
Abstract
Disturbance increases the probability of dominance by the little fire ant Wasmannia auropunctata (Formicidae: Myrmicinae), an indicator species of low diversity ant communities in Tropical Dry Forest fragments of the Cauca River Valley (Colombia). In order to study interactions of interspecific competition involving this species, we conducted experiments with soil-surface tuna-baits at four localities: Alejandría, El Medio, El Vínculo and San Julián. Three biotopes were distinguished at each locality: forest interior, forest margin and their surrounding matrices (sugar cane or pasture). Based on information obtained from the composition of ants at the baits, competition ability indices were calculated. Attraction to the baits was 91%, for a total of 66 species in 30 genera. Species composition was mainly influenced by locality conditions. The myrmicines Pheidole, Solenopsis and Crematogaster had the greatest interaction with W. auropunctata and partitioning of the baits was mostly influenced by biotope. Bait use by other species was determined by the abundance of W. auropunctata; if the bait had more than a mean of 500 workers, the resource was not shared. W. auropunctata emerged as the dominant species in the upper Cauca River watershed and this was mainly accomplished by exploitation competition, allowing the ants to rapidly take advantage of resources by recruiting a large number of workers.
Introducción
La competencia es uno de los temas primordiales en estudios de ecología de comunidades de hormigas (Hölldobler y Wilson 1990; Davidson 1998; Holway y Case 2001). La coexistencia entre especies implica varias estrategias competitivas, descritas según las características de cómo las diferentes especies luchan por el recurso. Estas estrategias son: oportunista, insinuadora y extirpadora (Hölldobler y Wilson 1990; Wilson 01971). Las especies oportunistas practican la competencia por explotación y son aquellas que descubren rápidamente el recurso alimenticio y lo utilizan eficientemente. Las especies extirpadoras exhiben competencia por interferencia al dominar el recurso mediante agresión; mientras que las insinuadoras, son especies discretas que pueden aprovechar el recurso al ser inconspicuas. Algunas hormigas pertenecientes al grupo de las extirpadoras reclutan por pistas de olor y pelean por los recursos como por ejemplo las especies de los géneros Pheidole, Crematogaster y Camponotus, la hormiga de fuego Solenopsis geminata (Fabricius, 1804) y la pequeña hormiga de fuego Wasmannia auropunctata (Roger, 1863) (Hölldobler y Wilson 1990).
Las hormigas vagabundas como W. auropunctata pueden comportarse como invasoras en sitios donde han sido introducidas (Passera 1994). Debido al desplazamiento de especies nativas o a la reducción de su abundancia, los invasores biológicos han perturbado severamente la estructura y función de algunas comunidades y ecosistemas nativos (Mack et al. 2000). Las hormigas invasoras muestran características típicas como la poliginia, unicolonialidad, obreras monomórficas pequeñas y una fuerte agresividad interespecífica (Passera 1994), permitiéndoles excluir a las especies nativas (Passera 1994; Hölldobler y Wilson 1990). Algunas introducciones han tenido impactos negativos en la fauna de hormigas así como en otros invertebrados y vertebrados (Clark et al. 1982; Walsh et al. 2004; Porter y Savignano 1990; Morrison 2002). Este fenómeno es liderado por W. auropunctata (Walsh et al. 2004), Solenopsis invicta (Buren, 1972) (Forys et al. 2002) y Linepithema humile (Mayr, 1868) (Suárez et al. 1998).
En su rango nativo W. auropunctata puede coexistir con otras especies de hormigas (Tennant 1994) o dominar en áreas fragmentadas (Brandão y Silva 2006), invadir campos cultivados (Posada et al. 1976; Ulloa-Chacón y Cherix 1990; Delabie et al. 1994) y ecosistemas muy intervenidos como el Bosque seco Tropical (Bs-T) del valle geográfico del río Cauca (Armbrecht y Ulloa-Chacón 2003). Es por esto que se hipotetiza que la perturbación desempeña un papel importante al incrementar la posibilidad de invasión por esta especie, la cual ha sido propuesta como indicadora de baja diversidad de la comunidad de hormigas en Bs–T (Armbrecht y Ulloa-Chacón 2003). En cuanto a su capacidad competitiva, experimentos con diferentes cebos indicaron que W. auropunctata es muy exitosa al remplazar a otras especies de hormigas en los cebos (Clark et al. 1982; Meier 1994; Tennant 1994; Le Breton et al. 2005).
