
Abstract
Females of the sand fly Lutzomyia ovallesi (Ortiz) (Diptera: Psychodidae) were fed with blood from various species of vertebrates and analyzed to determine energy reserves under laboratory conditions. L. ovallesi specimens were allowed to artificially feed to satiation through chicken membranes on blood from horse, dog, cow, chicken, goat, pig or human. Caloric reserves were calculated spectrophotometrically after females were homogenized in a solution of sodium dichromate and sulfuric acid. The caloric content of L. ovallesi varied according to the type of vertebrate blood on which it had fed. The highest content (cal/insect) was found in females fed on human blood (0.33), followed in decreasing order by dog, pig, cow, chicken, goat and horse (0.26). Statistical analysis showed significant differences (P < 0.05) among sources. The results showed that human and dog blood meals were more nutritionally efficient. The most inefficient diet for L. ovallesi was horse blood manifested by its poor nutritional quality.
Introduction
The ability of any hematophagous insect to survive and transmit pathogens depends principally on its caloric reserves (Van Handel 1972; Magnarelli and Modi 1988; Briegel et al. 2001). The energy requirements of female phlebotomine sand flies (Diptera: Psychodidae) are supplied by three sources: caloric reserves built up during the larval stage and sugar and vertebrate blood ingested as an adult (Van Handel 1972, 1984; Magnarelli and Burger 1984; Magnarelli and Modi 1988; Mostowy and Foster 2004).
Fecundity variations in sand flies according to bloodmeal source may be attributed to significant differences in the caloric content of carbohydrates, lipids, and proteins from the ingestion and metabolization of blood. Large caloric reserves could provide greater potential energy for egg production, oviposition survival, and flight capacity (Magnarelli and Modi 1988; Harre et al. 2001), increasing the biological potential of a specific sand fly population, resulting in increased transmission of Leishmania (Kinetoplastida) (Schlein et al. 1983; Daba et al. 1997; Schlein and Jacobson 1998; Hurd 2003). When available energy reserves in both sexes of the sand flies Lutzomyia longipalpis (Lutz & Neiva) and Phlebotomus papatasi (Scopoli) were quantified, those with access to fructose or sucrose solutions in the laboratory had higher levels than those supplied with labeled glucose. Caloric assays can be used to evaluate larval and adult diets (Magnarelli and Modi 1988).
The sand fly L. ovallesi (Ortiz) is the principal vector of Leishmania braziliensis in western and central Venezuela (Bonfante-Garrido et al. 1991a; 1991b; Feliciangeli 1991) and one of the most important vectors in the Venezuelan Andes (Añez et al. 1988). The purpose of the present study was to determine caloric contents of L. ovallesi fed with vertebrate blood from different sources, under laboratory conditions.
