
Abstract
Spodoptera frugiperda (J.E. Smith), is an insect pest which represent economic lost for different agricultural crops in our country. Instituto de Biotecnología from the Universidad Nacional de Colombia, have been interested in develop some strategies for the biological control of this pest, for instance it have been evaluating the biopesticide activity of native strains of Bacillus thuringiensis (Berliner) on this pest insect. Into this process it is important to remark the IBUN28.5 strain because it has a higher biopesticide activity against S. frugiperda, than the Bt subsp aisawai HD137 standard strain. In order to obtain colonies with different content of cry genes, and evaluate the role of each gene in the biopesticide activity, it was used the technique of curing plasmids. It was used acridine orange, etidium bromide, SDS, temperature and in some cases subculturing, like curing agents for the native strain IBUN28.5 and the standard strain HD137. The selection of the cured derivatives was made by microscopic observation, plasmid profiles, insecticidal cristal proteins profiles and PCR analysis. Selected strains were tested in bioassay against first instar larvae of S. frugiperda, by using two concentration of freezing dried proteins: 240 y 480 ng/cmβ. Percentage of mortality was the bioassay's response. It were obtained some colonies without a plasmid of around 80 MDa to the native strain and 77 MDa to the standard strain, both plasmids have the crylAb gene. Other derivatives lost cry1Ab, cry1C and cry1D genes, but it was not discovered which plasmid contain last two genes. The bioassay results suggest, that whole cured genes have an important role in the biopesticide activity of the IBUN28.5 and the standard strain Bt aisawai HD137. In addition to that, the results suggest that the difference in the biopesticide activity of both strains, is because of intrinsic characteristics of the cry1C and cry1D genes.
Introducción
Bacillus thuringiensis (Berliner) es el mi croorganismo más ampliamente emplea do como estrategia de control biológico a escala mundial (Feitelson y Kim 1992), de ahi el gran número de productos comer ciales a base de este microorganismo y de centros de investigación en todo el mun do, que poseen programas relacionados con la búsqueda y caracterización de ce pas nativas para el control de plagas loca les de los diversos países (Bravo et al. 1998; Hossain et al. 1997; Chak et al. 1994; Chilcot y Wigley 1993). El Instituto de Biotecnología de la Universidad Nacional ha venido desarrollando una colección de cepas nativas de Bt aisladas de diferentes partes del país. Las cepas de esta colec ción son sometidas a un proceso de carac terización riguroso que comprende desde la identificación molecular de los genes asociados a su patogenicidad, hasta su actividad biopesticida a nivel de campo, si la cepa amerita avanzar hasta esta instan cia en el proceso de caracterización.
La cepa nativa IBUN28.5 ha sido seleccio nada dentro de este proceso de caracteri zación, por su mayor actividad biopesticida contra larvas de S. frugiperda a nivel de laboratorio y campo, en comparación con la cepa de referencia HD137 (Grosso et al. 1997; Uribe et al. 1998). Como es bien sabido la especificidad y actividad bio pesticida en Bt está determinada por el contenido de genes cry dentro de su genoma, en -este sentido es importante tener en cuenta que ambas cepas mencio nadas poseen los genes crylAa, crylAb, cry1B, crylC y crylD. De acuerdo con la literatura, la actividad específica de estas cepas contra S. frugiperda reside princi palmente en los genes cry1C y cry1D (Aranda et al. 1996; Bravo et al. 1998). Esto sugiere que la diferencia en la activi dad biopesticida de estas dos cepas está basada en los niveles de expresión o la secuencia especifica de los genes que po seen la actividad contra S.frugiperda. Aun que cabe mencionar que el papel del gen cry1Ab no ha sido suficientemente aclara do, ya que si bien la mayoría de autores coinciden en que dicho gen solo no posee actividad contra S. frugiperda. algunos sugieren que si posee actividad (Bohorova et al. 1997) y otros proponen que su efec to reside simplemente en un efecto sinérgico, debido al incremento en la solubilidad de las proteínas que si poseen actividad biopesticida (Aronson et al. 1991).