Con base en lo anterior, este trabajo busca estimar la composición de hormigas atraídas a cebos de atún en cuatro sitios que conservan fragmentos de Bosque seco Tropical y en sus respectivas matrices (potreros y cultivos de caña de azúcar) para establecer si el sitio o los biotopos (bosque, matriz) son los factores que determinan la composición de especies. También se espera identificar las principales hormigas competidoras de la pequeña hormiga de fuego, y determinar si la abundancia de esta especie se relaciona con la riqueza de otras especies de hormigas en el recurso alimenticio.
Materiales y Métodos
Área de trabajo
El estudio se llevó a cabo en cuatro sitios localizados a lo largo de la cuenca alta del valle geográfico del río Cauca en jurisdicción de los departamentos de Cauca, Valle del Cauca y Risaralda. La temperatura promedio anual es de 24°C y la precipitación fluctúa entre 1.000 y 2.000 mm (Álvarez et al. 1998). En la tabla 1 se describen las principales características de cada sitio, incluyendo el fragmento de Bosque seco Tropical y su respectiva matriz.
Muestreo
Se realizó durante tres meses de época lluviosa (junio de 2004, marzo y abril de 2005), cuando las hormigas son más abundantes (Chacón de Ulloa 2003). En cada sitio se diferenciaron tres biotopos: interior de bosque, borde de bosque y matriz (caña o potrero). Se tomó como borde la franja perimetral de 50 m a partir del límite del bosque hacia su interior; excepto en el fragmento de San Julián donde no se pudieron diferenciar los biotopos del bosque por su forma alargada y reducida extensión. En cada biotopo se delimitó un transecto lineal donde se marcaron en promedio 33 estaciones equidistantes (20 m), en las cuales se capturaron hormigas mediante cebos epígeos de atún. Este método ha sido ampliamente usado en bosque seco (Armbrecht y Chacón de Ulloa 1997a; Armbrecht y Ulloa-Chacón 1999; Ramírez 2001; Armbrecht y Ulloa-Chacón 2003), y consiste en una pieza de papel bond blanco de 4 x 4 cm donde se ponen 9 g (± 1,7) de atún en aceite, luego se coloca sobre la superficie del suelo y se deja expuesto por tres horas. Seguidamente, se recolectan las hormigas que se encuentren directamente sobre el atún y se recoge el resto del cebo, incluyendo parte de suelo, ya que en la superficie inferior del papel se pueden encontrar especies muy pequeñas o menos agresivas (Bestelmeyer et al. 2000). El material se introdujo en bolsas de cierre hermético (16,5 cm x 14,9 cm) y se mantuvo refrigerado. En el laboratorio, las muestras se limpiaron y conservaron en etanol al 80%. La identificación a nivel de género se realizó siguiendo a Palacio y Fernández (2003) y el 53% de las hormigas se llevaron a especie utilizando la clave de Longino (2003), y por comparación con las colecciones del laboratorio de mirmecología del Centro de Pesquisas do Cacau CEPEC (Itabuba BA, Brasil) y del Museo de Entomología de la Universidad del Valle MEUV (Cali, Colombia), donde se depositaron los especímenes.
Análisis de datos
Para cada muestra se contabilizó el total de individuos de cada morfoespecie. Para caracterizar la composición de hormigas atraídas al atún, se describió el porcentaje de ocupación de cebos por las diferentes subfamilias y géneros. Con los datos de frecuencia de captura de todas las especies, se exploró la similitud entre biotopos y sitios de muestreo mediante análisis de agrupamiento (McCune y Grace 2002). Posteriormente, se utilizó el programa PC-ORD versión 4 (McCune y Mefford 1999) para comparar los grupos formados usando la técnica multivariada no paramétrica MRPP (Multi Response Permutation Procedure) la cual pone a prueba la hipótesis de no diferencia entre dos o más grupos de entidades y requiere grupos previamente definidos (Zimmerman et al. 1985; McCune y Mefford 1999). Esta misma técnica se realizó para comparar los ensamblajes de hormigas entre biotopos y entre sitios.
