
Abstract
The chaetotaxy of fourth instar larvae of Lutzomyia evansi, obtained from laboratory conditions, is redescribed and illustrated. Measurements of the setae are also provided. The comparative morphological study suggests that fourth instar larvae of L. evansi, as well as the other species of the verrucarum series, can be placed in Group 4 of the neotropical phlebotomine which posses antennae with a short basal segment and an ovoid distal segment, which seems to support the correct proposed groupings based on characters of the imagos. The significance of the importance to standardize the nomenclature of sand fly chaetotaxy and use of a single chaetotaxy system for their description is discussed.
Introducción
Un hecho que resalta en los estudios taxonómicos y sistemáticos de la subfamilia Phlebotominae (Diptera, Psychodidae), es el predominio de las estructuras imaginales en la conformación de subgéneros y los grupos de especies. Esto ha determinado que se desconozca en gran medida la relevancia de los estadios pre-imaginales en la taxonomía y sistemática de la subfamilia y que como bien lo señalan Bejarano et al. (2003), todavía no se disponga de claves taxonómicas para la identificación de los mismos. Probablemente esto se deba a la dificultad para detectar las larvas en sus nichos naturales y en menor medida a que su cría y mantenimiento en el laboratorio es laboriosa.
A esta problemática no escapa el grupo verrucarum del género Lutzomyia França, 1924. Este taxón comprende alrededor de 48 especies divididas en cuatro series, incluyendo verrucarum, townsendi, serrana y pia (Galati 1995, 2003; Bejarano et al. 2003). El grupo tiene un papel relevante en la salud pública, debido a que muchas especies son vectores comprobados o potenciales al ser humano de protozoarios del género Leishmania, la bacteria Bartonella bacilliformes y de los arbovirus del género Phlebovirus, agentes etiológicos de los diversos tipos de leishmaniosis, bartonelosis (enfermedad de Carrión) y la fiebre de los flebótomos (sandfly fever), respectivamente (Young y Duncan 1994; Bejarano et al. 2003; Acevedo y Arrivillaga 2008).
Lutzomyia evansi (Núñez-Tovar, 1924) es una especie de flebotomíneo perteneciente a la serie verrucarum del grupo verrucarum, que incluye además a L. verrucarum (Townsend, 1913), L. colombiana (Ristorcelli y Van Ty, 1941), L. ovallesi (Ortiz, 1952), L. nuneztovari (Ortiz, 1954), L. nevesi (Damasceno y Arouck, 1956), L. andina Osorno, Osorno y Morales, 1972, L. disiuncta Morales, Osorno y Osorno, 1974, L. moralesi Young, 1979, L. aulari Feliciangeli, Ordóñez y Manzanilla, 1984, L. deorsa Pérez, Ogusuku, Monje y Young, 1991, L. maranonensis Galati, Cáceres y Le Pont, 1995, L. cajamarcensis Galati, Cáceres y Le Pont, 1995, L. antioquiensis (Wolff y Galati, 2002) y L. falcaorum (Brazil y Andrade Filho, 2002) (Bejarano et al. 2003). A L. evansi se le considera un vector comprobado de leishmaniosis visceral (LV), con una distribución neotropical que abarca Centroamérica (Nicaragua, El Salvador, Honduras, Costa Rica y México) y Suramérica (Colombia y Venezuela) (Travi et al. 1990; Young y Duncan 1994; Bejarano et al. 2003; Ibáñez-Bernal et al. 2004). En Venezuela se ha capturado en 14 de las 23 entidades federales del país en altitudes comprendidas entre los 100 y 1200 m y en una amplia variedad de zonas de vida bioclimáticas (Feliciangeli 1988).
Cuando se hace un análisis detallado de las relativamente pocas descripciones de las larvas de Phlebotominae, y particularmente en el grupo verrucarum, llama la atención que la morfometría de las setas es aun más escasa. Esto ha limitado la cuantificación, mediante técnicas numéricas multivarian-tes (e.g., análisis de componentes principales o análisis de agrupamiento), de la similitud morfológica entre los grupos o subgéneros de flebotominos, toda vez que la morfometría de caracteres de imagos ha resultado exitosa para resolver problemas taxonómicos en este taxón (Bejarano et al. 2003).
