
Abstract
The coffee berry borer, Hypothenemus hampei, is one of the most devastating coffee pests (Coffea arabica). Aspartic proteases were identified in protein extracts of adult insects, showing two bands of proteolytic activity in zymograms and a maximum activity at pH 2.5 in the spectrophotometric assays. Considering its inhibition by pepstatin A and hemoglobin hydrolysis this enzyme was classified as cathepsin D. Seeds extracts of Lupinus bogotensis, Brachiaria humidicola, Amaranthus hypochondriacus, Phaseolus acutifolius, Phaseolus coccineus, Hyptis suaveolens, Centrosema pubescens, and Trifolium repens were evaluated to identify aspartic protease inhibitors. The greatest inhibitory activity of aspartic proteases was found in L. bogotensis extract with a specific activity of 74.1 IU/mg, compared with extracts of H. suaveolens, B. humidicola, and A. hypochondriacus that showed a lower inhibition of aspartic proteases. Other protein extracts did not inhibit aspartic protease activity. The proteolytic activity of H. hampei was inhibited by 90% with 100 µg of crude extract of L. bogotensis, whereas it took 1 mg of crude extract of H. suaveolens and B. humidicola to inhibit the activity at 70 and 60% respectively. The zymograms identified potential protease inhibitors in extracts of L. bogotensis, H. suaveolens, B. humidicola and A. hypochondriacus that blocked in vitro activity of aspartic proteases of the insect. The expression of protease inhibitor genes in coffee is an alternative to obtain resistant varieties.
Introducción
El café (Coffea spp., Rubiaceae) es el producto agrícola más importante en 70 países de la región tropical húmeda (Jaramillo et al. 2006). En Colombia, el cultivo de café representa el 12,4% del producto interno bruto agropecuario (MADR-IICA-OAC 2008). La broca del café, Hypothenemus hampei (Ferrari, 1867) (Coleoptera: Curculionidae) es el insecto plaga que causa las mayores pérdidas económicas al café; debido a que se alimenta del endospermo disminuyendo el rendimiento y alterando la calidad del grano (Duque y Baker 2003; Bustillo 2008).
Muchos caficultores controlan H. hampei con insecticidas como endosulfán, clorpirifos, pirimifos metil y fenitrotion (Villalba et al. 1995). La creciente preocupación ambiental y el incremento de la resistencia de H. hampei a los insecticidas (Navarro et al. 2010), han estimulado la búsqueda de estrategias de control amigables con el ambiente (Jaramillo et al. 2006). Entre estas estrategias, diversas proteínas vegetales tienen potencial para el control de insectos incluyendo las lectinas, los inhibidores de proteasas (IPs), los inhibidores de a-amilasas y las proteínas de almacenamiento de las semillas, vicilinas, leguminas y arcelinas (Macedo et al. 2007; Prasad et al. 2010; Barbosa et al. 2010; Sales et al. 2005).
La habilidad de los IPs para interferir con el crecimiento y desarrollo de insectos se atribuye a su capacidad para impedir el acceso al substrato, reduciendo la disponibilidad de aminoácidos de las proteínas ingeridas Ryan (1990); y en algunos casos ocasionando la muerte, como el inhibidor de tripsina Kunitz de soya sobre el coleóptero Anthonomus grandis Boheman, 1843 (Franco et al. 2004). Los IPs se clasifican según la enzima digestiva con la que interactúan en serín, cisteín, aspártico, metalo, glutámico y treonín proteasa (Laskowski et al. 2003). Según, Molina et al. (2010) la digestión en el intestino medio de H. hampei es facilitada por aspártico proteasas activas al pH ácido de esa región corporal (Ossa et al. 2000). De acuerdo con esta información, la inhibición de las aspártico proteasas es un mecanismo de defensa promisorio para el control de este coleóptero.
Todas las plantas tienen algún nivel de resistencia endógena al ataque por insectos plaga. Sin embargo, como resultado de la co-evolución los insectos se adaptan a los mecanismos de defensa de las plantas por evasión o detoxificación (Gatehouse 2002). Por ejemplo, los coleópteros se alimentan de leguminosas que tienen un alto contenido de compuestos tóxicos, mientras que otros insectos no utilizan estas semillas como fuente de alimento (Sales et al. 2005). Además, los insectos comprometen las estrategias de defensa de las plantas usando sus mecanismos de señalización, es así como Helicoverpa zea (Boddie, 1850) emplea las moléculas de señalización, jasmonato y salicilato de las plantas huésped, para activar cuatro de sus genes citocromo P450, facilitando la inducción de sus enzimas de detoxificación de manera rápida y específica (Li et al. 2002). Esta asociación es altamente específica, por esto semillas de muy pocas especies sirven de alimento a un insecto particular (Gatehouse 2002).
