
Abstract
Cervical cancer is strongly related to certain high-risk types of human papilloma virus infection. Breast cancer metastasis suppressor 1 (BRMS1) is a tumor suppressor gene, its expression being regulated by DNA promoter methylation in several types of cancers. This study aims to evaluate the methylation status of BRMS1 promoter in relation to high-risk types of human papilloma virus infection and the development of pre-cancerous lesions and describe the pattern of BRMS1 protein expression in normal, high-risk types of human papilloma virus–infected pre-cancerous and malignant cervical epithelium. We compared the methylation status of BRMS1 in cervical smears of 64 women with no infection by high-risk types of human papilloma virus to 70 women with proven high-risk types of human papilloma virus infection, using real-time methylation-specific polymerase chain reaction. The expression of BRMS1 protein was described by immunohistochemistry in biopsies from cervical cancer, pre-cancerous lesions, and normal cervices. Methylation of BRMS1 promoter was detected in 37.5% of women with no high-risk types of human papilloma virus infection and was less frequent in smears with high-risk types of human papilloma virus (11.4%) and in women with pathological histology (cervical intraepithelial neoplasia) (11.9%). Methylation was detected also in HeLa cervical cancer cells. Immunohistochemistry revealed nuclear BRMS1 protein staining in normal high-risk types of human papilloma virus–free cervix, in cervical intraepithelial neoplasias, and in malignant tissues, where staining was occasionally also cytoplasmic. In cancer, expression was stronger in the more differentiated cancer blasts. In conclusion, BRMS1 promoter methylation and aberrant protein expression seem to be related to high-risk types of human papilloma virus–induced carcinogenesis in uterine cervix and is worthy of further investigation.
Introduction
Cervical cancer is the fourth most common cancer in women globally and the seventh overall, with an estimated 528,000 new cases in 2012, 1 although prevention options, that is, molecular/cytological screening and immunization, are available and have been proven to hold significant value.2–4 The main etiological cause of cervical cancer is the infection by certain types of human papilloma virus (HPV), characterized as high-risk strains (high-risk types of human papilloma virus (HR-HPV)). Cytology, colposcopy, histology, and molecular HPV testing contribute to the diagnostic procedure, with well-discussed advantages and disadvantages, and remain the mainstay of diagnosis, while the need for better characterization of early lesions and understanding of the virus-related carcinogenetic process remains crucial.
DNA methylation, defined as the covalent addition of a methyl group at the 5-carbon of the cytosine ring (5-methylcytosine), is a well-characterized epigenetic mechanism of DNA modification, contributing to gene expression regulation. 5 Methylation is related to a variety of normal cellular functions such as silencing of transposable elements and defense against viral sequences.6,7 It is also associated with reactivation of cancer-associated and tumor suppressor genes. 5 Recently, much attention is raised to the evaluation of DNA methylation as a potential cancer and/or screening biomarker but also as a tool to unravel the carcinogenic process.8–11
Breast cancer metastasis suppressor 1 (BRMS1) is a tumor suppressor gene encoding predominately a nuclear protein.12,13 BRMS1 has been shown to regulate the expression of several metastasis-associated genes, leading to suppression of metastasis without blocking orthotropic growth, 14 whereas its cytoplasmic shuttling has been related to enhanced mitosis and more aggressive tumor behavior. 12 BRMS1 has been shown to be down-regulated by DNA methylation in breast, non–small cell lung, and other cancers. In addition, BRMS1 promoter methylation has been shown to be associated with poor prognosis and reduced disease-free survival.9,15
Although the role of BRMS1 has been studied in many types of cancers and in vitro models, there is no information available on its expression and function in uterine cervix and in cervical cancer. This knowledge gap is addressed in this study. Our aims were (a) to describe the methylation status of BRMS1 promoter in relation to HR-HPV infection and preclinical lesions in a clinically diagnosed cohort by methylation-specific quantitative polymerase chain reaction (qMSP) and (b) to describe the pattern of protein expression in biopsies from normal, HR-HPV-infected pre-cancerous, and malignant cervical epithelium.
