, squamous metaplastic epithelial cells also showed immunonegativity against p16 expression. The p16 expression was detectable in the nucleus as well as the cytoplasm of epithelial cells in all positive cases. Cells showing p16 expression also exhibited the morphological features of koilocytosis and papillomatosis caused by HPV infection.
The expression of p16 was found significantly associated with high-risk HPV infection. In HPV positive squamous cell carcinoma, 81.8% cases showed overexpression of p16. In contrast, overexpression of p16 was observed in 29.4% HPV negative cases. Neither HPV nor p16 overexpression was detected in 11.4% squamous cell carcinoma of the cervix cases. Also there was statistically insignificant correlation obtained between p16 expression frequency, menopausal status ( 0.6468), and age of the cancer patients. The association between p16 overexpression and HPV infection is summarized in Table 2.
Discussion
Our results showed that there is aberrant methylation and expression of p16 gene in HR-HPV infected cervical carcinoma patients of India where cancer of cervix is most prevalent among females. We found high percentage (83.8%) of high-risk HPV infection in Indian women suffering from cervical cancer. Our results showed that HPV 16 (95.5%) is more prevalent then HPV 18 (4.5%) in high risk HPV infected population. The etiopathological association of high risk HPV infection with squamous cell carcinoma of the uterine cervix has been well documented in a number of studies [3, 5, 11, 12, 14, 20]. Although there are variations reported in the occurrence of high-risk HPV in cervical cancer in different studies [3, 11, 12] in Indian population, the variations are not statistically significant and there is a high prevalence of HPV infection among Indian women. In India, early marriage, low economic status, illiteracy, and poor hygienic condition [3, 12], an early start of sexual activity and inadequate preventive measures effectively contributes to the high prevalence of HPV infection. This might be the reason as to why cervical cancer is rated the most prevalent cancer in Indian females.
Using p16 monoclonal antibody we found that more than 73% samples of squamous cell carcinoma of cervix exhibited over expression of p16. In normal cells, p16 expression was known to be low at both the mRNA and protein levels [10]. Therefore, expression of p16 may be detectable only when there is overexpression of the protein as in case of cervical cancer cells infected with high-risk HPV type 16/18. Previous studies have shown the overexpression of p16 protein not only in the cervical carcinoma [13, 24, 25] but also in the cervical cancer cell lines and human ectocervical cells infected with HPV 16 and 18 [6]. While, p16 seems to normally act as a negative regulator of cellular proliferation, its inhibitory role is without effect in high-risk HPV infected cell, which can proliferate even in the presence of high levels of p16. No association between aberrant p16 expression and clinical stages of the disease was observed. This was expected since p16 is a negative regulator of the cell cycle, and it has been reported to be overexpressed both in precancerous and invasive cervical cancer lesions especially in HPV positive samples. In this study, menopausal status did not appear to affect p16 expression.
We found that methylation is one of the important mechanisms for the inactivation of p16 gene in HPV associated SCC. This is evident from our findings that promoter CpG island hypermethylation of p16 gene was detected in 20.9% tissue samples of cervical carcinoma and 20 out of 22 samples of hypermethylated promoter, expression of p16 protein was either nil or very low. In different populations hypermethylation of p16 promoter has been demonstrated in 21–61% of cervical carcinomas by methylation-specific PCR [14, 15, 20, 21, 22, 23]. Our results of MSP analysis of the same p16 region in invasive SCC are in close agreement with the above findings. However, there were some reports that showed only 3–6% hypermethylation in the population of Japan and India by using Sma I/Eag I and Hpa II/Msp I restriction enzymes in squamous cell carcinoma of the cervix [16]. Numerous investigations have suggested that hypermethylation of promoter CpG islands located in the proximal promoter region, correlates with transcriptional silencing of the genes including tumor suppressor genes [9, 14, 15, 17, 18]. These methylation changes can be found in virtually every type of human neoplasm, including cervical cancer [14, 15, 18]. Hypermethylation of p16 promoter is a well known mechanism of suppression of the gene transcription [17, 18]. Nuovo et al. through the MSP-ISH technique demonstrated that inactivation of p16 expression has been a result of hypermethylation of the promoter of the p16 gene and it occurs early in the development of squamous cell carcinoma of lung and the cervix [18]. In respect of expression level of hypermethylated gene our study is in agreement with these studies [14, 15]. However, it is in contrast with the study of Nehls et al. [19] that claim p16 hypermethylation does not implicate loss of p16 expression in HPV induced tumours which implies that the E2F can induce transcription even in hypermethylated promoter of p16 gene, that may not be case as we have find in our study that inactivation of p16 gene in HPV associated SCC is via the methylation mechanism. Thus in conclusion we can say that an association between p16 methylation, expression, and HR-HPV infection imply that the compliance of HPV infection and aberration of p16 gene play a synergic effect on development of cervical cancer.