La co-evolución de H. hampei y el género Coffea sugiere que es necesario evaluar el efecto de IPs de plantas nohuésped como leguminosas y solanáceas, en las cuales se han identificado IPs efectivos contra insectos (Sumikawa et al. 2010; Tamhane et al. 2007). Recientemente se identificó el primer inhibidor de aspártico proteasa de Lupinus bogotensis (LbAPI) que tiene una identidad de 76% con vicilinas de las semillas de Lupinus albus (Linnaeus, 1753). Esto sugiere que los IPs de aspártico proteasas son proteínas multifuncionales que actúan como proteínas de almacenamiento de las semillas, inhiben aspártico proteasas, y también pueden unirse a las estructuras que contienen quitina en el intestino medio de coleópteros como H. hampei (Molina et al. 2010). Sin embargo, para conseguir resistencia durable contra un insecto es necesario utilizar varios IPs, por esto el objetivo de este trabajo es la identificación de inhibidores de las aspártico proteasas en plantas no-huésped y sus variaciones en especificidad contra las proteasas digestivas de H. hampei.
Materiales y Métodos
Materiales
Se evaluaron semillas de cinco plantas de la familia Fabaceae: L. bogotensis (Benth, 1845), Centrosema pubescens (Benth, 1839), Trifolium repens (Linnaeus, 1753), Phaseolus acutifolius (Gray, 1852) y Phaseolus coccineus (Linnaeus, 1753) y de una planta de la familia Poaceae: Brachiaria humidicola; estas semillas se adquirieron en la empresa Semicol (Bogotá, Colombia). También se probaron semillas de una Amaranthaceae: Amaranthus hypochondriacus (Linnaeus, 1753), y una Lamiaceae: Hyptis suaveolens (L., Poit. 1806), suministradas por el Centro de Investigaciones y de Estudios Avanzados (Cinvestav), Irapuato, México.
Los insectos adultos de la broca del café proporcionados por la unidad de cría de Cenicafé (Chinchiná, Colombia), se criaron en café pergamino seco con una humedad de 45% y se mantuvieron bajo condiciones controladas a una temperatura de 27°C con una humedad relativa del 65-75%; los insectos empleados para la extracción estaban recién emergidos de las semillas de café.
Se utilizaron reactivos grado analítico, NaCl, HCl, cloroformo, metanol y ácido acético de Merck (Darmstadt, Germany). La hemoglobina bovina, albúmina sérica bovina (BSA), pepsina (EC 3.4.23.1), pepstatina A, tripsina (EC 3.4 21.4), N a-benzoyl-L-arginine ethyl ester (BAEE), fenilmetilsulfonilo fluoruro (PMSF), ácido etilendiaminotetraacético (EDTA), trans epoxisuccinilo-l-leucilo-amida (4) guanidino butano (E-64), Tris, dodecil sulfato de sodio (SDS), glicerol, casa comercial Sigma-Aldrich (St. Louis, MO, USA). La solución de Bradford, la solución de acrilamida-bis-acrilamida, persulfato de amonio de Bio-Rad (Hercules, CA, USA). La resina de exclusión Bio-Gel® P-6DG empacada en una columna Econo-Pac® 10DG de Bio-Rad (Hercules, CA, USA). Las membranas de ultrafiltración de 30 kDa (Amicon, Millipore) y el dispositivo de filtración centrifuga Centriprep®, membrana Ultracel YM-3, limite de peso molecular nominal 3 kDa (Amicon, Millipore) (Beverly, MA, USA). Las membranas de diálisis de 3,5 kDa (Spectra/Por®) (Broadwick Street, CA, USA).
Extracción de las aspártico proteasas de H. hampei (APH)
Para la extracción de las aspártico proteasas se maceraron 100 g de insectos adultos en 0,2 M de búfer succínico pH 6,0 en una relación 1:5 p/v a 4°C. El homogeneizado se centrifugó a 9.500 g por 15 min a 4°C. El sobrenadante se dializó tres veces cada seis horas contra 5 1 de H20 con 100 ml de búfer succínico 50 mM pH 6,0 utilizando una membrana de 3,5 kDa (Spectra/Por®), el dializado se liofilizó y se utilizó como fuente de aspártico proteasas.
Ensayos de actividad inhibitoria
La actividad proteolítica de las aspártico proteasas de H. hampei y pepsina se determinó evaluando la hidrólisis de hemoglobina a 280 nm en 0,2 M de búfer citrato, 0,1 M NaCl pH 2,5 (Lenney 1975). El extracto proteínico se incubó con 1,3 ml de hemoglobina al 1% como sustrato. Después de dos horas se adicionaron 2,5 ml de Cl3CO2H 0,34 N para detener la reacción. Una unidad de actividad enzimática se definió como la cantidad de enzima que cataliza un incremento en 0,01 unidades de absorbancia. El inhibidor se incubó con el extracto proteínico de H. hampei durante 15 min (Lenney 1975). Una unidad de inhibición se definió como la diferencia entre las actividades proteolíticas medidas en ausencia y presencia del inhibidor. Las variables medidas fueron: las unidades de inhibición por ml de extracto (UI ml-¹), las unidades de inhibición por gramo de tejido (UI g-¹), las unidades de inhibición totales (UI) y la actividad específica, unidades de inhibición por miligramo de tejido (UI mg-¹). Se evaluó la inhibición de las aspártico proteasas por E-64 (Oliveira et al. 2002); EDTA; PMSF (Schwertz y Takenaka 1955) y pepstatina A (Lenney 1975). Una concentración final de 1 µM se utilizó para todos los inhibidores con la excepción de pepstatina A, el cual se usó en una concentración final de 1 μΜ.