Materials and methods
Study groups
Our study was approved by the Ethics review board of the University General Hospital of Alexandroupolis/Greece. Residual archive total DNA material from cervical smears previously collected for HPV molecular diagnosis (n = 134) for the purpose of our former HEllenic Real life Multicentric cErvical Screening (HERMES) study was used. 16 Between them, 64 had shown no infection by HR-HPV (individual identification of 16 and 18, other high-risk strains, that is, 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 66, and 68) (mean age, 35.31 ± 9.081) and 70 of them had proven infection with varying cervical intraepithelial neoplasia (CIN) stage (mean age, 33.39 ± 9.567). These samples had also been previously tested for the presence of herpes simplex virus (HSV), Epstein–Barr virus (EBV), and cytomegalovirus (CMV) infection. In addition, archived formalin-fixed, paraffin-embedded cervical tissue specimens obtained for routine histopathology clinical assessment were used, diagnosed for squamous cervical carcinoma (n = 10) (mean age, 55.0), squamous intraepithelial lesion (varying CIN stage) (n = 13, Table 2) (mean age, 36.8), or characterized as normal HPV-free cervix tissue (n = 2) (mean age, 49.2). Clinical and demographic data from subjects enrolled were also available (Tables 1 and 2). Archived placental formalin-fixed, paraffin-embedded tissue sections were also used as controls in immunohistochemistry (as suggested by antibody manufacturer Abcam, Cambridge, UK). We also investigated the methylation status of the BRMS1 promoter in HeLa human cervical cancer cells, kindly donated by Prof. P. Mavromara, Department of Molecular Biology and Genetics, DUTH.
Demographic and clinical characteristics of the cervical smear study population.

HPV−VE: HPV negative; HPV+VE: HPV positive; SD: standard deviation; HPV-OHR: HPV-Other (no 16 and 18) high risk; CINI, CINII, CINIII: cervical intraepithelial neoplasia grade I, II, III; ASCUS: atypical squamous cells of undetermined significance; LGSIL: low-grade squamous intraepithelial lesion; HGSIL: high-grade squamous intraepithelial lesion; HSV: herpes simplex virus; CMV: cytomegalovirus; EBV: Epstein–Barr virus.
Demographic and clinical characteristics of the formalin-fixed, paraffin-embedded cervical tissue specimens.

SD: standard deviation; SCCC: squamous cell cervical carcinoma; CINI, CINII, CINIII: cervical intraepithelial neoplasia grade I, II, III.
DNA extraction
Genomic DNA had been previously extracted from cervical liquid-based cytology (LBC) specimens using the automated system of the Cobas 4800 HPV PCR by Roche, which was also used for their subsequent analysis for the detection of the genome of 14 HR-HPV types. Total genomic DNA was also isolated from HeLa cell line using the Qiagen DNA Mini Kit (Qiagen, Hilden, Germany). In addition, DNA was extracted from paraffin-embedded cervical abnormal and adjacent tissue as directed by standard eosin staining, using the High Pure PCR Template Preparation Kit (Roche Diagnostics, Germany) following the manufacturer’s instructions. Three samples were excluded from further analysis due to low quality of DNA, as assessed by ACTB quantitative polymerase chain reaction (qPCR) assay. The extracted DNA was stored at −20°C until further use.