Debido e la importancia de S. frugiperda como plaga de diferentes cultivos de inte rés económico en Colombia (Vélez 1997) y el potencial de Bt como alternativa de con trol de esta plaga, es de gran importancia establecer estudios de caracterización es pecíficos que permitan conocer, a nivel genético, en que residen las diferencias de actividad biopesticida entre ambas cepas. Gracias a que la distribución de los genes cry dentro del genoma de Bt es principal mente a nivel plasmídico (Faust et al. 1983; González et al. 1981; González y Carlton 1980), en el presente estudio se sometie ron ambas cepas (Bt subsp. aizawai HD137 e IBUN28.5) a la técnica de curado de plásmidos, con el objeto de obtener colonias derivadas de las cepas silvestres, que presentaran diferente contenido de genes cry en su genoma, para así poder establecer relaciones entre la pérdida de algunos genes y la actividad biopesticida de las mismas.
Materiales y Métodos
Los experimentos de curado de plásmidos se realizaron empleando la temperatura como agente físico de curado y los agen tes naranja de acridina y bromuro de etidio (EtBr), (agentes intercalantes de ADN) y dodecil sulfato de sodio (SDS) (surfactante que altera la permeabilidad de la mem brana) como agentes químicos de curado. Así mismo se evaluaron dos combinacio nes de estos agentes: naranja de acridina SDS y EtBr-SDS con él ánimo de observar el efecto de ambos agentes mezclados. Por otra parte se buscó optimizar la inducción de curado de plásmidos empleando en al gunos tratamientos la técnica de subcultivo como herramienta complemen taria a los agentes de curado (labia 1). Al término de cada uno de estos tratamien tos se realizaron diluciones seriadas en agua DDE, las cuales se sembraron en agar nutritivo. A las 24 horas de incubación, se seleccionaron 50 colonias de cada experi mento de la dilución que presentara un crecimiento entre 30 y 300 unidades formadoras de colonia (o menos en el caso de no crecer las 50 colonias). Todas las colonias fueron analizadas a nivel micros cópico. Cabe mencionar que gracias a que la cepa de mayor interés por su actividad biopesticida es la cepa nativa IBUN28.5, ella fue empleada para realizar la optimización del procedimiento de cura do. Una vez seleccionados los mejores pro cedimientos, éstos fueron evaluados en la cepa de referencia HD137. En la tabla 1 se hace un resumen de los diferentes trata mientos empleados en el curado de plásmidos. La mayor parte de los experi mentos fueron realizados en medio líqui do excepto algunos que están indicados en la labia 1.
Resumen de los diferentes tratamientos de curado de plásmidos empleados con la cepa nativa de B. thuringiensis IBUN28.5 y la cepa de referencia HD137
Experimento realizado con la cepa nativa de B. thuringiensis IBUN28.5 y la cepa HD137.
La concentración del agente de curado se iba incrementando a medida que se avanzaba en los subcultivos.
3 Las rondas se realizaron cada 24 horas.
*Experimentos realizados en agar nutritivo (el experimento con incrementos de concentración de SDS se realizó en medio líquido).
Resultados y Discusión
De acuerdo con el diseño metodológico propuesto, la cepa nativa de Bt IBUN28.5 y la cepa de referencia Bt subsp aizawai HD137 fueron tratadas con diferentes agen-tes de curado (Tabla 1). En la Tabla 2 se muestra un resumen de los resultados ob-tenidos con cada uno de las condiciones de curado ensayadas en este estudio. En la tabla 2 merece la pena resaltar el bajo ren-dimiento obtenido con los diferentes trata-mientos de curado de plásmidos, ya que menos de 10% de las colonias analizadas presentaron la pérdida de alguno de sus plásmidos, además, a pesar de la diversi-dad de experimentos llevados a cabo, no se logró generar colonias totalmente acristaliferas con ninguna de las dos cepas sometidas a estos procedimientos.
Resultados obtenidos con los diferentes tratamientos de curado de plásmidos con la cepa nativa IBUN28.5 (experimento 1 a 19) y la cepa de referencia HD137 (experimentos 20 a 23)
: Colonias que presentan menor cantidad o tamaño y/o diferente forma de cristal que la cepa silvestre.