Electroforesis en gel de poliacrilamida
La electroforesis en gel de poliacrilamida dodecil sulfato de sodio (SDS-PAGE) se realizó según Schägger y von Jagow (1987). Las proteínas usadas como estándar de peso molecular fueron: miosina (210 kDa), ẞ-galactosidasa (118 kDa), BSA (82 kDa), anhidrasa carbónica (40 kDa), inhibidor tripsina de soya (31 kDa), lisozima (17 kDa) y aprotinina (6 kDa). Las proteínas se detectaron por tinción con 0,1% de azul de Coomassie brillante G-250 por 30 min y la destinción se realizó con ACOH:H2O (1:9, v/v) por 1 h.
Zimogramas
Para el zimograma de actividad proteolítica 20 mg de extracto proteínico del intestino medio de H. hampei se separaron en un gel nativo al 10%. El gel se incubó en una solución de hemoglobina al 1% en 0,2 M de búfer citrato pH 2,5 durante 1 h a 4°C. Después, se retiró el exceso de substrato y el gel se incubó durante 4 h a 30°C. Finalmente, el gel se tiñó con azul de Coomassie durante 30 min, y se destiñó con ACOH:H2O (1:9, v/v) por 1 h.
En el zimograma de inhibición se preincubaron 20 µg de extracto proteínico del intestino medio de H. hampei con 40 µg de los extractos de las semillas de L. bogotensis, A. hypochondriacus, B. humidicola e H. suaveolens durante 30 min a 30°C. Después, se siguió el mismo procedimiento descrito en el zimograma de actividad proteolítica.
Para el zimograma del inhibidor, los extractos crudos de las semillas se separaron por SDS-PAGE. El gel se incubó en una solución de 1% de hemoglobina bovina en 0,2 M de búfer citrato pH 2,5 durante 1 h a 4°C. Luego, el gel se lavó con H2O desionizada y se incubó en una solución con 10 mg/ml de extracto proteínico de H. hampei, durante 4 h a 30°C. Finalmente, el gel se tiñó con azul de Coomassie durante 30 min, y se destiñó con AcOH:H2O (1:9, v/v) por 1 h. Los inhibidores de aspártico proteasas se observaron como bandas azules sobre fondo claro, porque las aspártico proteasas digieren la hemoglobina presente en el gel, a excepción del lugar donde la banda (inhibidor) esta localizado.
Actividad de APH en función del pH
Para determinar la actividad de las aspártico proteasas en función del pH, el extracto crudo de adultos de H. hampei (1 mg/ml) se incubó 30 min a 37°C con diferentes búfer (100 mM): citrato de sodio (pH 2-4), acetato de sodio (pH 5), fosfato de sodio (pH 6-7), Tris-HCl (pH 7,5). Después, se ajustó el pH a 2,5 para evaluar la actividad inhibidora contra las aspártico proteasas de H. hampei según Lenney (1975). Los resultados representan el promedio de tres pruebas independientes.
Identificación de los inhibidores de aspártico proteasas (IAP) en las semillas
Para la identificación de los IAP se realizaron ensayos enzimáticos y zimogramas. 200 g de las semillas secas se molieron en un molino criogénico marca Restch, la harina se tamizó a través de dos mallas de 1,4 mm y 0,85 mm en un separador estándar. La fracción más fina (150 g) se desgrasó en agitación durante 30 min en una mezcla de CHCl3:MeOH (750 ml, 4:1, v/v); este proceso se repіtió tres veces. La extracción de proteínas a partir de harina seca (100 g) se realizó en H2O (500 ml) o 0,2 M de búfer succínico pH 4,5 por 6 h a 4°C. Con todas las semillas se utilizó una relación 1:5, p/v a excepción de T. repens con una relación de 1:10, p/v. La suspensión se clarificó por centrifugación a 7.500 g por 30 min a 4°C. El precipitado se descartó y la suspensión liofilizada de los extractos de las semillas se adicionaron a una columna de exclusión por tamaño Econo-Pac® 10DG, empacada con Bio-Gel P-6DG para eliminar pigmentos (Molina et al. 2010). La proteína del extracto proteínico se precipitó con 80% de (NH4)2SO4. El precipitado se disolvió en búfer Tris-HCl 20 mM pH 8,0, se dializó contra H2O desionizada en una membrana de 3,5 kDa (Spectra/Por®) y se liofilizó. Las proteínas liofilizadas se disolvieron en 10 ml de búfer Tris-HCl 20 mM pH 8,0 para fraccionarlas por ultrafiltración con una membrana de 30 kDa. Las fracciones eluídas de la membrana de 30 kDa, que mostraron actividad inhibidora contra las aspártico proteasas de H. hampei, se concentraron utilizando el dispositivo Centriprep®, límite de exclusión 3 kDa y se utilizaron como fuente de inhibidor. Se evaluó la actividad inhibidora de los extractos crudos de las semillas contra H. hampei siguiendo el método de Lenney (1975).
Determinación de proteína
La concentración de proteína se determinó por el ensayo de Bradford modificado por Zor y Selinger (1996) usando BSA como estándar.
Análisis estadístico
Para la determinación de las mejores condiciones de extracción de los IPs se utilizó un experimento multifactorial, los tratamientos evaluaron el empleo o no de desgrase y la extracción con agua o 0,2 M de búfer succínico pH 4,5, se utilizó un diseño experimental completamente al azar y el número de repeticiones 3. Para la identificación de la semilla que produce la mayor inhibición de las aspártico proteasas de H. hampei se realizó un experimento unifactorial, se utilizó un diseño experimental completamente al azar y el número de repeticiones 3. La unidad experimental fue un tubo conteniendo las aspártico proteasas de H. hampei, el inhibidor de aspártico proteasas, el substrato y ácido tricloroacético.