Ubicación geográfica y características de los cuatro fragmentos de Bosque seco Tropical estudiados.
Para conocer el porcentaje de interacción, se examinó el número de cebos compartidos entre W. auropunctata y otras especies de hormigas. Además, se realizaron pruebas de Chi-cuadrado (Zar 1996) para determinar si la partición del cebo es independiente del biotopo. Para establecer si el número de especies de hormigas que se encuentra en los cebos se relaciona con la abundancia de W. auropunctata, se aplicó una prueba de Kruskall-Wallis (H) usando el programa STATISTICA 6.1 (Statsoft 1998). Para las cinco especies de hormigas más abundantes, se calcularon dos índices de habilidad competitiva: Explotación (a), razón entre el número de grupos de forrajeo grandes (> de 20 obreras por cebo) y el número total de ocurrencias; e Interferencia (b), razón entre el número de ocurrencias de obreras de una sola especie y el número total de ocurrencias (Von Aesch y Cherix 2005).
Resultados
Composición de hormigas en cebos
De los 397 cebos colocados el 90,9% resultaron positivos para hormigas (Tabla 2) y la atracción promedio fue del 90,3% (± 9,06). El número total de hormigas contabilizadas fue de 79.391 agrupadas en 30 géneros y 66 morfoespecies (Apéndice). Se registraron ocho subfamilias con diferentes grados de atracción: Myrmicinae (80%), Formicinae (9,9%), Dolichoderinae (5,1%), Ponerinae (2,0%), Pseudomyrmecinae (1,3%), Ectatomminae (1,0%), Ecitoninae (0,7%) y Heteroponerinae (0,2%). Cinco géneros que se encontraron en todos los sitios, presentaron la mayor frecuencia de ocupación: Wasmannia (42,1%), Pheidole (30,7%), Solenopsis (23%), Crematogaster (15%) y Camponotus (6,3%).
Ensamblajes de hormigas
El dendograma resultante (Fig. 1) presentó un encadenamiento del 15,15% y separó cuatro grupos, con un nivel de retención de información por encima del 60%. El primer grupo correspondió a los tres biotopos de Alejandría. El segundo, lo formaron los biotopos boscosos (interior y borde) de El Medio y El Vínculo. El tercer grupo abarcó a San Julián y a la matriz de El Vínculo; y el cuarto grupo, solamente agrupó dos matrices. Hubo diferencias significativas (MRPP, A = 0,226; P < 0,01) cuando se compararon todos los grupos formados mediante el análisis de agrupamiento, es decir que la composición de especies de hormigas observada difiere de la que se esperaría si los grupos se formaran al azar. Seguidamente, al comparar los grupos definidos solamente por los sitios (Alejandría, El Medio, San Julián y El Vínculo), también se encontraron diferencias significativas (MRPP, A = 0,144; P < 0,01); pero no ocurrió lo mismo cuando se compararon los grupos con base en los biotopos (interior bosque, borde bosque, potrero y caña) (MRPP, A = 0,019; P = 0,31).
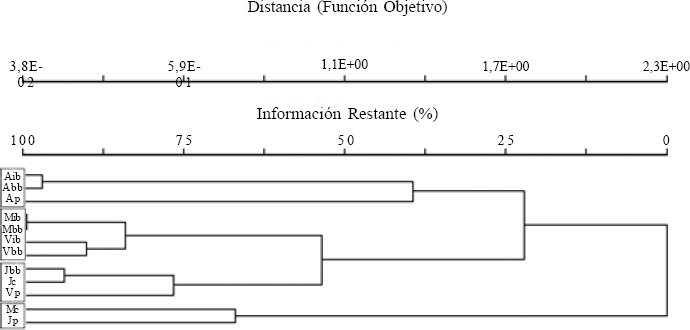
Agrupamiento de los ensambles de hormigas de los cuatro sitios y sus respectivos biotopos, con base en el índice de disimilitud de Sorensen (Bray-Curtis) y el método de agrupamiento Beta-flexible (β = -0.25). Las letras mayúsculas se refieren a los sitios A: Alejandría, M: El Medio, V: El Vínculo y J: San Julián. Las letras minúsculas denominan los biotopos ib: interior de bosque, bb: borde de bosque, p: potrero y c: caña de azúcar.