: Colonias que pierden uno o más plásmidos en relación con la cepa silvestre.
: Naranja de acridina
: Bromuro de etidio
: No se realizó.
El bromuro de etidio (EtBr) y la naranja de acridina (NA), son agentes intercalantes de ADN empleados como agentes de curado de plásmidos en Bt y otras especies de bac-terias (Srivinas et al. 1997; Dussan et al. 1997; Sekar et al. 1987); sin embargo, en este estudio ninguno de los tratamientos con estos agentes indujeron pérdida de plásmidos en las cepas de Bt empleadas. La efectividad baja de estos agentes de curado, posiblemente resida en la agresi-vidad de los tratamientos. Esta agresivi-dad se manifiesta por la disminución en la viabilidad celular, representada por la in-capacidad de crecimiento de las células de Bt en diluciones de siembra más altas de 10−2 (Tabla 2). Un ejemplo de esto se ob-serva para el caso de EtBr, el cual cuando es empleado a 50 mg/mll y temperaturas entre 37°C y 42°C la biomasa celular recu-perada no permitió un crecimiento supe-rior a la dilución de siembra de 101, mientras que el mismo EtBr evaluado a una menor concentración (1 mg/ml) y a 42°C permitió un crecimiento hasta la di-lución de 10−5 (Tabla 2). Situación similar ocurre con la NA donde el cultivo va per-diendo viabilidad a medida que se incre-menta la temperatura de incubación (Tabla 2). Estos resultados si bien no descartan los agentes intercalantes de ADN como al-ternativa para inducción de curado de plásmidos en Bt, si sugieren que las con-diciones aquí evaluadas no son idóneas para este propósito.
Las condiciones de experimentación que permitieron obtener colonias curadas de plásmidos tanto para la cepa nativa IBUN28.5 conio para la cepa de referencia HD137 estuvieron asociadas al SDS (a concentraciones crecientes entre 0.002% y 0.012% ο una concentración fija de 0.02%), una temperatura de 42°C y/o la técnica de subcultivo o repiques sucesivos en agar o medio líquido.
En la tabla 2 se observa que los experi-mentos con la mezcla entre SDS y tempe-ratura, o SDS y subcultivos fueron las condiciones más exitosas para la induc-ción de pérdida de plásmidos en ambas cepas, ya que el total de las colonias cura-das fueron obtenidas por alguna de estas técnicas. Estos resultados complementan las condiciones sugeridas por González et al. (1981) y Mahillon et al. (1988), los cua-les evalúan estas condiciones de forma aislada.
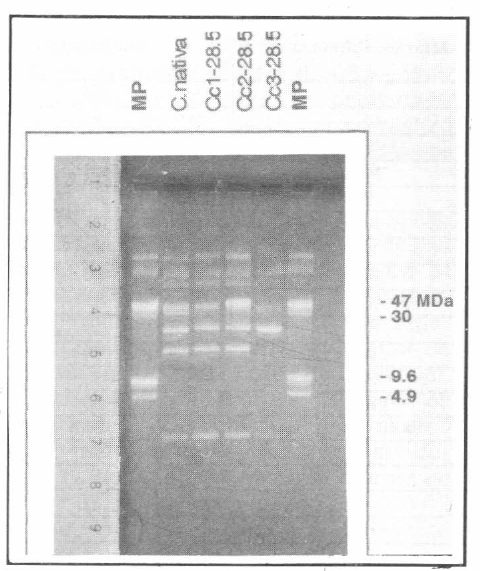
Perfil plasmídico de las colonias curadas obtenidas de la cepa nativa IBUN 28.5. MP: Marcador de peso molecular (cepa Bt subsp. kurstaki HD1). C. nativa: Cepa IBUN 28.5 sin tratar. Cc1-28.5: colonia sin el plásmido de 80 MDa. Cc 2-28.5 colonia sin el plásmido de 80MDa. Cc3-28.5: Colonia sin plásmidos de 95 MDa y 17 MDa.