A los experimentos se les aplicó un análisis descriptivo: se estimaron el promedio y la varianza para las unidades de inhibición totales (UI). Se comprobó la homogeneidad de varianza y se realizó el análisis de varianza, bajo un diseño completamente al azar. La comparación de medias se realizó mediante prueba de Duncan al 5%. Los datos se procesaron en un computador IBM con el programa SAS versión 9.1.
Resultados
Zimograma de las APH
Los zimogramas mostraron dos bandas de actividad aspártico proteasa en los extractos de intestinos (Fig. 1A) y de adultos completos de H. hampei (Fig. 1B), resultados similares registró Molina et al. (2010). También, Preciado et al. (2000) encontraron tres bandas de actividad proteolítica en el intestino medio y en brocas adultas. El hallazgo de actividad aspártico proteasa en el intestino medio confirmó su función digestiva. De acuerdo con estos resultados y considerando que H. hampei es de 0,8 a 0,7 mm de longitud (Bergamin 1943), se utilizaron extractos proteínicos de adultos completos, para obtener la cantidad suficiente de enzima para los ensayos de actividad proteolítica y de inhibición.
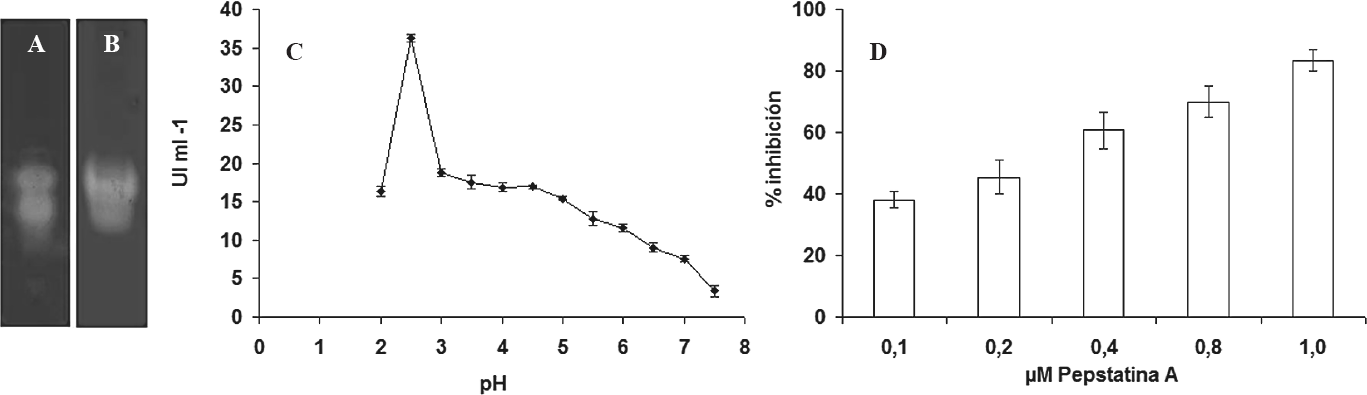
A. Zimograma del extracto crudo de intestino de H. hampei usando un perfil electroforético nativo al 10%. B. Zimograma de extracto crudo de insectos adultos de H. hampei usando un perfil nativo al 10%. C. UI ml-¹ de las APH en función del pH. D. Porcentaje de inhibición de la actividad APH en función de la concentración de pepstatina A (μΜ).
Actividad de las APH en función del pH
Las APH son activas a pH entre 2 a 5, con una actividad máxima a pH 2,5 (Fig. 1C). Una característica particular de este tipo de enzimas es su rango óptimo de actividad a pH entre 2 - 6 (Silva y Xavier-Filho 1991; Blanco et al. 1996; Wilhite et al. 2000; Liu et al. 2004; Valencia y Arboleda 2005), el cual coincide con el pH ácido del intestino medio de H. hampei (Ossa et al. 2000). La actividad de las APH se evaluó usando como sustrato hemoglobina y BSA, encontrando que las aspártico proteasas digirieron 100% de hemoglobina y 10% de BSA. Esto demostró que estas enzimas digestivas son catepsina D, como previamente registraron Valencia y Arboleda (2005).
Inhibición de la actividad de las APH por IPs
Pepstatina A en una concentración de 1 µM inhibió la actividad proteolítica de H. hampei en 83,3% (Fig. 1D), al igual que lo registrado por Valencia y Arboleda (2005). Mientras que los inhibidores de serín proteinasa: PMSF (1mM), cisteín proteinasa: E-64 (1mM) y metaloproteinasa: EDTA (1mM), no inhibieron las enzimas digestivas del insecto. Esto confirmó la presencia de aspártico proteasas en el intestino medio.