Sodium bisulfite conversion
Bisulfite conversion was performed by EZ DNA Methylation-Gold™ Kit (Zymo Research Co., Orange, CA, USA) as described by the manufacturer. During conversion, all unmethylated, but not methylated, cytosines are converted to uracil. One microgram of DNA was used for each conversion reaction, and after conversion, DNA was eluted in 50 µL of elution buffer. The converted DNA was stored at −80°C until use.
qMSP
The KAPA SYBR Fast Master Mix PCR Kit (KAPA Biosystems, London, UK) was employed for qMSP and primers specific for fully methylated and fully unmethylated BRMS1 promoter sequences. 17 A methylation-independent assay with non-CpG bearing primers for the β-actin gene (ACTB) were used in order to verify sufficient DNA quality and quantity of converted DNA and to normalize our results. Primer sequences and related references are shown in Table 3. All primers were obtained from TIB MOLBIOL (Berlin, Germany). For each 20 µL PCR reaction, 4 µL of SB-converted DNA, 10 µL of KAPA SYBR Fast Master Mix, 0.4 µL of each forward and reverse primer for mBRMS1 and uBRMS1, 0.3 µL for ACTB primer pair, 0.8 µL MgCl2, and DNase-free water were added to a final volume of 20 µL. Quantitative PCR was performed using the RotorGene 6000 (Qiagen) with the following cycling parameters: 95°C for 5 min, followed by 45 cycles of 95°C for 20 s, annealing temperature: mBRMS1 and uBRMS1 59°C for 20 s, ACTB 62°C for 20 s, 72°C for 20 s. Melting curve was obtained from 55°C to 99°C. Correct length of PCR products were verified by gel electrophoresis. Samples with a quantification cycle (Ct) >31 for ACTB gene were excluded from further analysis.
Primer sequences for qMSP for BRMS1 and ACTB genes.

mBRMS1: methylated breast cancer metastasis suppressor 1; uBRMS1: unmethylated breast cancer metastasis suppressor 1; ACTB: beta-actin.
Specificity and cross reactivity of methylated BRMS1 primers and unmethylated BRMS1 primers were evaluated using unconverted genomic DNA (gDNA), converted DNA from human placenta (0% methylated) (Sigma-Aldrich, St. Louis, MO, USA) and a M. SS1 (CpG Methyltransferase) (New England Biolabs, Massachusetts, USA) treated DNA (100% methylated). qMSP sensitivity for BRMS1 gene was evaluated using serial dilutions of 100% and 0% methylated, SB-converted DNA (100%, 50%, 10%, 5%, 1%, and 0.5%). Efficiencies of mBRMS1 and ACTB assays were evaluated using serial dilutions of 100% methylated SB-converted DNA in H2O (standard curve).
Results were calculated using the Rotor-Gene Series Software 1.7 (Qiagen). Analysis was performed according to the RQsample (Relative Quantification) = 2−ΔΔCT method. 18 Specifically, ΔΔCT values were generated for each target after normalization by ACTB values and using 1% methylation as calibrator. Negative and positive control samples of 0% and 100% methylated SB-converted DNA were included in every run.
Immunohistochemistry
Immunohistochemistry was conducted as previously described. 19 Tissue specimens were fixed in formalin and embedded in paraffin, according to standard procedures for clinical diagnosis. Four-micrometer sections (4 µm) of representative blocks were deparaffinized, rehydrated, and treated with 0.3% H2O2 for 5 min in methanol to prevent endogenous peroxidase activity. After washing, slides were incubated for 1 h with the primary rabbit anti-human BRMS1 monoclonal antibody (clone EPR7202) (Abcam) at room temperature (RT), diluted 1:400 in antibody diluent (DAKO Denmark A/S, Denmark). Optimal primary antibody incubation and concentration were determined by serial dilutions on placental tissue (positive control). Control slides were incubated for the same period with normal mouse serum IgG. Immunostaining was detected by the Dako REAL TM EnVision TM Detection System, Peroxidase/DAB+, Rabbit/Mouse kit (DAKO), using a standard streptavidin/biotin detection method, following the instructions of the manufacturer. Finally, bound antibody complexes were stained for 5 min with 0.05% diaminobenzidine, counterstained with Mayer’s hematoxylin, and mounted and observed under a Nikon Eclipse 50i microscope.