Footnotes
Acknowledgments
Authors are thankful to Council of Scientific and Industrial Research (CSIR) (File No: 09/466(75)/2004-EMR-I) for providing fellowship to the first author and financial support for the study.
References
1.
SchiffmanM.CastleP.E.JeronimaJ.RodruguezA.C. and WacholderS., Human papillomavirus and cervical cancer, Lancet370 (2007), 890–907.
2.
DasB.C.BhartiA.C. and MausumiB., Human papillomavirus & cervical cancer: Looking ahead, Indian J Med Res130 (2009), 210–211.
3.
DasB.C.SehgalA.MurthyN.S.GopalkrishnaV.SharmaJ.K.DasD.K.SinghV. and LuthraU.K., Human papillomavirus and cervical cancer in Indian women, Lancet2(8674) (1989), 1271.
4.
KambA.Shattuck-EidensD.EelesR.LiuQ.GruisN.A.DingW.HusseyC.N. and Cannon-AlbrightL.A., Analysis of the p16 gene (CDKN2) as a candidate for the chromosome 9p melanoma susceptibility locus, Nature Genet8 (1994), 22–26.
5.
KeatingJ.T.InceT. and CrumC.P., Surrogate biomarkers of HPV infection in cervical neoplasia screening and diagnosis, Adv Anat Pathol8 (2001), 83–92.
6.
KhleifS.N.DeGregoriJ.YeeC.OttersonG.A.KayeF.G.NevinsJ.R. and HowleyP.M., Inhibition of cycline D-CDK4/CDK6 activity is associated with an E2F-mediated induction of cyclin kinase inhibitor activity, Proc Natl Acad Sci USA93 (1996), 4350–4354.
7.
BelinskyS.A.NikulaK.J.PalmisanoW.A.MichelsR.SaccomannoG.GabrielsonE.BaylinS.B. and HermanJ.G., Aberrant methylation of p16 (INK4a) is an early event in lung cancer and a potential biomarker for early diagnosis, Proc Natl Acad Sci USA95 (1998), 11891–11896.
8.
SambrookJ.FritshE.F. and ManiatisT., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989.
9.
HermanJ.G.GraffJ.R.MyohanenS.NelkinB.D. and BaylinS.B., Methylation-specific PCR: A novel PCR assay for methylation status of CpG islands, Proc Natl Acad Sci USA93 (1996), 9821–9826.
10.
GeradtsJ.KratzkeR.A.NiehansG.A. and LincolnC.E., Immunohistochemical detection of the cyclin-dependent kinase inhibitor 2/multiple tumor suppressor gene 1 (CDKN2/ MTS1) product p16 in archival human solid tumors: Correlation with retinoblastoma protein expression, Cancer Res55 (1995), 6006–6011.
11.
DasB.C.SharmaJ.K.GopalkrishnaV.DasD.K.SinghV.GissmannL.zur HausenH. and LuthraU.K., A high frequency of human papillomavirus DNA sequences in cervical carcinomas of Indian women as revealed by Southern blot hybridization and polymerase chain reaction, J Med Virol36(4) (1992), 239–245.