Extracción de los IAP en las semillas de L. bogotensis y B. humidicola
El extracto acuoso de L. bogotensis con harina desgrasada presentó 8.764 UI y una actividad específica de 97,4 UI mg¯¹, siendo esta inhibición significativamente superior al resto de los tratamientos, según la prueba de Duncan al 5%. Del mismo modo, el extracto acuoso de B. humidicola con harina desgrasada mostró la mayor inhibición de las APH con 2.426 UI son miles no decimas (Tabla 1). El empleo de harina desgrasada en la extracción con agua o con búfer succínico 0,2 M pH 4,5, aumentó significativamente la inhibición (P<0,0001) de las APH (4.284 UI) en comparación con la harina sin desgrasar (2.572 UI). Esto indica que la grasa interfiere con la determinación de la actividad inhibidora de las APH. La extracción de los IPs fue mejor con agua, obteniéndose una inhibición mayor de las proteasas de H. hampei (4.215 UI) que con búfer succínico 0,2 M pH 4,5 (2.641 UI). Con base en estos resultados se seleccionó la extracción de los inhibidores de aspártico proteasas de harina desgrasada y agua en una relación 1:5, p/v.
Actividad inhibidora de los IAPs de las semillas de L. bogotensis y B. humidicola contra las APH.
= p < 0.0001.
Identificación de IAP en los extractos vegetales
100 µg del extracto conteniendo los inhibidores de aspártico proteasas de L. bogotensis (IAPLb) disminuyeron en 100% y 90% la actividad de pepsina y de las APH, respectivamente (Fig. 2A); mientras que con B. humidicola e H. suaveolens se requiere 1 mg de extracto crudo para inhibir la actividad de las APH en 60% y 70%, respectivamente (Fig. 2B). Estos resultados son promisorios ya que se inhibió la actividad de las APH con una concentración muy baja de IAPLb.
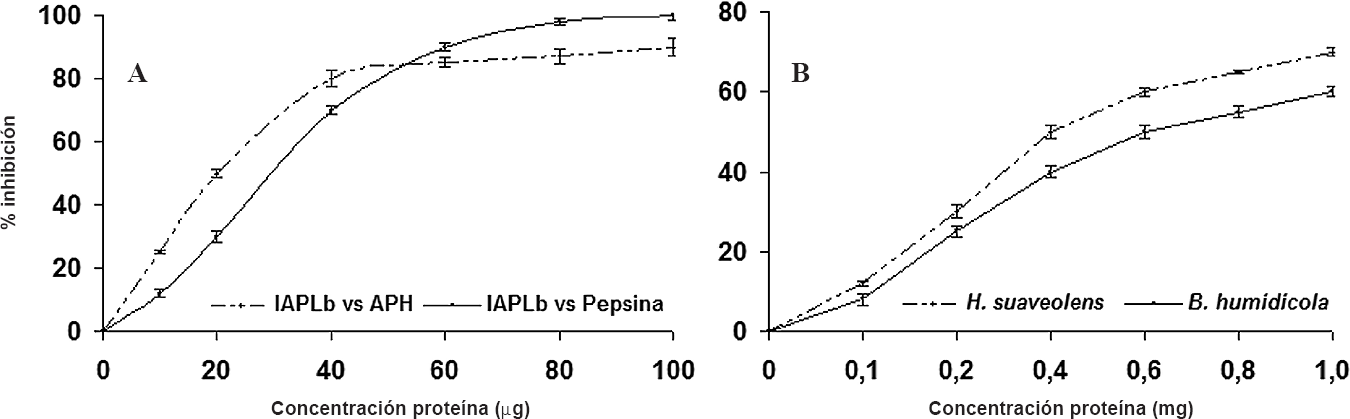
A. Porcentaje de inhibición de las APH y pepsina en función de la concentración de proteína (ug) de los IAPLb. B. Porcentaje de inhibición de las APH en función de la concentración de proteína (mg) del extracto de B. humidicola e H. suaveolens.
El extracto de L. bogotensis produjo la mayor inhibición de las proteasas de H. hampei con 5.560 UI y una actividad específica de 74,1 UI mg¯¹, en comparación con H. suaveolens, B. humidicola y A. hypochondriacus que fueron iguales, según la prueba de Duncan al 5% (Tabla 2). Los extractos proteínicos de P. coccineus, P. acutifolius, T. repens y C. pubescens no inhibieron las aspártico proteasas de H. hampei. Las diferencias en la inhibición podrían deberse a la selectividad que los inhibidores de plantas han desarrollado para reconocer enzimas específicas de insectos, a la inactivación de los IPs por proteasas digestivas y a variaciones en afinidad entre los IPs y las enzimas digestivas (Ryan 1990).
Actividad inhibidora de los IAPs de las semillas de L. bogotensis, B. humidicola, A. hypochondriacus e H. suaveolens contra las APH.
Promedios seguidos por la misma letra son iguales (P<0,0001).
Zimogramas para la identificación de IAP
los zimogramas son determinaciones cualitativas, que se utilizaron para identificar las bandas de IPs en los extractos proteínicos, mientras que los ensayos de inhibición cuantifican la inhibición de las aspártico proteasas de H. hampei por los IPs de extractos de plantas. Valencia et al. (2007) usaron este método para la detección de inhibidores de a-amilasas de Phaseolus vulgaris (Linnaeus, 1753). La separación electroforética de los extractos proteínicos de las semillas mostró la presencia de bandas en un rango de masa molecular entre 6 a 30 kDa (Fig. 3A). El zimograma mostró que las bandas de proteína de B. humidicola, H. suaveolens, L. bogotensis y A. hypochondriacus son resistentes a la digestión por las APH del insecto; esto sugiere que son inhibidores tipo aspártico proteasa (Fig. 3B). Mientras que las proteínas de los extractos de C. pubescens, T. repens, P. acutifolius y P. coccineus, son digeridas por las aspártico proteasas de H. hampei y no actúan como IPs; confirmando los resultados de los ensayos de inhibición.
