
Abstract
In this study, the phylogeny of species in the genus Syncirsodes Butler, based on morphological characters, was determined. Species included were: S. distictaria, S. hyadesi, S. primata, and S. straminea. Twenty-seven morphological characters of adult external and internal structures were used, with principal emphasis on male and female genitalia and metafurcasternum. A parsimonious cladogram (L = 51, CI = 0.94, RI = 0.83) was made. The phylogenetic hypothesis suggests two groups of sister species: S. distictaria + S. straminea and S. hyadesi + S. primata.
Introducción
A pesar de los muchos trabajos realizados en el campo de la sistemática de insectos de América del Sur Austral (Needham y Bullock 1909; Monrós 1953; Davis 1986; Roig-juñent 2000; Domínguez 2005; Parra & Hernández 2010; entre otros), muchos grupos aún permanecen relativamente poco conocidos, existiendo en la actualidad pocos taxónomos en relación a la enorme diversidad de especies conocidas y por conocer en esta región (Domínguez et al. 2008). Los lepidópteros no escapan a esta realidad.
El actual conocimiento sobre los lepidópteros en Chile data de mediados del siglo XIX con los primeros naturalistas extranjeros (Blanchard 1852; Mabille 1855; Felder et al. 1875; Butler 1882; Prout 1910; Rindge 1971; Rindge 1986). Posteriormente, varios investigadores chilenos han contribuido con revisiones específicas a nivel de familias y grupos menores, enfatizando el estudio de los distintos estados de desarrollo de las especies (Ureta 1956; Angulo y Weigert 1975; Parra & Santos-Salas 1992; Parra 1996; Angulo et al. 2004; Parra & Hernández 2010). Los lepidópteros son uno de los órdenes de insectos más diversos en Chile (Parra 1995) y el mundo (Heppner 1991; Nieukerken et al. 2011). Muchas de sus especies se conocen a través de los caracteres morfológicos externos de los imagos, referidos principalmente a las alas (venación, forma, y maculación), otros relacionados con las antenas, piezas bucales (principalmente palpos y proboscis), presencia o ausencia de ocelos, forma de patas, tamaño, color general del adulto y las armaduras genitales (Borror et al. 1989). En la actualidad, la mayor importancia la tienen las armaduras genitales, principalmente porque la morfología de estas estructuras se considera un rasgo fuertemente especie-específico (hipótesis "llave-cerradura") pues conlleva de manera explícita mecanismos de aislamiento reproductivo (Shapiro y Porter 1989; Scoble 1995). De este modo, la separación en distintos géneros y la proposición de nuevos taxa en Lepidoptera, frecuentemente, se sugieren sobre la base de las diferencias morfológicas en la armadura genital de los adultos de ambos sexos (Scoble 1995; Parra y Pascual-Toca 2003), debido a que en muchos casos las estructuras morfológicas externas asociadas a la cabeza, cuerpo y alas corresponden a homoplasias (convergencias y paralelismos), que hacen indistinguibles los diferentes taxa y no muestran las verdaderas relaciones genealógicas entre ellos.
El metafurcaesterno, también aparece como una estructura con valor taxonómico y filogenético en los lepidópteros y su validez ha sido comprobada en los grupos Dytrisia (Brock 1971), microlepidópteros (Davis 1986) y nóctuidos (Angulo et al. 1986). En los Palaephatidae (Davis 1986) este carácter tiene un patrón característico para los distintos representantes de la familia, aportando atributos diagnósticos que permiten discriminar géneros y especies. El metafurcaesterno, endoesternito del metatórax, sirve como estructura de soporte para la inserción de los diversos músculos que se encuentran asociados a las alas (Ross 1964); de esta manera existe una relación directa entre el desarrollo del metafurcaesterno y un mayor desarrollo del vuelo, por ello hay una serie de estados asociados a esta estructura que permitirían establecer diferencias a distinto nivel taxonómico (Angulo et al. 1986).