Statistical analysis
Initially, Kolmogorov–Smirnov test was applied, in order to check for normality in distribution. Due to lack of normality, non-parametric statistics were used, for example, Mann–Whitney analysis for comparison between groups. Additionally, for comparison between categorical variables, the chi-square test was used. The median age was used in order to divide groups according to age (33 years). Logistic regression analysis was applied to estimate whether BRMS1 methylation could be predicted by different variables. Statistical significance was set at p value <0.050 (IBM SPSS 19.0 statistical software; IBM Corp., 2010, IBM SPSS Statistics for Windows, Version 19.0, Armonk, NY, USA).
Results
Validation of qMSP
The unmethylated DNA control (placental DNA) yielded product only with unmethylated BRMS1 primers (melting temperature 78°C; product length 130 bp), whereas methylated DNA control yielded product only with methylated BRMS1 primers (melting temperature 80°C, product length 130 bp). ACTB assay yielded product with melting temperature 77.5°C and product length 77 bp. No product was observed either with methylated or unmethylated primers, when we used unconverted gDNA confirming specificity of reactions. Concerning the analytical sensitivity of qMSP, assay could detect methylation in a 0.5% BRMS1 methylated sample and efficiency was 103%.
Promoter methylation of BRMS1 in cervical specimens
We initially assessed promoter methylation of BRMS1 in cervical smears of 64 women with no infection by HR-HPV and 70 women with proven infection. Methylation was detected more often in cervical smears of women with no HR-HPV infection than in infected women (p < 0.001, χ2 = 12.502) (Figure 1(a)). Less methylation was detected in the subgroup of women infected by HPV-18 (p = 0.040, χ2 = 4.224) and other high-risk HPV (HPV-OTH) (p = 0.015, χ2 = 5.872). Samples with pathological histology (CIN) were found methylated less frequently than samples with no pathological findings (p = 0.007, χ2 = 7.351), but cytological type (atypical squamous cells of undetermined significance (ASCUS), low-grade squamous intraepithelial lesion (LGSIL), high-grade squamous intraepithelial lesion (HGSIL) did not correlate to methylation (Figure 1(b) and (c)). Interestingly, methylation of BRMS1 promoter was present more often in older (>33) than younger (≤33) women (p = 0.013, χ2 = 6.236) (Figure 1(d)), but this difference, although present in the non-infected group, could not be detected in the HR-HPV-infected women. BRMS1 promoter methylation was present more often in women who had not been previously pregnant than in those who have given birth at least once (p = 0.026, χ2 = 4.968). However, logistic regression analysis assessing the effect of age or pregnancy history on BRMS1 methylation showed no significant association (age: odds ratio (OR) = 1.05; 95% confidence interval (CI) = 0.996–1.108; p = 0.071; pregnancy: OR = 0.641; 95% CI = 0.222–1.852; p = 0.411). 20 Finally, BRMS1 promoter methylation was not significantly correlated with smoking habits, abortions, test papanicolaou (PAP) frequency, usage of condom, and infection by HSV, CMV, and EBV viruses.