Hasta el presente, de las 48 especies que integran al grupo verrucarum sólo se ha descrito la quetotaxia de cinco especies, incluyendo a L. serrana, L. youngi, L. ovallesi, L. verrucarum y L. evansi, lo que equivale al 10,42% del grupo (Bejarano et al. 2003; Galati 2003). Sin embargo, la descripción de las larvas de estadio IV de L. evansi hecha por Montoya-Lerma (1996) a partir de hembras capturadas en Colombia, posee la limitación de que no se detectaron o detallaron, o no se ilustraron, setas u otras estructuras anatómicas cuya importancia en la taxonomía del grupo aún falta por dilucidar en su real dimensión.
En virtud de lo planteado, en el presente trabajo se redescribe e ilustra la morfología y quetotaxia de larvas del estadio IV de L. evansi, obtenidas a partir de la colonización y cría de hembras capturadas en la región andino-venezolana. Así mismo, se dan las medidas de las setas con las que se espera en futuros trabajos cuantificar las diferencias y afinidades morfológicas y taxonómicas interespecíficas e intraespecíficas en la quetotaxia del grupo, mediante la implementación de técnicas estadísticas de análisis multivariante.
Materiales y Métodos
Las hembras paridas de L. evansi se capturaron utilizando el método de la trampa lumínica de Shannon, en la población “Montañas de Peraza” (9°27’55”N, 70°31’33.3”W), ubicada a 403 m en el Municipio Pampán del estado Trujillo, región andino-venezolana. La región posee una zona de vida bioclimática correspondiente al Monte Seco Tropical, con cultivos extensivos de piña y es un área endémica para leishmaniosis tegumentaria (LT) y visceral (LV) (Ewell et al. 1976; Oviedo et al. 1999).
Las larvas de estadio IV se obtuvieron mediante técnica de colonización y cultivo propuesta por Añez y Oviedo (1985). Las larvas se sacrificaron con vapores de cloroformo, se clarificaron en solución de Nesbitt durante 12 - 24 horas y se montaron en líquido de Berlesse. La quetotaxia se describió según nomenclatura y sistema de numeración empleados por Forattini (1973), sistema mejorado por dicho autor a partir de las propuestas nomenclaturales de Barretto (1941) y de Hanson (1968). El sistema de Forattini (1973) es muy sólido pues se basa en un amplio rango de larvas IV de especies flebotominas del Neotrópico a diferencia de otros propuestos como el de Ward (1976) que se creó exclusivamente para los subgéneros Nyssomyia y Psychodopygus, o el de Leite y Williams (1996, 1997) para larvas de L. longipalpis.
Los detalles de cada seta se identificaron a 400 X con microscopio fotónico Axiostar Plus (Carl Zeiss). Las estimaciones morfométricas se hicieron con un micrómetro ocular previamente calibrado, dibujándose las estructuras morfológicas con una cámara lúcida. Las medidas, todas expresadas en micras (μm), se presentan incluyendo la media aritmética y el intervalo de las mismas en paréntesis.
Resultados y Discusión
Descripción de la quetotaxia larval de L. evansi.
La descripción se basa en el examen de 18 larvas IV obtenidas de distintas hembras. El material entomológico se encuentra almacenado en la colección de Entomología Médica del Laboratorio de Entomología, Parasitología y Medicina Tropical (L.E.P.A.M.E.T.) del Centro de Investigaciones Biomédicas (C.I.B.), Universidad Nacional Experimental “Francisco de Miranda” (UNEFM), Coro, Estado Falcón, Venezuela.
Longitud promedio del cuerpo, desde el margen anterior de la cápsula cefálica hasta el margen posterior del peine del noveno segmento abdominal 2855,6 μm (rango 2240-3320).