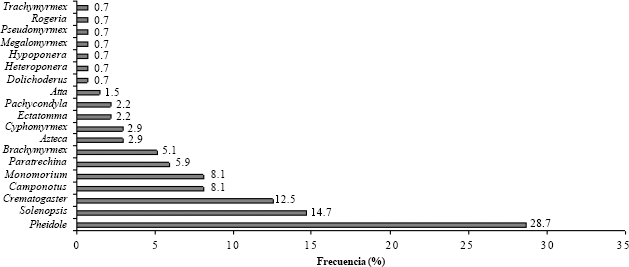
Géneros de hormigas que interactúan con W. auropunctata en cebos de atún.
Interacciones por el recurso alimenticio
19 géneros (Fig. 2) y 40 especies (Apéndice) se encontraron compartiendo los cebos con la pequeña hormiga de fuego. Sobresalen Pheidole, Solenopsis y Crematogaster que en conjunto representaron el 56% de todas las interacciones en el área de estudio. Cinco especies: Pheidole mendicula (Wheeler, 1925) (17%), Solenopsis pollux (Forel, 1893) (9,6%), Crematogaster carinata (Mayr, 1862) (8,1%), Monomorium floricola (Jerdon, 1851) (8,1%) y Camponotus novogranadensis (Mayr, 1870) (5,9%) (Tabla 3), conformaron el 48,7% del total de interacciones.
Partición del recurso alimenticio
La partición del cebo en los diferentes sitios fue afectada significativamente por el biotopo (χβ = 46,226; P = 5,47 x 10−7; g.l. = 9) (Fig. 3). Así, en interior de bosque, la partición del cebo fue más homogénea ya que se encontraron cebos compartidos y cebos monopolizados por una especie. Los biotopos borde de bosque y potrero favorecieron el compartimiento del cebo mientras que la matriz de caña es preferiblemente dominada por una especie.
Abundancia de W. auropunctata y aprovechamiento del cebo por otras especies
La presencia de otras especies de hormigas en los cebos varió significativamente con respecto al número de obreras reclutadas de la pequeña hormiga de fuego (H: 166; P < 0,001; g.l. = 5; n = 358) (Fig. 4). Con una abundancia promedio de 515 (± 40,6) obreras de W. auropunctata por cebo, el recurso no fue compartido con ninguna otra especie de hormiga; pero cuando la densidad de W. auropunctata fue aproximadamente cinco veces más baja (< 100 obreras por cebo), el recurso se compartió hasta con otras cinco especies.
Oferta de cebos y efectividad de atracción para hormigas en el área de estudio.
Número de interacciones entre especies de hormigas que comparten el cebo con W. auropunctata.
Intensidad de competencia
En la tabla 4 se describen los dos índices de habilidad competitiva cuyos valores fluctúan entre 0 y 100%. Por ejemplo, para Crematogaster evallans (Forel, 1907) el número total de ocurrencias en cebos fue de 14 y de éstas en 13 se contaron más de 20 obreras por cebo, lo que resulta en un índice a igual a 93%. Por el contrario, para S. geminata, el número total de ocurrencias en cebos fue de 38 y en 31 de éstas se encontró solamente a esta especie, lo que resulta en un índice b igual a 82%.