Materials and methods
Results
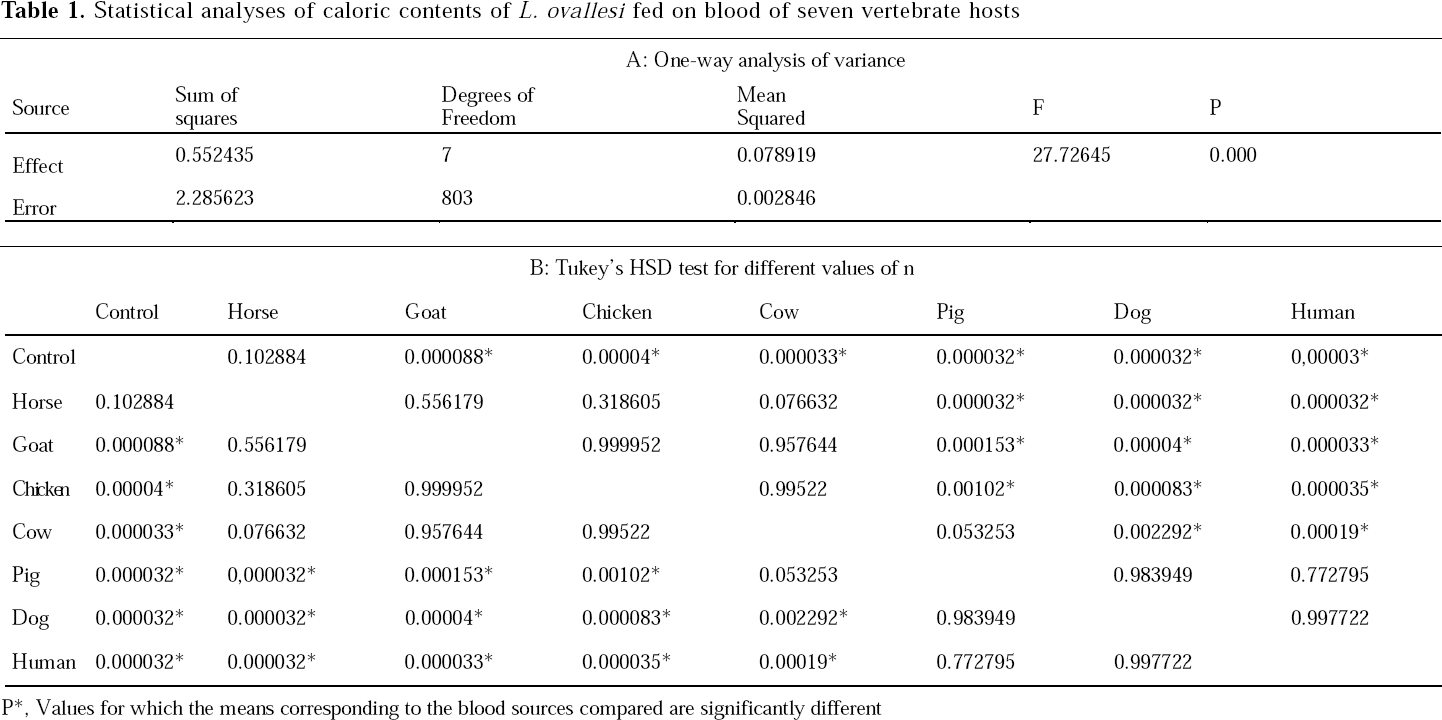
Caloric contents of L. ovallesi fed on blood from each of seven vertebrate species are shown in Figure 1. The highest caloric content (cal/female) was obtained from insects fed on human (x = 0.33; range 0.18-0.47), and the lowest from those fed on horse blood (x = 0.26; 0.15-0.39). In decreasing order of magnitude the caloric content for L. ovallesi fed on different types of blood was as follows: control < horse < goat < chicken < cow < pig < dog < human. Significant differences (P ≤ 0.05) were seen for the following comparisons: cow vs dog, human vs control; pig vs horse, goat, chicken and control; dog vs cow, horse, goat, chicken blood and control; horse vs pig, dog, human and control; and human vs cow, horse, goat, chicken, and control (Table 1).
Discussion and Conclusions
The enormous reproductive potential of hematophagous insects is largely due to the female's success in locating a host, approaching it to feed, utilizing the blood to mature an optimal number of eggs and then finding a suitable site for oviposition. This pattern of behavior also means that females can transmit pathogens between hosts (Briegel 1990).
Ingestion of blood swells the epithelial cells, causing reversible phenomena such as secretion of the peritrophic matrix and liberation of proteolytic enzymes. The main products of blood digestion are amino acids. When blood is digested, the final product is excreted as ammonium urate (Rudin and Hecker 1982; Magnarelli and Burger 1984).
Dichromate solution oxidizes the insect completely, with proteins, carbohydrates, lipids and chitin being converted into carbon dioxide (Van Handel 1984; Magnarelli and Modi 1988). This technique is both rapid and sensitive and can be used to determine the nutritional state of females in a laboratory colony or assess the value of blood from different vertebrate hosts. Based on our results, all the blood sources provided energy for L. ovallesi although the caloric value of sand flies fed on horse blood was not significantly greater than that of unfed flies. These values obtained do not necessarily reflect the total caloric reserves available to the females, since they were gravid; a large proportion of the caloric reserves is used in egg production, with very little used for female nutrition (Rudin and Hecker 1982; Magnarelli and Burger 1984).
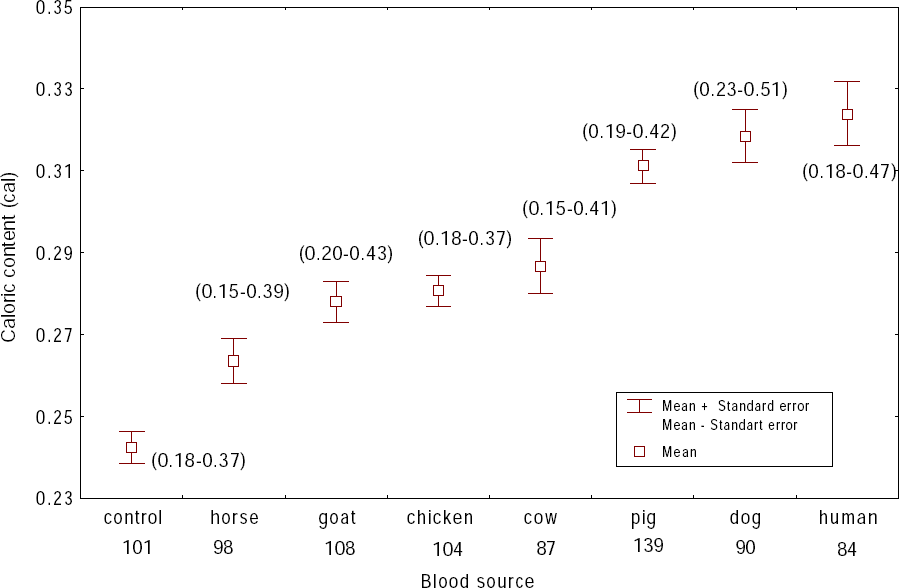
Caloric contents of sand flies L. ovallesi fed on blood from seven vertebrate hosts. Numbers in parentheses are ranges and numbers given below the blood source are values of n.