En Chile, los geométridos (Lepidoptera: Geometridae) están siendo estudiados taxonómica y sistemáticamente con el objetivo de corroborar las clasificaciones actuales, basadas únicamente en caracteres morfológicos externos, los cuales han generado clasificaciones parafiléticas y polifiléticas, que han ocasionado confusión entre géneros y especies con la consecuente creación de una larga lista de sinonimias (Parra y Pascual-Toca 2003). De las seis subfamilias que conforman la familia Geometridae, cinco están presentes en nuestro país, con 87 géneros, y un total aproximado de 320 especies descritas. Ennominae es la subfamilia más diversa a nivel Neotropical (Heppner 1991; Scoble 1995; Nieukerken et al. 2011), y también lo es en Chile (Parra 1995, 1999a; Parra y Vargas 2000). Se han realizado revisiones de algunos géneros de esta subfamilia, entre ellos Psilaspilates (Parra 1999a), Euclidiodes (Parra 1999b), Neorumia (Parra y Vargas 2000), Oratha y Hasodima (Parra y Pascual-Toca 2003) y además, la tribu Lithinini (Parra y Hernández 2010); contribuyendo al correcto reconocimiento de sus especies y al ordenamiento taxonómico de estos grupos. Esto corresponde a una pequeña fracción del total de géneros señalados para Chile, restando aún una serie de revisiones que permitan tener un mejor conocimiento de los distintos taxa que conforman la subfamilia Ennominae.
Dentro de la inmensa diversidad observada en este grupo, Syncirsodes se destaca porque sus especies presentan una gran variabilidad inter e intraespecífica, lo que llevó a su propio autor (Butler 1882, 1893) a confundir las distintas especies. Syncirsodes fue creado por Butler (1882) para la especie chilena Syncirsodes straminea, basándose en la morfología externa. Felder y Rogenhofer (1875) describen una nueva especie para Chile Paragonia deustata, muy similar a S. straminea. Posteriormente, Butler (1882) describe cinco especies de Paragonia incluyendo la especie de Felder y Rogenhofer (1875). Mabille (1885) describe dos especies más en los géneros Hippoplectis y Ennomos, muy cercanas y similares a Paragonia. Más tarde, Butler (1893) crea el género Anisogonia, en el cual incluye las especies determinadas como Paragonia. Warren (1894), redescribe el género Anisogonia agregando a éste las especies Aspilates? primata Walker (1862) y Macaria subornata Walker, 1863. Scoble (1999) en su catálogo mundial de la familia Geometridae, señala que el género Ennada es el sinónimo senior del género Anisogonia e incluye dentro de éste a las especies: E. deustata, E. distictaria, E. flavaria, E. hyadesi, E. primata y E. subornata. Por último, Pitkin (2002) en su revisión de los géneros neotropicales de Ennominae, indica que Anisogonia es el sinónimo junior del género Syncirsodes y que Ennada es un género monotípico que pertenecería a la subfamilia Larentiinae. De esta manera, las especies incluidas en Ennada por Scoble (1999) son incorporadas a Syncirsodes (Pitkin 2002): S. deustata, S. distictaria, S. hyadesi, S. primata, S. straminea y S. subornata. Finalmente (Bocaz y Parra 2005) consideran a S. deustata como sinónimo junior de S. primata, señalando que el género está compuesto por cinco especies: S. primata (Walker, 1862), S. straminea Butler, 1882 (Fig. 1), S. distictaria (Mabille, 1885), S. hyadesi (Mabille, 1885) y S. subornata (Walker, 1862-1863).

Imagos de S. straminea en vista dorsal. A. Macho; B. Hembra.
Sin embargo, a pesar de los aportes de Pitkin (2002), y Bocaz y Parra (2005) aún es importante esclarecer esta clasificación mediante la sistemática filogenética considerando el argumento de Scoble (1995) respecto a la importancia de la genitalia para la clasificación de los Lepidoptera y teniendo en cuenta el valor del metafurcaesterno como carácter diagnóstico, permitiendo evaluar si el género Syncirsodes es una entidad natural monofilética cuyas especies poseen una historia evolutiva común, y establecer las relaciones de parentesco entre las distintas especies que lo conforman, sustentadas sobre apomorfias morfológicas derivadas de las armaduras genitales del macho y la hembra (Fig. 2), así como del metafurcaesterno, identificando los caracteres que poseen un mayor valor filogenético y aquellos que permiten discriminar entre especies como caracteres diagnósticos.