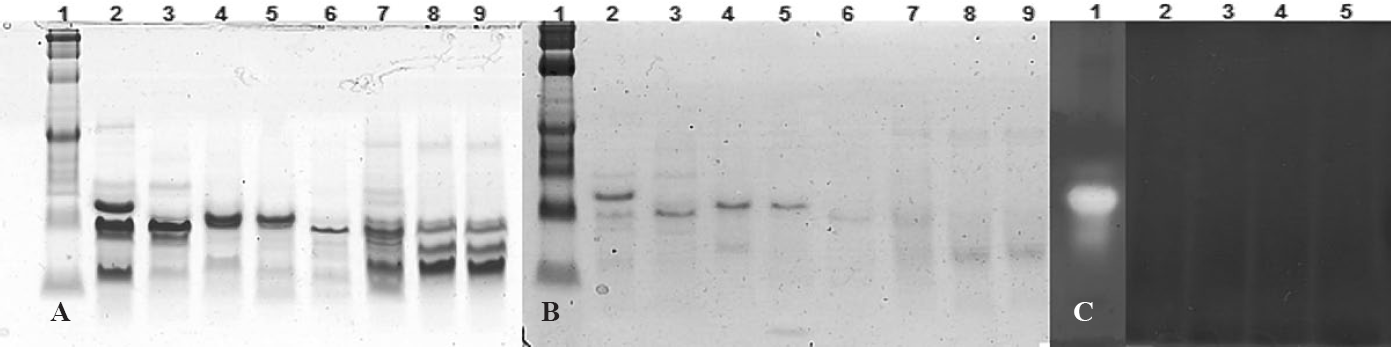
A. Separación electroforética de los extractos proteínicos de las semillas de plantas. B. Zimograma de los extractos proteínicos de las semillas de plantas, en condiciones denaturantes al 16%. 1. Marcador de peso molecular. 2. B. humidicola. 3. H. suaveolens. 4. L. bogotensis. 5. A. hypochondriacus. 6. C. pubescens. 7. T. repens. 8. P. acutifolius. 9. P. coccineus. C. Zimograma de inhibición de las APH por inhibidores de proteasas de los extractos proteínicos, usando electroforesis nativa al 10%. 1. Control, extracto de adultos de H. hampei. 2. L. bogotensis. 3. B. humidicola. 4. H. suaveolens. 5. A. hypochondriacus.
Zimograma de inhibición
En este zimograma (Fig. 3C) no se observaron las bandas de actividad proteolítica cuando se incubó el extracto de H. hampei con los inhibidores presentes en los extractos de L. bogotensis, B. humidicola, H. suaveolens y A. hypochondriacus, confirmando los resultados de los ensayos de inhibición. Esto indica que los IPs se unen al sitio activo de las aspártico proteasas inhibiendo su actividad, lo cual se refleja en la desaparición de las dos bandas de actividad proteolítica.
Discusión
Para la identificación de inhibidores de las APH se utilizaron extractos proteínicos de insectos adultos que representan la actividad aspártico proteasa. De forma similar, extractos de las larvas del coleóptero Tribolium castaneum (Herbst, 1797) se emplearon para purificar la aspártico proteinasa tipo catepsina D, debido a que los zimogramas mostraron actividad proteolítica en intestinos y larvas, demostrando la naturaleza digestiva de esta enzima (Blanco et al. 1996). En otra investigación se encontró que en Prostephanus truncatus (Horn, 1878) la actividad a-amilasa es mayor en larvas de segundo instar y en adultos (Mendiola et al. 2000). Además, la actividad de a-amilasa y de tripsina de P. truncatus fue más alta en adultos que en larvas; esto sugiere que es mejor utilizar adultos para evaluar el efecto de inhibidores de estas enzimas (Vásquez et al. 1999).
Las aspártico proteasas de H. hampei presentaron un rango óptimo de actividad a pH 2 a 5. De manera similar, otros coleópteros como Callosobruchus maculatus (Fabricius, 1775), T. castaneum, P. truncatus, Hypera postica (Gyllenhal, 1813), Zabrotes subfasciatus (Boheman, 1833), Leptinotarsa decemlineata (Say, 1824) y Diabrotica virgifera (LeConte, 1868), que utilizan aspártico proteasas para la digestión de las proteínas del alimento, tienen una actividad proteolítica óptima en un rango de pH de 3 a 6 (Silva y Xavier-Filho 1991; Blanco et al. 1996; Vásquez et al. 1999; Wilhite et al. 2000; Lemos et al. 1990; Wolfson y Murdock 1987; Liu et al. 2004). En general, las aspártico proteasas se han encontrado en insectos de los ordenes hemiptera y coleóptero que se caracterizan por el pH ácido del intestino medio (Houseman y Downe 1983).