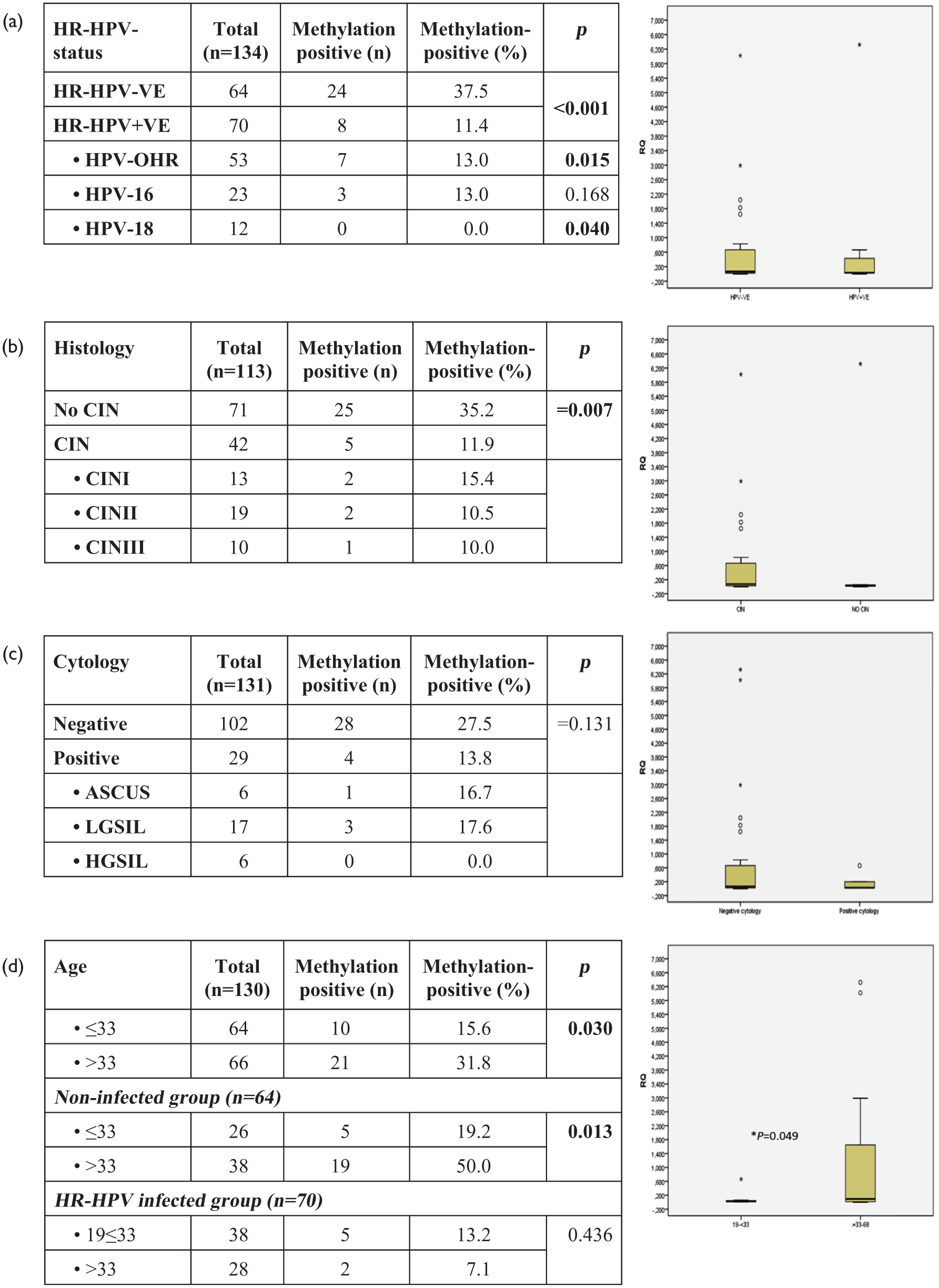
The status of BRMS1 promoter methylation in cervical smears as assessed by qMSP in relation to (a) HR-HPV infection, (b) histology diagnosis, (c) cytological diagnosis, and (d) age.
Mean RQ values (%), reflecting levels of methylation in positive samples in every group, were calculated. Analysis showed that levels of methylation were not associated with HR-HPV infection, being 71 ± 139 and 91 ± 220 RQ (%) in HPV negative and HR-HPV-infected samples, respectively (p = 0.749), 70 ± 136 and 128 ± 282 in no CIN and CIN samples, respectively (p = 0.516), and 83 ± 168 and 23 ± 0.30 RQ (%) in groups with normal cytology diagnosis and pathological findings, respectively (0.934). However, the wide range of values detected within each group might have not allowed differences to emerge. Still, levels were marginally correlated in a statistically significant way with women’s age, being higher in older women than younger women (>33–68 RQ (%) = 116 ± 192; 19–33 RQ (%) = 8 ± 19; p = 0.049).