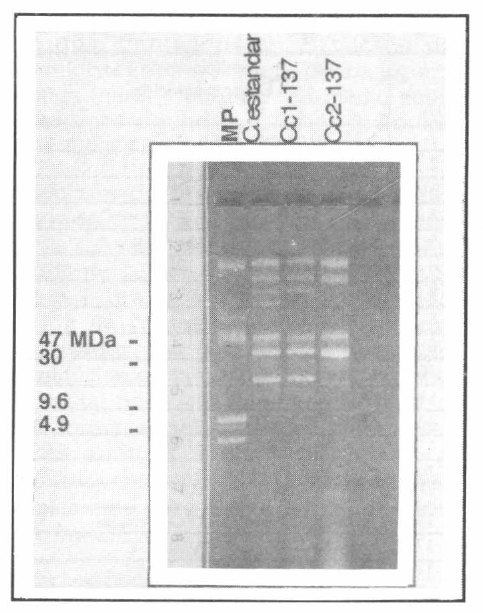
Perfil plasmídico de las colonias curadas obtenidas de la cepa de referencia Bt subsp. aizawai HD 137. MP: Marcador de peso molecular (cepa Bt subsp. kurstaki HD1). C. estándar: cepa de referencia Bt subsp. aizawai sin tratar. Cc1-137: colonia sin el plásmido de 77 MDa. Cc2-137: Colonia sin plásmidos de 99 MDa y 19 MDa.
A pesar de esto, para las colonias seleccio-nadas por la falta de 1 o más plásmidos, el perfil de proteínas sugiere la pérdida de una pequeña banda ubicada alrededor de 131 kDa, la cual coincide con el peso molecular de la proteína Cry1Ab (resulta-dos no mostrados). Dada la dificultad para sacar alguna conclusión con este tipo de resultados era necesario realizar una prue-ba más contundente. Las figuras 3 y 4 muestran los resultados de PCR de las ce-pas silvestres empleadas en este estudio, así como los perfiles de genes cry de las cepas curadas. Estos resultados son muy importantes ya que permiten establecer asociaciones entre los plásmidos perdidos durante el procedimiento de curado y los genes que no dan reacción positiva en esta prueba. En este contexto los resultados de este estudio sugieren que los plásmidos de 80 MDa y 77 MDa para las cepas IBUN28.5 y HD137 respectivamente, son portadoras del gen cry1Ab. Por otra parte, los genes cry1C y cry1D pueden estar co-dificados en los plásmidos de 95 o 17 MDa para la cepa nativa o en los plásmidos de 99 o 19 MDa para la cepa de referencia, ya que las colonias que pierden los tres plásmidos, pierden a su vez los genes cry1C y cry1D (además del cry1Ab) (Figs. 1-4). Es importante mencionar que los plásmidos asociados con la actividad biopesticida de las cepas de Bt usualmen-te son plásmidos de tamaños superiores a 40 MDa (Wilks et al. 1998; Lereclus et al. 1989; Mahillon et al. 1988), lo que hace pensar que los genes cry1C y cry1D esta-rían ubicados en los plásmidos de 95 MDa y 99 MDa para la cepa nativa y de referen-cia respectivamente, aunque se deben rea-lizar trabajos posteriores para comprobar esta hipótesis.