Statistical analyses of caloric contents of L. ovallesi fed on blood of seven vertebrate hosts
Nasci (1986) reported that large females of the mosquito species Aedes aegypti possess large energy reserves at eclosion, providing them with great flight potential and the ability to contact more hosts and transmit pathogens. However, Landry et al. (1988) found that significant seasonal differences in the body size of Ae. triseriatus had no effect on flight potential or life-span. Harre et al. (2001) found that P. papatasi fed on blood from eight species of mammals and detected no appreciable difference between these hosts with respect to sand fly mortality rates after 24h, number of eggs laid per blood-fed female or egg viability. Laboratory-reared males and females of both L. longipalpis and P. papatasi which had access to fructose or sucrose solutions had greater mean available energy reserves (x = 1.3 cal/insect) than individuals provided with glucose solution (x= 0.55). Available caloric reserves were low in natural populations of P. papatasi and these insects probably must feed repeatedly on vertebrate hosts and sugar sources to obtain sufficient nutrients for survival and reproduction (Magnarelli and Modi 1988).
Although, a high number of sand fly species have been successfully colonized during the last decade, the factors limiting their productivity and fecundity in the laboratory are unknown (Montoya et al. 1998; Luitgards-Moura et al. 2000). Knowledge about the physiological events taking place in the vector is important in understanding vector-parasite interactions necessary for disease transmission. Nutritional quality of blood varies between host species and may influence egg productivity, reduces development rates, longevity, and fecundity of the insects (Alexander et al. 2002). For an understanding the role of blood meal sources on sandfly biology, physiology, and Leishmania transmission both more field observations and laboratory studies comparing egg productivity of sandflies fed on different hosts, are necessary (Alexander et al. 2002; Hurd 2003).
The compatibility of the sand fly and its specific Leishmania parasite depends on the choice of host animals available, it could be an important factor in the distribution of leishmaniasis (Schlein et al. 1983). The proteins from the blood meal are digested by the sand fly gut. It appears that the enzymatic processes in the sand fly gut, functions differently when triggered by different types of meals, and the blood meal from distinct animal sources can be lethal to Leishmania (Adler 1964). L. tropica infection was inhibited in P. papatasi fed on turkey blood because a relatively high DNAase activity level was induced in the sand fly gut by nucleated erythrocytes (Schlein et al. 1983). However, the blood meals from different species of vertebrates have no deleterious effect on the development of either L. braziliensis and L. amazonensis in the gut of L. migonei; also, parasite development was compatible with digestion, independent of the blood meal source (Nieves and Pimenta 2002). The development of L. infantum infection was associated with suppression of blood protein digestion by sand flies fed on human or dog blood (Schlein et al. 1983; Daba et al. 1997). It also was demonstrated that the rate of blood meal digestion in P. langeroni varied according to the source of the vertebrate blood and Leishmania species involved (Daba et al. 1997).
Very little is known about how these nutrients are used during adulthood. Sand fly reproduction depends on the availability of blood meal sources such as domestic animals and synanthropic species. In endemic areas where some species of domestic animals are sources of blood meals, a higher number of sand fly vectors with more parasites occur. This fact provides a selective advantage to the vector competence in transmitting Leishmania to vertebrates. This was possible due to the relatively high isoleucine content in rodent blood, as opposed to its role as a limiting factor for oogenesis with human blood. Important role of isoleucine explained the results of several previous reports that showed variable mosquito fecundity with different host (Briegel 1990). Similar physiological mechanisms may play a role in the sand flies. Although, feeding on blood from rodents was superior to that from humans with respect to fecundity in Ae. aegypti, it may be sub-optimal energetically (Briegel 1990). L. braziliensis has been found in domestic animals as dogs and equines as well as in wild mammals such as rodents, edentata and opossums (Aguilar et al. 1984; Grimaldi and Tesh 1993). L. ovallesi feeds upon a variety of vertebrate hosts, and could be considered as an opportunistic species (Añez et al. 1988; Nieves et al. 2004). Based on the results of the present study, there are significant differences in the caloric contents of female L. ovallesi fed on blood from different sources, with human, dog, and pig blood providing most energy. It might therefore, benefit females of this species to feed preferentially on these hosts. Values for females fed on horse blood were as low as those in the control group, which had been fed only in sugar. Further studies are required to determine how certain dietary factors affect vector potential and their consequences for Leishmania transmission. This information may enable health authorities to adopt policies concerning the presence of domestic animals in endemic areas and may comprise factor risk for Leishmania transmission.
Footnotes
Acknowledgments
We thank Carlos Araque for assistance with the laboratory colony; Luis Chavez for his invaluable help and cooperation; Efrain Entralgo, Guillermo Bianchi and Paolo Ramoni for guidance in statistical analysis; Leyda Quintero for help in providing blood samples; and the CDCHTULA (C-1278-04-03-B) and CONICIT (S1-2000000818) for providing financial support for this work.