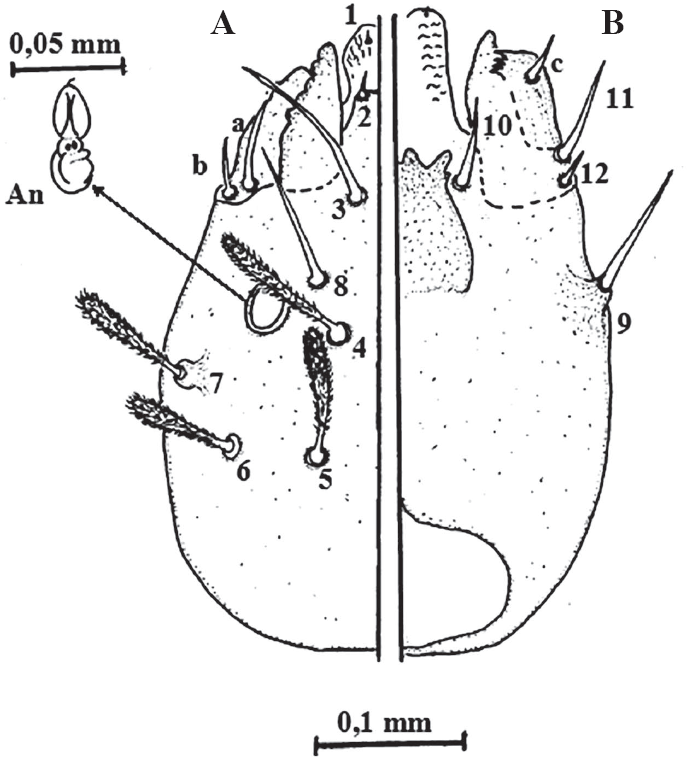
Quetotaxia de la región cefálica dorsal (A) y ventral (B) de larvas del estadio IV de Lutzomyia evansi. An= antena. Las setas a, b y c, no aparecen descritas en el sistema de numeración de Forattini (1973).
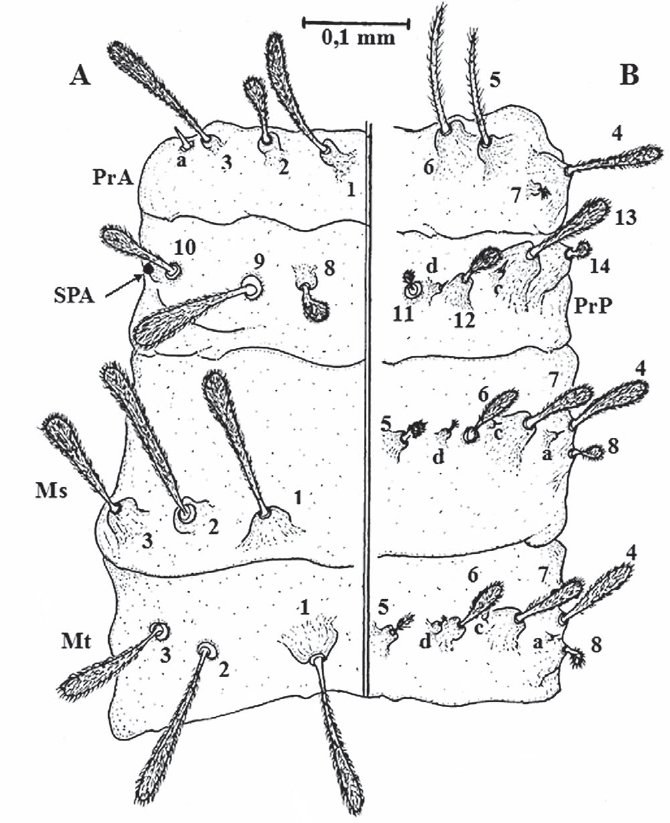
Quetotaxia de los segmentos torácicos dorsales (A) y ventrales (B) de larvas del estadio IV de Lutzomyia evansi. PrA= protórax anterior; PrP= protórax posterior; Ms= mesotórax; Mt= metatórax; SPA= espiráculos anteriores.