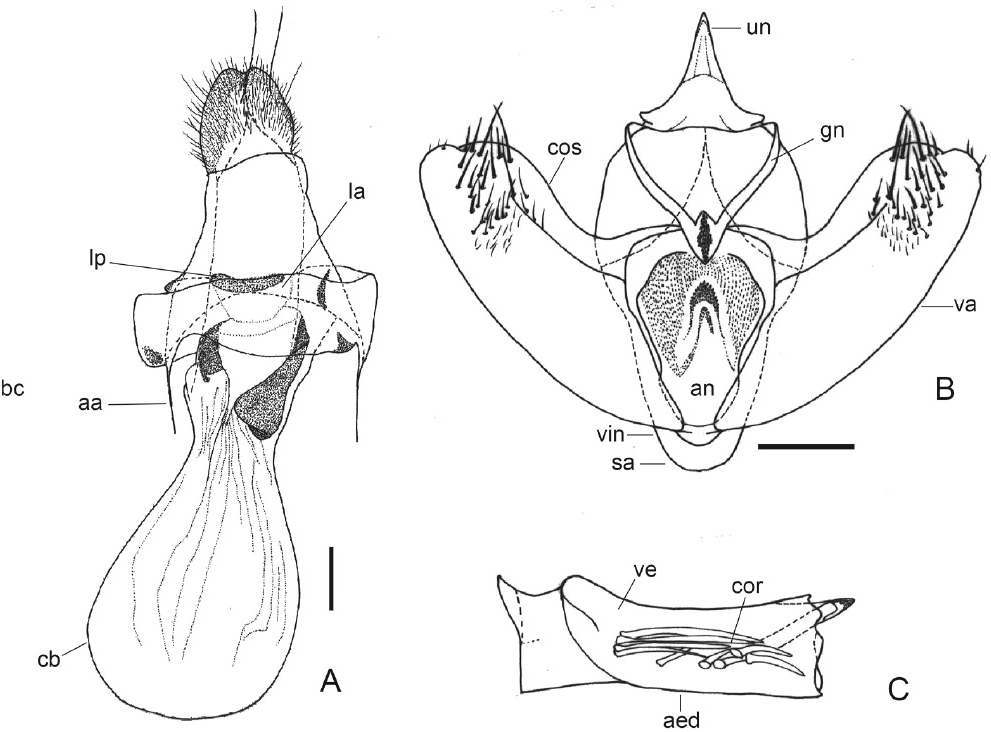
Genitalia del macho y hembra de S. primata. A. Genitalia del macho en vista ventral; B. Phallus en vista lateral; C. Genitalia de la hembra en vista ventral. aa apófisis anteriores, aed phallus, an anellus, bc bursa copulatrix, cb corpus bursae, cos costa de las valvas, cor cornuti, gn gnathos, la lamella antevaginalis, lp lamella postvaginalis, sa saccus, un uncus, va valva, vi vinculum. Escala 1 mm.