Discusión
Los cebos de atún son útiles para estimar composición y riqueza de especies de hormigas que forrajean en el suelo (Bestelmeyer et al. 2000), evaluar competencia (Hölldobler y Wilson 1990) y medir eficiencia de forrajeo y comportamientos competitivos cuando se contabiliza la abundancia (Greenslade y Greenslade 1971). En el presente estudio, la efectividad de atracción de hormigas a cebos fue superior al 90%, resultado similar al obtenido por otros autores en Bosque seco Tropical (Bs-T). Así, Armbrecht y Chacón de Ulloa (1997a) observaron una atracción del 98% en cebos epígeos y arbóreos, y Ramírez (2001) reportó una efectividad del 82%, siendo el segundo método con mayor porcentaje de captura de hormigas, después del método manual o captura directa. Sin embargo, el número de especies puede variar si se utilizan combinaciones de diferentes métodos de muestreo (Delabie et al. 2000).
Las 66 especies de hormigas recolectadas en los cuatro fragmentos de bosque seco representan el 33% del total de especies (200) conocidas para el área de estudio (Chacón de Ulloa y Armbrecht 2006). Este porcentaje es importante ya que los cebos atraen principalmente a hormigas generalistas y pueden excluir a especies que tengan otras preferencias alimenticias como las cortadoras de hojas o depredadoras especializadas (Bestelmeyer et al. 2000). Sin embargo, ocasionalmente se encontraron en los cebos, especies cultivadoras de hongos como Atta cephalotes (Linnaeus, 1758), Cyphomyrmex rimosus (Spinola, 1851) y Trachymyrmex sp. 1. Ocho subfamilias de Formicidae fueron atraídas a los cebos, confirmando las observaciones de Ramírez (2001) y Lozano (2002), para la misma zona de estudio. La subfamilia más diversa fue Myrmicinae con 41 morfoespecies (Apéndice) que representó el 62% de la composición de hormigas y de la cual se registraron 17 de los 27 géneros conocidos para el bosque seco del valle geográfico del río Cauca (Chacón de Ulloa y Armbrecht 2006).
Tres géneros de Myrmicinae fueron los mejor representados en los cebos, Pheidole (12 spp.), Crematogaster (7) y Solenopsis (6), y el género con mayor porcentaje de captura fue Wasmannia con la única especie W. auropunctata, lo que puede estar indicando desplazamiento de otras hormigas en términos de eficacia competitiva, historia natural y explotación de sitios perturbados (Armbrecht y Ulloa-Chacón 2003; López 2005).
La composición de los ensamblajes de hormigas presentó influencia local (Fig. 1); tendencia notable especialmente en los biotopos de bosque (interior y borde) que se agruparon por sitios. Las matrices de tres sitios (El Medio, El Vínculo, San Julián) presentaron composición de hormigas más parecidas entre ellas que con sus bosques vecinos, es decir, las matrices mantienen una diversidad de especies propia que se diferencia de la de los biotopos con cobertura arbórea, indicando un marcado efecto de borde. En esta área de alta perturbación se encuentran pocas especies, las cuales comparten hábitos generalistas y toleran condiciones agresivas como alta intensidad de luz y baja humedad (Pauchard y Alaback 2006).
Índices de habilidad competitiva para las cinco especies de hormigas más abundantes en el área de estudio.
No se encontraron diferencias en cuanto a la composición de hormigas entre los biotopos, pero si se encontraron diferencias significativas entre los sitios. Este resultado proporciona evidencia de que en el área de estudio existe un marcado efecto de sitio y por consiguiente, la composición de la fauna de hormigas puede estar siendo influenciada por variables como grado de perturbación e historia de manejo de cada sitio (Armbrecht y Chacón de Ulloa 1997b).
Las 40 especies que se encontraron compartiendo los recursos con W. auropunctata (Apéndice) se pueden dividir en tres grupos: 1. Especies grandes que forrajean solitariamente como Ectatomma ruidum (Roger, 1861) y Pachycondyla constricta (Mayr, 1884); 2. Especies que reclutan masivamente como algunas Pheidole, Crematogaster y Solenopsis; y 3. Especies que se encuentran en frecuencias muy bajas como Rogeria belti (Mann, 1922) y Megalomyrmex wallacei (Mann, 1916). Tennant (1994) realizó observaciones similares en selvas húmedas de Costa Rica y Panamá; donde W. auropunctata ignoró a las especies grandes y solitarias que acarreaban fuera del cebo pedacitos de atún (grupo 1). También registró la ocurrencia de especies que compiten fuertemente con W. auropunctata en los cebos (grupo 2). Finalmente, el tercer grupo estuvo representado por especies que utilizaron el recurso con pocas obreras y en muchas ocasiones las interacciones no fueron visibles ya que la partición del cebo ocurría espacialmente, una especie forrajeaba en la parte superior del cebo y las otras alrededor o por debajo.