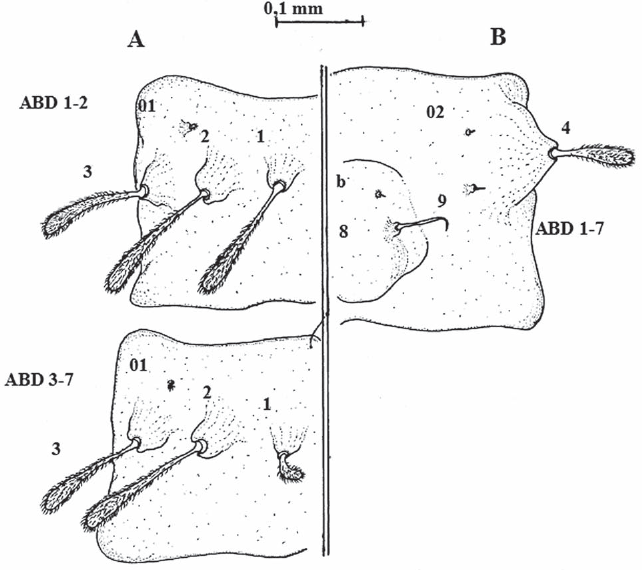
Quetotaxia de los segmentos dorsales (A) y ventrales (B) abdominales de larvas del estadio IV de Lutzomyia evansi. ABD= segmentos abdominales 1, 2, 3 y 7.
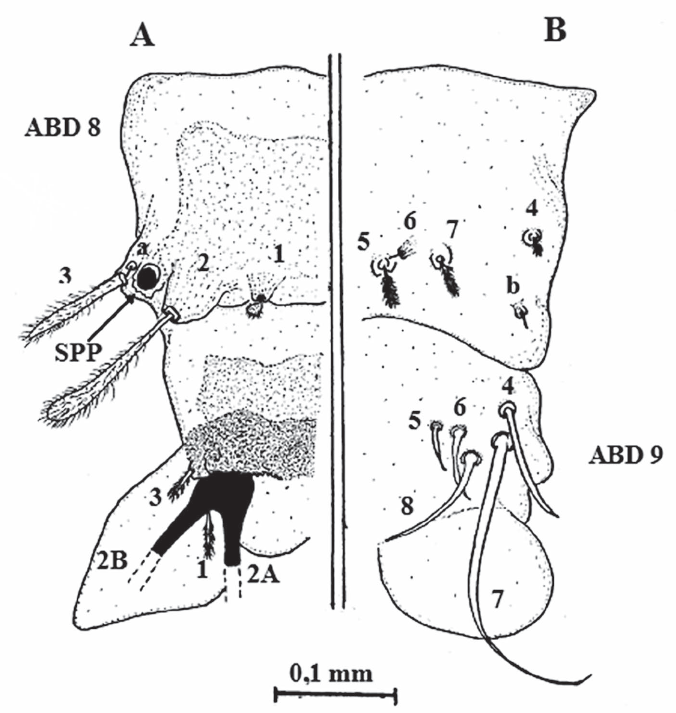
Quetotaxia de los segmentos abdominales 8 y 9 dorsales (A) y ventrales (B), de larvas del estadio IV de Lutzomyia evansi. ABD 8 = segmento abdominal 8; ABD 9= segmento abdominal 9; 2A y 2B= setas caudales; SPP= espiráculos posteriores.
En la Tabla 1 se presentan las medidas (μm) con media aritmética y rangos de las setas descritas.