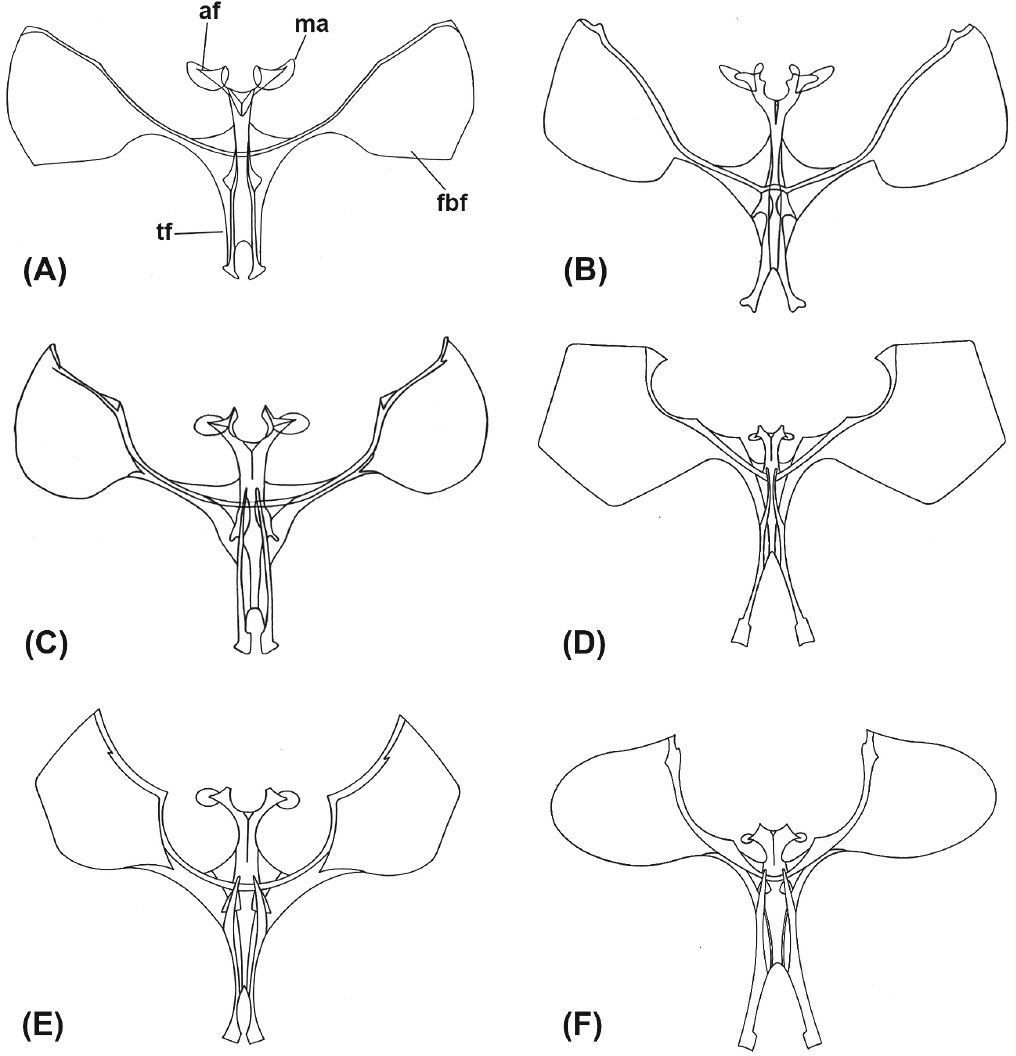
Metafurcaesterno de A: Cotophoenissa dibapha (af: ápice de la furca, tf: tallo de la furca, ma: membrana del ápice y fbf: forma del brazo de la furca) B: Tetracis chilenaria, C: S. primata, D: S. straminea, E: S. hyadesi y F: S. distictaria.
Materiales y métodos
Se examinaron 187 individuos correspondientes a las cuatro especies que comprenden Syncirsodes, a 115 de ellos se les realizaron disecciones para obtener la genitalia de machos y hembras, así como para extraer el metafurcaesterno. Para el análisis de genitalia se utilizaron las técnicas de Parra (1991) y la nomenclatura de Klots (1970). Para el análisis y terminología del metafurcaesterno se siguió a Angulo et al. (1986) y Grassé (1951). Para el análisis filogenético se utilizaron como grupos externos a los géneros Tetracis y Catophoenissa, puesto que ambos se encuentran claramente emparentados con Syncirsodes al compartir características morfológicas externas e internas, poseer una distribución similar y pertenecer a la misma subfamilia (Parra y Hernández 2010).
Se seleccionaron 27 caracteres en total (Tabla 1), los cuales incluyen caracteres externos e internos de los adultos de machos y hembras, que se polarizaron con el grupo externo Tetracis y Catophoenissa. Los caracteres utilizados están basados en la revisión del género Syncirsodes publicado por Bocaz y Parra (2005).