Next, we investigated promoter methylation of BRMS1 in a small panel of formalin-fixed, paraffin-embedded cervical tissue specimens (2 normal, 10 invasive squamous cervical carcinomas, 1 CINI, 5 CINII, 5 CINIII, and 12 adjacent non-malignant tissues). BRMS1 promoter methylation was detected only in 4 out of 10 cancers (0/3 differentiation grade II, 2/2 grade II/III, and 2/5 grade III), whereas all the other samples were unmethylated. Notably, BRMS1 promoter methylation was also detected in HeLa cervical cancer cells.
Immunohistochemical analysis of BRMS1 protein expression in cervical biopsies
Immunohistochemistry revealed clear nuclear BRMS1 staining in both normal HR-HPV free tissues, which was detected in almost all the width of squamous multilayer epithelium of ectocervix and in the monolayer cylinder epithelium of endocervix, but also in cervical stroma and inflammatory cells (Figure 2(c) and (d)). BRMS1 protein was expressed in CINs of all histological types (CIN I, II, and III), showing a similar pattern to the normal tissues, except that the superficial layer of the squamous epithelium in the ectocervix was negative (Figure 2(e)–(g)). In malignant tissues, strong nuclear BRMS1 staining was found in the well-differentiated cancerous blasts (Figure 2(h)); however, cytoplasmic staining could also been occasionally detected (Figure 2(k)). Expression in cancer cells of well-differentiated cancerous blasts was stronger than the surrounding adjacent tissue (Figure 2(h)). However, expression was weaker in less differentiated blasts, being practically absent (<10%) in grade III cancers (Figure 2(i) and (j)).

Immunohistochemical staining of BRMS1 in (a) placental tissue (positive control), (b) normal cervical epithelium following staining without primary antibody (negative control), (c) normal ectocervix, (d) normal endocervix, (e) cervical intraepithelial neoplasia grade (CIN) I, (f) CIN II, (g) CIN III, (h) cancer blast of high (grade I), (i) moderate (grade II), and (j) low (grade III) differentiation, (k well-differentiated cancer blast with nuclear and occasionally cytoplasmic staining.
Discussion
It is well documented that HR-HPV is connected to the initiation of cervical carcinogenesis.21,22 In addition, there are many studies supporting that infection by HR-HPV is associated with DNA methylation related to the development of cervical cancer.23,24 For instance, it has been proposed that oncoproteins E6 and E7 of HR-HPV increase the expression and activity of the methyltransferase 1 (DNMT1). In addition, several researchers suggest that methylation biomarkers could be of clinical value as cervical screening tools24–27 or/and in therapeutic monitoring. 28 On the other hand, BRMS1 is a metastasis suppressor that is down-regulated in several cancers.29–31 The mechanism by which BRMS1 lose its expression is still unknown; however, it has been suggested that DNA methylation could cause its down-regulation.13,29 In our present exploratory study, we addressed for the first time the methylation status of BRMS1 promoter in normal uterine cervix and we correlated it with HR-HPV infection and the development of pre-malignant lesions. A cohort of cervical smears from a female population previously tested for HR-HPV infection by molecular techniques, cytology and histology, was employed. We also first report BRMS1 protein expression in a different small cohort of healthy, pre-cancerous, and cancerous cervical biopsies.
Our results demonstrate that BRMS1 promoter was found methylated in 37.5% of cervical smear samples in the population of non-HR-HPV-infected healthy women. This was already a surprising high methylation percentage, as other studies report low to zero percentages of BRMS1 methylation in circulating DNA from healthy individuals or in adjacent to cancerous tissues such as breast, lung, and laryngeal mucosa.9,15,32,33 Still, as methylation is a predominately tissue-specific event,34–36 our observations have to be attributed to the specific histological microenvironment of cervical tissue.