De las cinco especies flebotominas del grupo verrucarum a las cuales se les ha descrito la quetotaxia, tres pertenecen a la serie verrucarum, incluyendo L. ovallesi, L. evansi y L. verrucarum; sin embargo, se debe decir que a esta última especie flebotomina sólo se le han descrito las setas cefálicas (Forattini 1973; Ogusuku y Pérez 1995; Bejarano et al. 2003). Como ya se mencionó, Montoya-Lerma (1996) describió la quetotaxia de los estadios inmaduros de L. evansi, incluyendo las larvas de IV estadio, que fueron descritas a partir de tres especímenes. Cuando se analiza la descripción de la región cefálica en dicho trabajo, se destaca en primer lugar que no se describe ni se ilustra la forma de las antenas y además se indica, contrariamente a lo observado en el presente trabajo, que el “segmento proximal es ligeramente más largo que el distal”, aunque no se ofrecen las medidas. En segundo lugar, no se describieron ni ilustraron las setas 1 y 2, así como tampoco las señaladas en el presente trabajo como a, b y c, y que tampoco aparecen señaladas en los sistemas de numeración de quetotaxia larval hasta ahora conocidos. Para Arrivillaga et al. (1999) estas setas forman parte de la región bucal y no deberían utilizarse para la quetotaxia de las larvas IV de Lutzomyia, por lo que en dicho trabajo, basado en el estudio de cinco especies flebotominas que incluían a L. evansi, propusieron un nuevo sistema de nomenclatura, aunque sólo para la cápsula cefálica. En relación con las regiones torácicas y abdominales, Montoya-Lerma (1996) tampoco describió ni ilustró las setas: a, c, d y 7 (protórax); a, c, d y 8 (mesotórax); a, c, d y 8 (metatórax); 01, 02 y b (segmentos abdominales 1-7), y 6 y b (segmento abdominal 8).
Medias e intervalos (μm) de las longitudes de setas del IV estadio larval de Lutzomyia evansi (N= 18).
(-): seta ausente,
= corresponde a las setas ventrales, que son similares en los segmentos abdominales 1 y 7,
= En la región abdominal dorsal, la seta 1 de los segmentos 3-7 es de menor longitud que la de los segmentos 1-2, +=setas de la región cefálica que no aparecen descritas en el sistema de numeración de Forattini (1973).
De acuerdo con los criterios de Leite y Williams (1996), las larvas del estadio IV de los flebotominos neotropicales se pueden agrupar en cuatro categorías, tomándose como principal criterio la morfología de sus antenas. De acuerdo con este sistema las larvas del estadio IV de L. evansi se ubicarían en el grupo cuatro, por presentar un tubérculo antenal anular, un segmento basal corto y el distal ovoide (Leite y Williams 1996). Similar ubicación tendrían L. ovallesi y L. verrucarum, de acuerdo con las descripciones dadas por Hanson (1968) y Ogusuku y Pérez (1995), lo que apoyaría la ubicación de L. evansi y sus congéneres en la serie verrucarum según datos moleculares (Bejarano et al. 2003).
A la luz de lo comentado, pensamos que no debería subestimarse la importancia de las setas o de cualquier otro carácter anatómico (e.g., sensilas, espiráculos), hasta no tener un número significativo de descripciones que permitan hacer comparaciones para así llegar a conclusiones o aseveraciones más acertadas. En este sentido, son ilustrativos los estudios hechos por Leite y Williams (1996, 1997) en larvas I y IV de L. longipalpis. En efecto, estos autores detectaron mediante alta resolución por Microscopía Electrónica de Barrido (MEB) setas, sensilas y hasta tubérculos parecidos a espiráculos en el tegumento de las larvas, algunas de las cuales aún no se conoce su significado taxonómico y sistemático y otras que pueden ser distintivas a nivel específico y genérico en Phlebotominae.
Debido a la detección de estas estructuras mediante MEB hasta ahora no reveladas en los sistemas quetotáxicos comúnmente implementados, Leite y Williams (1996, 1997) debieron adaptar y modificar el sistema de numeración propuesto por Barreto (1941) para resolver las necesidades del momento, tal como han hecho otros autores para otros grupos de flebotominos, e.g., Ward (1976) para Nyssomyia y Psychodopygus. Por lo tanto, como han venido insistiendo los autores del presente trabajo desde hace más de dos quinquenios (Cazorla y Oviedo 1998), es conveniente modificar y homogenizar los sistemas de numeración hasta ahora propuestos para la setación de Phlebotominae, lo cual se logrará cuando se realicen una cantidad apreciable de nuevas descripciones.
Footnotes
Agradecimientos
Los autores agradecen al personal técnico del Laboratorio de Biología de Lutzomyia del Insectario “Pablo Anduze”, Instituto Experimental “José Witremundo Torrealba”, Núcleo Universitario “Rafael Rangel”, Universidad de Los Andes, Trujillo, estado Trujillo, Venezuela, por su colaboración en las capturas de campo y la cría de los flebotominos.