La presencia de aspártico proteasas en el intestino medio de H. hampei se confirmó por su inhibición con pepstatina A, este inhibidor ha sido utilizado para identificar aspártico proteasas en el intestino medio de varios coleópteros como T. castaneum, C. maculatus, Z. subfasciatus e H. hampei, por la pérdida de la actividad proteolítica de la enzima (Blanco et al. 1996; Silva y Xavier-Filho 1991; Valencia y Arboleda 2005). La actividad aspártico proteasa del intestino medio de las larvas de Z. subfasciatus fue inhibida completamente con el empleo de 14 µM de pepstatina A (Lemos et al. 1990). Del mismo modo, la actividad proteolítica del intestino medio de H. postica se redujo en 50% por la adición de pepstatina A y del inhibidor de catepsina D de papa (Mares et al. 1989). Mientras que el inhibidor de aspártico proteasas de calabaza (SQAPI), que inhibe pepsina pero no catepsina D (Christeller et al. 1998), no inhibió la actividad proteolítica de H. postica, esto confirmó que catepsina D es la principal aspártico proteinasa de este gorgojo (Wilhite et al. 2000).
Se concluye que las aspártico proteasas de H. hampei son de tipo catepsina D porque digieren casi exclusivamente hemoglobina. De forma similar, la enzima aislada de T. castaneum se clasificó como catepsina D porque digiere únicamente hemoglobina (Blanco et al. 1996). También, la aspártico proteinasa presente en el intestino medio de las larvas de C. maculatus es catepsina D porque digiere 100% de hemoglobina y 30% de BSA (Silva y Xavier-Filho 1991). Catepsina D es una de las aspártico proteasas mejor estudiadas en vertebrados y artrópodos; esta enzima es responsable de la digestión intracelular y extracelular de proteínas en el lumen del intestino medio de muchos coleópteros donde las cisteín proteasas son las enzimas digestivas más importantes (Ahn y Zhu-Salzman 2009). En Aedes aegypti (Linnaeus, 1762) está involucrada en la producción de vitelogenina (Cho y Raickel 1992). En Ceratitis capitata (Wiedemann, 1824) su actividad coincide con la hidrólisis del cuerpo graso (Rabossi et al. 2004). En Bombix mori (Linnaeus, 1758) su acción en la muerte celular programada es clave en la metamorfosis para la transformación de pupa a larva (Lee et al. 2009). Esta enzima también participa en la digestión de la sangre en ácaros y garrapatas (Boldbaatar et al. 2006).
En C. maculatus y Z. subsfasciatus las aspártico y cisteín proteasas son responsables de la digestión de las proteínas del alimento (Silva y Xavier-Filho 1991). Es probable que en H. hampei ambas enzimas son responsables de la proteólisis como ocurre en otros coleópteros, debido a que las cisteín proteasas son activas en un rango de pH entre 5 a 7, cercano al de H. hampei. El conocimiento del papel de los diferentes tipos de proteasas en el tracto digestivo del insecto, ayudará a entender los sitios de vulnerabilidad bioquímica, por esto es necesario continuar con la caracterización de las enzimas digestivas de la broca del café.
Las mejores condiciones para la extracción de los inhibidores de las aspártico proteasas de semillas de plantas fueron el empleo de agua y de harina desgrasada. La extracción con agua ha sido empleada para aislar inhibidores de Cicer arietinum (Linnaeus, 1753), Gossypium arboreum (Linnaeus, 1753), Arachis hypogea (Linnaeus, 1753), Psophocarpus tetragonolobus (L., DC. 1825), Solanum tuberosum (Linnaeus, 1753), e H. suaveolens (Harsulkar et al. 1999; Giri et al. 2003; Aguirre et al. 2004). El procedimiento de desgrase se ha utilizado con éxito para la identificación y purificación de IPs en las semillas de A. hypochondriacus, Inga laurina (Sw., Willd. 1806), Cicer echinospermum (Davis, 1869), A. hypogea, y P. tetragonolobus (Valdes et al. 1993; Rodríguez et al. 2007; Harsulkar et al. 1999).
Se identificaron inhibidores de las aspártico proteasas de H. hampei en las semillas de L. bogotensis, B. humidicola, H. suaveolens y A. hypochondriacus, estos IPs bloquean la digestión de las proteínas del alimento en el intestino medio y podrían utilizarse para el control del insecto. La broca del café es monófaga, sugiriendo que su mecanismo de defensa podría ser menos complejo y más vulnerable a la acción de IPs de plantas no-huésped que los insectos polífagos. Sin embargo, los estudios con IPs han mostrado que cuando se inhiben las proteasas más importantes para la digestión de las proteínas del alimento en el intestino medio, se incrementa la actividad de otras enzimas insensibles al inhibidor y que en su ausencia no están sobre-expresadas. Esto permite la digestión de las proteínas de la dieta en presencia del inhibidor y es fundamental para la supervivencia de los insectos (Zhu-Salzman et al. 2003; Amirhusin et al. 2007). En consecuencia, para una inhibición efectiva de las enzimas digestivas de los insectos es necesaria la expresión de diversos IPs (Jongsma y Bolter 1997).