12.
MunirajanA.K.KannanK.BhuvarahamurthyV.IshidaI.FujinagaK.TsuchidaN. and ShanmugamG., The status of human papillomavirus and tumor suppressor genes p53 and p16 in carcinomas of uterine cervix from India, Gynecol Oncol69(3) (1998), 205–209.
13.
KlaesR.FriedrichT.SpitkovskyD.RidderR.RudyW.PetryU.Dallenbach-HellwegG.SchmidtD. and von Knebel DoeberitzM., Overexpression of p16INK4A as a specific marker for dysplastic and Neoplastic epithelial cells of the cervix uteri, Int J Cancer92 (2001), 276–284.
14.
DongS.M.KimH.S. and RhaS.H., Promoter hypermethylation of multiple genes in carcinoma of the uterine cervix, Clin Cancer Res7 (2001), 1982–1986.
15.
VirmaniA.K.MullerC.RathiA.Zoechbauer-MuellerS.MathisM. and GazdarA.F., Aberrant methylation during cervical carcinogenesis, Clin Cancer Res7(3) (2001), 584–589.
16.
NakashimaR.FujitaM. and EnomotoT., Murata, Alteration of p16 and p15 genes in human uterine tumours, Br J Cancer80(3–4) (1999), 458–467.
17.
GuoS.TakiT.OhnishiH.PiaoH.TabuchiK.BesshoR.HanadaR.YanagisawaM. and HayashiY., Hypermethylation of p16 and p15 genes and Rb protein expression in acute leukemia, Leukemia Res24 (2000), 39–46.
18.
NuovoG.J.PlaiaT.W.BelinskyS.A. and HermanJ.G., In situ detection of the hypermethylation-induced inactivation of the p16 gene as an early event in oncogenesis, Proc Natl Acad Sci USA96 (1999), 12754–12759.
19.
NehlsK.VinokurovaS.SchmidtD.KommossF.ReuschenbachM.KisseljovF.EinenkelJ.von Knebel DoeberitzM. and WentzensenN., p16 methylation does not affect protein expression in cervical carcinogenesis, Eur J Cancer44(16) (2008), 2496–2505.
20.
SilveiraF.A.AlmeidaG.FurtadoY.SilvaK.S.MaldonadoP.CavalcantiS. and Carvalho MdaG., HPV DNA genotyping and methylation of gene p16 INK4A in cervical LSIL, Exp Mol Pathol98(2) (2015), 308–311.
21.
JhaA.K.NikbakhtM.JainV.CapalashN. and KaurJ., p16 (INK4a) and p15 (INK4b) gene promoter methylation in cervical cancer patients, Oncol Lett3(6) (2012), 1331–1335.
22.
GuptaA.AhmadM.K.MahndiA.A.SinghR. and PradeepY., Promoter Methylation and Relative mRNA Expression of the p16 Gene in Cervical Cancer in North Indians, Asian Pac J Cancer Prev.17(8) (2016), 4149–4154.
23.
LiJ.ZhouC.ZhouH.BaoT.GaoT.JiangX. and YeM., The association between methylated CDKN2A and cervical carcinogenesis, and its diagnostic value in cervical cancer: A meta-analysis, Ther Clin Risk Manag18(12) (2016), 1249–1260.
24.
KanthiyaK.KhunnarongJ.TangjitgamolS.PuripatN. and TanvanichS., Expression of the p16 and Ki67 in Cervical Squamous Intraepithelial Lesions and Cancer, Asian Pac J Cancer Prev.17(7) (2016), 3201–3206.
25.
ZouheirY.FechtaliT. and ElgnaouiN., Human Papillomavirus Genotyping and p16 (INK4a) Expression in Cervical Lesions: A Combined Test to Avoid Cervical Cancer Progression, J Cancer Prev.21(2) (2016), 121–125.