Caracteres polarizados para el análisis filogenético de las especies Syncirsodes.
Para el análisis, se utilizó el software NONA 2.0 (Goloboff 1993) ejecutado por medio de Winclada (Nixon 2002), considerando todos los caracteres como desordenados (no aditivos). La matriz resultante (Tabla 2) se analizó mediante réplicas de secuencias de adición al azar, con permutaciones de ramas de tipo "tree bisection reconnection" (TBR) (Swofford y Olsen 1990) y evaluando posteriormente el apoyo de los grupos resultantes con "parsimony bootstrap" (Felsenstein 1985).
Matriz de datos de caracteres polarizados para el género Syncirsodes. Los números del 1 al 27 representan los 27 caracteres de la Tabla 1.
Resultados
El análisis cladístico encontró un único árbol más parsimonioso, resultante de la búsqueda exhaustiva (Fig. 4) con una longitud de 45, índice de consistencia 0,91 y un índice de retención de 0,77. Las especies analizadas forman un grupo monofilético (95% apoyo "bootstrap"), sustentado por siete sinapomorfias (Tabla 1); estas son: 2(1) presencia de un lóbulo entre la costa y vena M3 en el ala anterior de la hembra; 10(1) región media del gnathos densamente espinosa; 14(1) anellus armado con microespinas; 16(1) anellus esclerosado y con forma de tridente; 17(1) anellus unido a una membrana esclerosada; 21(1) signum ausente; 23(1) corpus bursae globoso.
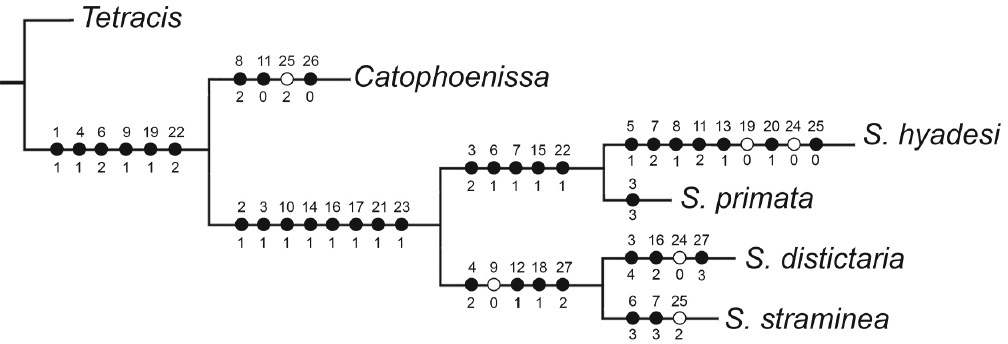
Hipótesis filogenética para el género Syncirsodes (L = 51; CI = 0.94; RI = 0.83). Círculos negros: sinapomorfias y autapomorfias; círculos blancos: caracteres multiestados que no representan sinapomorfías para el grupo y subgrupos. Números sobre los círculos corresponden al carácter y bajo los círculos al estado del carácter; los valores de bootstrap se ubican en cuadros en la base de los clados.
El análisis del grupo interno muestra una dicotomía que da origen a dos grupos. El primer grupo está formado por S. hyadesi y S. primata, sustentado por cuatro sinapomorfias (91% "bootstrap"): 6(1) forma del brazo de la furca subtriangular; 7(1) ángulo del cuerpo de la furca, 125° respecto del tallo de la furca; 12(1) presencia de un grupo de espinas largas en la zona posterior de la valva; 15(1) presencia de una proyección triangular esclerosada en el ápice del phallus; 22(1) ductus bursae subigual o un poco más largo que ancho. El segundo grupo está constituido por S. distictaria y S. straminea (82% "bootstrap"), condición sustentada por cuatro sinapomorfias: 4(2) apófisis de la furca 1/10 del largo del tallo de la furca; 12(1) ausencia de espinas en las valvas; 18(1) sin lamella postvaginalis; 27(2) saccus de forma rectangular. Ambos clados constituyen entonces grupos hermanos.