El bosque con menor número de interacciones por el recurso fue San Julián, el cual presentó la riqueza de hormigas más baja y el mayor número de obreras de W. auropunctata por cebo. Armbrecht y Ulloa-Chacón (2003) encontraron que San Julián posee la menor diversidad de mirmecofauna y sugieren que la gran abundancia y alta agresividad interespecífica de la pequeña hormiga de fuego incrementa el desplazamiento de otras especies de hormigas. W. auropunctata es una especie muy eficiente en encontrar los recursos y generalmente es la primera en dominarlo, además, sus nidos cubren una gran área y reclutan en masa (Clark et al. 1982; Meier 1994; Tennant 1994).
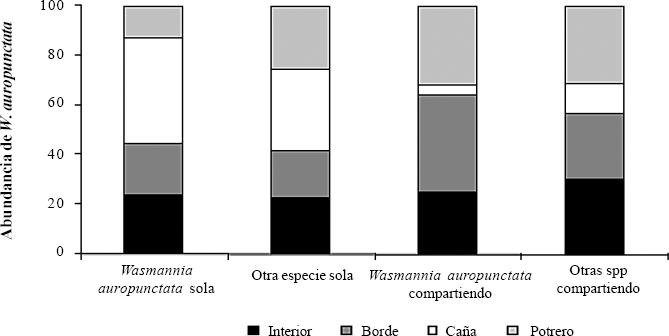
Frecuencia (%) de partición del cebo según el biotopo en toda el área de estudio.
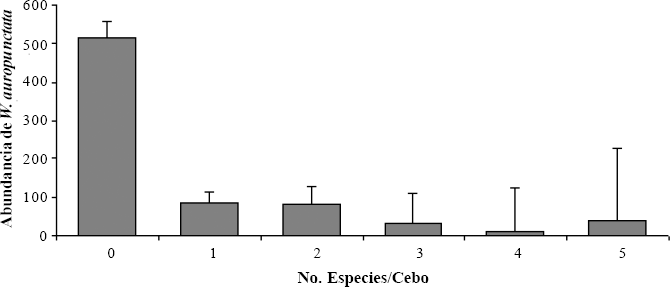
Otras especies de hormigas en cebos y abundancia promedio (± desviación estándar) de obreras de W. auropunctata.
Los biotopos que presentan cobertura arbórea proporcionan condiciones más homogéneas (sitios para anidar, alimento, microclima) y por lo tanto, es más probable encontrar diferentes tipos de partición del recurso en similar proporción. En contraste, las matrices (caña de azúcar y potrero) son ambientes donde se facilitan las condiciones para que exista dominancia de una especie sobre el recurso alimenticio; por ejemplo, en caña de azúcar, W. auropunctata encuentra disponibilidad de recursos y baja diversidad de hormigas, lo que le posibilitaba dominar ese ambiente. En potrero, a pesar de que la diversidad de hormigas también es baja, las condiciones de humedad no son adecuadas para que la pequeña hormiga de fuego se establezca y domine sobre otras especies, lo que explica la dominancia de S. geminata, una especie que aprovecha la perturbación y su hábitat principal es la matriz transformada (Davidson 1998).