Methylation was less frequently detected in those women with proven infection by HR-HPV (11.4%) than in non-HR-HPV-infected women. Methylation status was related to the type of HR-HPV, being less frequent in HPV-18 and HPV-OHR. Moreover, BRMS1 methylation was also significantly correlated to histology, being found less frequently in samples with pathological histology (CIN) (11.9%) than samples with no pathological findings (35.2%). Methylation was not statistically correlated to CIN stage, probably due to the small number of positive cases in each group (2, 2, and 1 for CIN I, II, and III, respectively). Methylation was also not correlated to cytological diagnosis. In fact, our results showed that cytology failed to detect cervical pathology in women found positive for HR-HPV and had pathological histology result (CIN). Percentages of a pathological result in our study group were 52%, 37%, and 22% for HR-HPV DNA test, histology, and cytology, respectively. This was also concluded by our previous HERMES study, in a population of 4009 women, 16 demonstrating that HPV testing has much better sensitivity than cytology to identify high-grade cervical lesions with slightly lower specificity and that HPV testing with individual HPV-16/HPV-18 genotyping could represent a more accurate methodology for primary cervical cancer screening in comparison to LBC. Lack of statistical correlation to the methylation status to cytology could therefore be due to the false-negative cases of this diagnostic method.
Our results indicate an HR-HPV-related demethylation of BRMS1 promoter and are differing to those findings supporting an HPV-induced DNA methylation of tumor/metastasis suppressor genes contributing to carcinogenesis. In most cases, methylation signatures are used from tumor suppressor genes that will get more intensely methylated during cancer progression in order to switch off expression. Here, in contrast, the loss of methylation in a gene that may be supporting cellular transformation and metastasis seems to be associated with cancer initiation. It could be postulated that during the initial phase of carcinogenesis, host tissue mechanisms are responsible for demethylation of BRMS1 in certain patients leading to protein expression, as a rescuing response to HR-HPV infection. In fact, as a metastasis suppressor gene, multiple anti-cancer functions have been attributed to BRMS1, such as enhancement of apoptosis, cell adhesion, immune recognition as well as inhibition of cell survival, angiogenesis, invasiveness, and immigration.37–41 Epigenetic regulation in cancer initiation and development via demethylation has been studied 42 and there are previous reports of increased gene expression as a result of demethylation, in support to our findings. For example, Akiyama et al. 43 have shown that the metastasis suppressor gene Maspin is hypomethylated in intestinal metaplasia as compared to gastric normal epithelia, resulting in protein expression. In addition, the tumor suppressor genes P27, P53, and RB1 were found hypomethylated more frequently in odontogenic myxoma than in dental pulp. 44
Interestingly, BRMS1 was found methylated in the cervical squamous carcinoma cell line HeLa, which could therefore serve as an in vitro model for the study of BRMS1 methylation in carcinogenesis in uterine cervix by future functional demethylation studies.
Methylation of BRMS1 was also related to women’s age, as it was found more frequently in older women of our study population. This difference was also significant in the healthy individuals group, but could not be detected in the HR-HPV-infected group; therefore, both older and younger infected women were found more rarely methylated. However, logistic regression analysis in the total population and in subgroups showed no correlation between age and BRMS1 promoter methylation, weakening the significance of these findings. Age has been shown to be related to the methylation status of other genes too, such as FAM19A4, interfering with the sensitivity and specificity of its clinical performance as a biomarker for the triage of HR-HPV-infected women. 45
No other correlation was found between BRMS1 promoter methylation in uterine cervical tissue with abortions, test PAP frequency, usage of condom, and infection by HSV, CMV, and EBV viruses, although methylation was present more often in women who had not been previously pregnant. As far as smoking habits, again we could not find any correlation, such as that previously was reported in the lung of non–small cell lung cancer (NSCLC) patients. 33