Para conseguir resistencia durable contra H. hampei, los IPs de las semillas de L. bogotensis, H. suaveolens y B. humidicola pueden transferirse a café mediante ingeniería genética para producir plantas trangénicas con variaciones en la resistencia contra este insecto. Plantas de tabaco que expresan genes del inhibidor de proteinasa de tomate I (fuerte inhibidor de quimotripsina y débil de tripsina) y II (fuerte inhibidor de tripsina y quimotripsina) mostraron diferencias en la especificidad contra Manduca sexta Linnaeus, 1763 (Johnson et al. 1989). El crecimiento de las larvas de M. sexta que se alimentaron de las hojas de tabaco transformadas con el inhibidor de proteinasa II se redujo significativamente en comparación con el crecimiento de las larvas que se alimentaron de las hojas sin transformar. Mientras, que el inhibidor de proteinasa I tuvo poco efecto en el crecimiento de las larvas de este lepidóptero (Johnson et al. 1989). Estos resultados muestran la importancia de identificar nuevos IPs con variaciones en la especificidad contra diferentes enzimas de los insectos (Mendiola et al. 2000).
H. hampei y C. arabica co-evolucionaron favoreciendo la adaptación del insecto a los compuestos tóxicos de la planta. Los IPs de L. bogotensis, H. suaveolens y B. humidicola, plantas no-huésped de la broca del café, inhiben las proteasas del insecto; mientras que los inhibidores de la planta huésped no bloquean las proteasas del insecto. Otros estudios han mostrado que los IPs de A. hypogaea, P. tetragonolobus y S. tuberosum, plantas no-huésped de Helicoverpa armigera (Hübner, 1805) inhiben las isoproteinasas y el crecimiento de las larvas de este lepidóptero, en comparación con los IPs de las plantas huésped (Harsulkar et al. 1999; Giri et al. 2003). De igual manera, el inhibidor de tripsina de Momordica charantia (Linnaeus, 1753) planta no-huésped de H. armigera y Spodoptera litura (Fabricius, 1775), afectó la fertilidad y fecundidad de las larvas de ambos insectos (Telang et al. 2003). También, las vicilinas de leguminosas no-huésped de C. maculatus como P. vulgaris, Phaseolus lunatus (Linnaeus, 1753), Canavalia ensiformis (L., DC. 1825), y Glicine max (L., Merr. 1917) retardaron el desarrollo de las larvas del insecto (Yunes et al. 1998). Mientras que las vicilinas de Vigna angularis (Willd, 1969) una especie genéticamente cercana a la planta huésped Vigna Unguiculata (L., Walp. 1843) no afectaron el desarrollo de las larvas de este insecto (Yunes et al. 1998). Estos resultados muestran la importancia de evaluar IPs de plantas no-huésped contra las enzimas de insectos en la búsqueda del control de plagas vía transgénesis.
Los inhibidores presentes en el extracto crudo de L. bogotensis produjeron la mayor inhibición de las aspártico proteasas de H. hampei usando una concentración de extracto proteínico de 100 µg, menor en comparación con otras investigaciones que usaron 160 µg de extracto crudo de M. charantia con una inhibición del 84% de la actividad de las proteinasas del intestino de H. armigera (Telang et al. 2003). Del mismo modo, una concentración diez veces mayor de extracto crudo de P. vulgaris y Amaranthus spp., sólo inhibió la actividad a-amilasa de H. hampei en un 80% y 40%, respectivamente (Valencia et al. 2000). De forma similar, 1 mg de extracto crudo de Amaranthus cruentus (Linnaeus, 1759) sólo inhibió 80% de la actividad amilasa de H. hampei (Valencia et al. 2000).
Estos resultados sugieren que los IAPLbs podrían ser buenos candidatos para transferir a café mediante ingeniería genética para producir plantas resistentes contra H. hampei, debido a que inhiben las enzimas del insecto a bajas concentraciones y al pH encontrado en el intestino medio del insecto. Además, esta leguminosa no ha estado expuesta a H. hampei, proporcionando nuevos y más potentes inhibidores de las proteasas de este insecto. Esto coincide con investigaciones previas que muestran que los IPs de plantas no-huésped son mejores para inhibir las proteasas de un insecto particular que los inhibidores de las plantas huésped (Harsulkar et al. 1999; Giri et al. 2003).
Conclusiones
Se identificaron inhibidores de las aspártico proteasas de H. hampei en diferentes semillas; los inhibidores de L. bogotensis produjeron una mayor inhibición de las aspártico proteasas del insecto en comparación con los IPs de H. suaveolens, B. humidicola y A. hypochondriacus. Los IPs se unen al centro activo de la enzima bloqueando su actividad proteolítica. Esto muestra que los IPs de plantas no-huésped pueden suministrar nuevos y más potentes inhibidores de las proteasas de H. hampei. Estos inhibidores de proteasas pueden transferirse a café en la búsqueda de resistencia durable contra H. hampei. Este trabajo junto con el de Scarafoni et al. (2008) y Molina et al. (2010) confirma la existencia de inhibidores de proteasas en las semillas de Lupinus spp., contrario a lo que se había publicado de la falta de actividad inhibitoria de las proteasas de L. albus y Lupinus luteus (Linnaeus, 1753) (Gallardo et al. 1974).
Footnotes
Agradecimientos
Agradecemos al Ministerio de Agricultura y Desarrolo Rural de Colombia por la donación de recursos para llevar a cabo este proyecto, a Colciencias por la beca de doctorado de Diana Molina, y a los revisores del manuscrito.