S. hyadesi es la especie que presenta el mayor número de autapomorfias, siete caracteres derivados de las armaduras genitales y del metafurcaesterno del macho y de la hembra: 4(2) apófisis del metafurcaesterno 1/7 del largo del tallo de la furca; 5(1) extremo apical de la apófisis de la furca romo; 7(2) ángulo del cuerpo de la furca respecto del tallo de la furca sobre 110°; 8(1) uncus espatulado; 11(2) forma de las valvas subrectangulares; 13(1) proceso en la costa de la valva; 20(1) lamella postvaginalis esclerosada; 25(0) longitud de las apófisis posteriores de menos de 1mm. En cambio, S. primata tiene solo una autapomorfía: 3(3) franja medial en las alas anteriores de las hembras oscura.
En el otro clado, S. distictaria presenta tres autapomorfías asociadas a las armaduras genitales del macho y de la hembra y del metafurcaesterno: 3(4) franja medial en las alas anteriores de las hembras bicolor; 16(2) anellus en forma de tridente no esclerosado; 27(3) saccus de forma triangular. Con respecto a S. straminea presenta dos autapomorfías: 6(3) membrana de la furca subromboidal; 7(3) ángulo de 100° de inclinación del cuerpo de la furca respecto del tallo de la furca; constituyen las autapomorfias que sustentan la especie.
Discusión
El análisis filogenético permite señalar que el género Syncirsodes constituye un grupo natural cuya monofilia está sustentada (95% apoyo "bootstrap") por siete sinapomorfias: seis corresponden a caracteres de la armadura genital de los imagos y una está relacionada a la forma de las alas en las hembras.
Pitkin (2002), propone las microespinas presentes en el anellus como el carácter diagnóstico del género; en este trabajo se reafirma esta proposición y se suman como caracteres relevantes la presencia de espinas dispuestas densamente en el gnathos, la ausencia de signum, el anellus esclerosado y con forma de tridente y la forma globosa del corpus bursae; evidencias de una historia evolutiva común.
En el análisis los caracteres de la armadura genital, tanto del macho como de la hembra, resultaron fundamentales en la delimitación del género, también como evidencia en los procesos de divergencia de los grupos de especies observados, como también en los caracteres diagnósticos de cada una de las especies. Sin embargo, el metafurcaesterno solamente entregó caracteres diagnósticos para distinguir especies, tales como la relación entre el largo de la apófisis (af) y el tallo de la furca (tf), así como también en la forma del brazo de la furca (fbf) y en el tamaño de ésta (Fig. 3). El aporte de estos caracteres ya había sido reportado para otros grupos de lepidópteros como los Palaephatidae (Davis 1986) para separar géneros y especies. La evidencia aportada por esta estructura en sus diversos caracteres y estados, en el presente estudio, es el indicio de que el metafurcaesterno puede entregar caracteres valiosos para distinguir especies, al menos en los Geometridae.
Así como los cambios en atributos particulares de las armaduras genitales tienen directa relación con la selección sexual (Arnqvist 1997), las variaciones y cambios en las estructuras del metafurcaesterno tendrían relevancia en el vuelo de los individuos (Angulo et al. 1986), esto debido a que su función estructural es servir como soporte para la inserción de los diversos músculos que se encuentran asociados a las alas (Ross 1964). Las variaciones en el metafurcaesterno entre los linajes, se relacionan de esta manera directamente con las distintas capacidades de vuelo en las especies, comportamiento que se asociaría a diferencias en la capacidad de dispersión, tanto para la búsqueda de recursos como de pareja en el período de reproducción (Van Dyck y Matthysen 1999). Seguramente, estos cambios morfológicos son los responsables de las diferencias en los patrones de comportamiento entre individuos de distintas poblaciones, permitiendo el aislamiento reproductivo y posterior especiación y diversificación de este género.
Footnotes
Agradecimientos
A los proyectos VRID 210.113.079-1.0 y 214.113.087-1.0, por el apoyo económico brindado en esta iniciativa.