Descriptive immunohistochemistry revealed that BRMS1 protein was expressed in the nucleus of normal HR-HPV-free cervical epithelium, in almost all the width of squamous multilayer epithelium of ectocervix and in the monolayer cylinder epithelium of endocervix, and also in stromal and inflammatory cells. In addition, intraepithelial neoplasias of all stages (CIN I, II, and III) showed a positive immunostaining, with a similar expression pattern except for the superficial layers of ectocervix where staining was less extended. These results are in accordance with what was previously reported for skin, where nuclear BRMS1 expression was found more often and was higher in benign nevi as compared to primary melanoma. 46 On the other hand, in malignant tissue, BRMS1 protein expression was associated with pathological grade, being reduced in less differentiated cancerous blasts and even absent in grade III undifferentiated blasts. Our results indicate a correlation between BRMS1 down-regulation and cervical cancer progression and aggressiveness, as it would be expected for a tumor/metastasis suppressor gene related to metastatic potential, and are in agreement with studies in glioma, 41 melanoma, 46 nasopharyngeal carcinoma, 47 and nasal and paranasal sinus carcinomas 48 showing correlations to stage and patient survival. Expression in cancer cells of blasts was stronger than the surrounding adjacent tissue in agreement to what reported in breast, where BRMS1 protein and messenger RNA (mRNA) were higher in cancer than the adjacent normal breast epithelium. 12
BRMS1 protein was found predominately in the cell nucleus in all healthy, pre-cancerous, and malignant biopsies, but also occasionally in the cytoplasm of cancer cells. It has been reported that in breast cancer, BRMS1 localization can shift from nuclear to cytoplasmic and that this phenomenon is related to more aggressive tumors with lower estrogen receptor (ER), lower progesterone receptor (PR), and higher Ki-67 expression and to poorer overall patient survival. 12 This was, however, in contrast to what was shown in melanomas, where cytoplasmic BRMS1 appears to restrict melanoma progression, whereas nuclear expression is associated with cell invasion. 46 A larger cohort of cervical cancer biopsies would be necessary to clarify the relation of nuclear/cytoplasmic BRMS1 protein index to uterine cervix cancer aggressiveness, metastasis, and patient survival, thus contributing to the understanding of subcellular shifting and function of this protein.
In our methylation analysis of BRMS1 promoter in formalin-fixed, paraffin-embedded cervical tissue specimens, methylation was detected only in 4/10 squamous cell carcinoma cancers that were all of advanced grade. This percentage cannot be directly compared to our findings in the cervical smear cohort, as a different methodological approach was employed (biopsies vs smears) and the study group was very small, not suitable for statistical correlations. However, these results were consistent to the immunohistochemistry findings, showing less to no protein expression in the less differentiated, higher grade cancers and suggest that protein expression down-regulation might be the result of gene promoter methylation in these tissues. Previous studies have shown BRMS1 promoter methylation in breast and lung cancer tissues and patient cell-free DNA, as well as associations with worst prognosis.9,15
Overall, our study in the cervical biopsies showed that BRMS1 is strongly expressed in CIN and well-differentiated cancerous blasts, whereas expression was weaker in less differentiated/more aggressive blasts. DNA extracted from these tissues showed BRMS1 promoter methylation in some cases, leading to the conclusion that this may be the responsible mechanism for protein down-regulation. These findings are easily interpreted, as the metastasis suppressor protein is expressed in this as in other cancer types, and the lack of its expression is related to aggressiveness and most probably metastasis and worst prognosis. Unfortunately, no follow-up data were available for these patients, which could reveal a prognostic value.
In conclusion, our results suggest the BRMS1 promoter methylation status in uterine cervical smears is related to HR-HPV infection and CIN diagnosis, thus its clinical significance in the prognosis of cervical pre-cancer in HR-HPV-infected women needs to be evaluated. BRMS1 protein is expressed in cell nuclei of healthy uterine cervices and in pre-cancerous lesions, whereas in cancers its expression is also cytoplasmic and is related to cell differentiation, being absent in the low differentiated cancer blasts. It is supported by our results that this could be related to gene promoter methylation, leading to protein expression down-regulation and loss of a prophylactic function of the protein, higher mitotic index, and greater metastatic potential. Although taken together, our results do not consolidate a causal relationship, BRMS1 promoter methylation seems to present a protein expression regulatory level in uterine cervix related to HR-HPV infection and carcinogenesis, worthy of further investigation.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.