En áreas muy perturbadas como el Bs-T de la cuenca alta del Río Cauca, W. auropunctata alcanza abundancias muy grandes y monopoliza el cebo cuando en promedio están presentes 500 obreras. Armbrecht y Ulloa-Chacón (2003), reportan que áreas donde la densidad de la pequeña hormiga de fuego es alta, la riqueza en el ensamblaje de hormigas disminuye significativamente. Adicionalmente, Clark et al. (1982) reportan que en el 35% de los casos donde W. auropunctata estuvo presente en los cebos, los grupos de forrajeo estaban compuestos por más de 128 obreras y excluían completamente a sus competidoras. Otro estudio similar, en la isla de Nueva Caledonia (Le Breton et al. 2005), encontró que en áreas invadidas todos los cebos son descubiertos por obreras de W. auropunctata, y rápidamente los ocupan con un gran número de individuos como resultado de su efectivo reclutamiento en masa. Además, cuando la especie alcanza densidades de más de 100 obreras no se observan otras especies de hormigas sobre los cebos a pesar de que algunas exploradoras se observan cerca del cebo.
Entre las cinco especies más abundantes se encuentran diferencias en los tipos de competencia que pueden presentar. S. geminata y C. evallans presentaron los índices de competencia más altos; estas especies son dominantes en los sitios donde se encuentran y pueden excluir a sus competidores por agresión física y siendo más eficientes en aprovechar los recursos. W. auropunctata fue la especie dominante en el estudio y su abundancia fue superior comparada con las otras especies (Apéndice); esta dominancia es lograda principalmente por mecanismos de explotación ya que presenta un poderoso reclutamiento en masa cuando detecta una fuente de alimento lo que le permite aprovechar eficientemente los recursos (Clark et al. 1982; Passera 1994; Armbrecht y Ulloa-Chacón 2003; Le Breton et al. 2005). Adicionalmente, la pequeña hormiga de fuego presenta características que le ayudan a excluir a sus competidoras como unicolonialidad, poliginia, baja agresión intraespecífica y alta agresión interespecífica (Passera 1994; López 2005); además de haber sido ubicada junto a las especies extirpadoras (Wilson 1971), las cuales desplazan a las otras especies mediante mecanismos competitivos (Clark et al. 1982; Meier 1994; Le Breton et al. 2005).
Conclusiones
Wasmannia auropunctata fue la especie de hormiga dominante tanto en la utilización del recurso alimenticio como en el número de obreras. Esta dominancia es alcanzada, en parte, por su habilidad competitiva, principalmente por explotación. Adicionalmente, la abundancia de la pequeña hormiga de fuego determina, en cierta medida, el aprovechamiento del recurso alimenticio por parte de otras hormigas, por lo cual, se refuerza la idea de que esta especie es indicadora de baja diversidad de hormigas en la cuenca alta del Valle del río Cauca. Las matrices (potrero y caña) poseen una diversidad mucho menor que las de los biotopos con cobertura arbórea; además, se observa que las relaciones entre hormigas en los biotopos productivos son menos complejas, ya que la intensidad de competencia es más baja al existir especies únicas que aumentan la probabilidad de que el recurso alimenticio sea monopolizado.
Debido a que la composición de hormigas probablemente está determinada por la localidad, la pérdida de alguno de los fragmentos de bosque supondría la desaparición de algunas especies a escala local y regional. Por consiguiente, sería importante aumentar la conectividad estructural entre parches así como evitar la degradación continuada y mejorar las formas de manejo de las matrices.
Agradecimientos
Al Instituto de Investigación de Recursos Biológicos Alexander von Humboldt por su apoyo financiero a través del convenio No. 192 con el grupo de investigación en Biología, Ecología y Manejo de Hormigas de la Universidad del Valle. Al Dr. Jacques H. Delabie por haber facilitado la determinación de algunas especies de hormigas en el CEPEC. A los evaluadores cuya revisión fue fundamental para mejorar la versión final del manuscrito. A los propietarios y administradores de los bosques por permitir el trabajo en sus predios: Hilda Sanint (Alejandría), Carlos Ávila (El Medio), Luis Carlos Sadovnik (San Julián) e INCIVA (El Vínculo). A los biólogos Pilar López, Paloma Vejarano, Christian Bermúdez, Néstor Zúñiga y Ana Osorio, por su valiosa ayuda en el trabajo de campo.
Footnotes
Apéndice.
Abundancia de las especies de hormigas atraídas a cebos de atún en cuatro sitios de la cuenca media del río Cauca, Colombia.