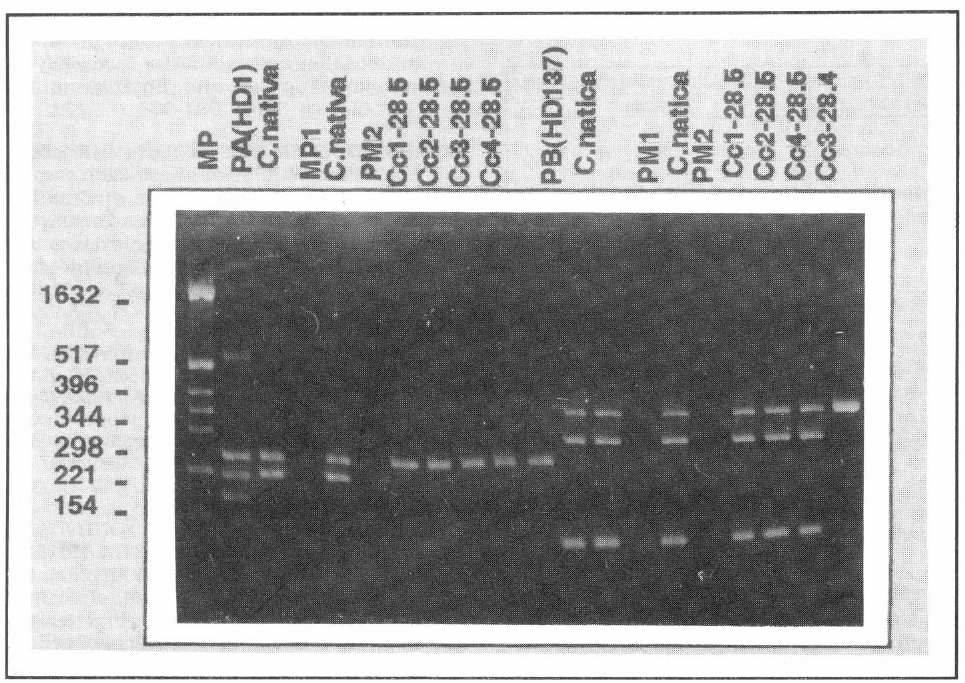
Análisis electroforético de los productos de amplificación de PCR para la cepa IBUN 28.5. MP: Marcador de peso molecular. PA(HD1): Patrón para la mezcla de reacción A, con primers específicos para los genes crylAa, cry1Ab y crylAc. C.nativa: Cepa nativa 28.5 sin tratar. PM1 y PM2: son muestras sin cargar pérdidas (no cargadas). Cc1-28.5 a Cc3-28.5 mismas colonias que en figura 1. Cc4-28.5 colonia que pierde el plásmido de 80 MDa. PB: patrón para la mezcla de reacción de B con primers específicos para los genes cry1B y cry1D.
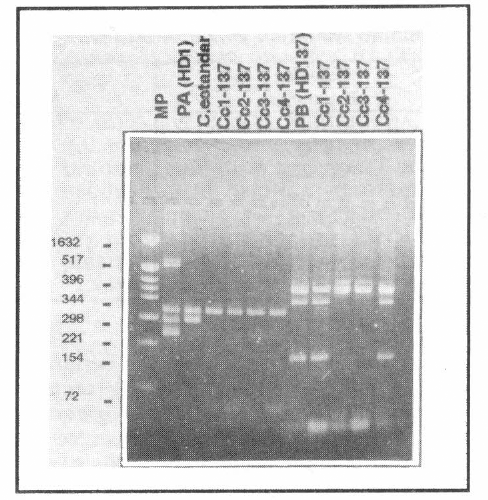
Análisis electroforético de los pro-ductos de amplificación de PCR para la cepa HD 137. MP: Marcador de peso molecular. PA(HD1): Patrón para la mezcla de reacción A, con primers específicos para los genes crylAa, cry1Ab y crylAc. C. estándar: Cepa de referencia HD 137 sin tratar. Cc1-137 y Cc2-137 mismas colonias que en figura 2. Cc3-137 у Сc4-137 colonia que pierden los plásmidos de 99 MDa, 77 MDa y 19 MDa, respectivamente. PB: patrón para la mezcla de reacción de B con primers específicos para los genes cry1B, cry1C y cry1D.
Análisis previos efectuados con otra cера de Btt subesp. aizawai, la Bt aisawai 7.29, sugieren que esta cepa posee una copia del gen cry1Ab en un plásmido de 45 MDa, y otra copia de este mismo gen a nivel cromosomal; dicha cepa posee ade-más otros tres genes crylAa, cry1C y crylD los cuales son de origen presu-miblemente cromosomal (Lereclus et al. 1989, Lereclus et al. 1993). La ubicación a nivel cromosomal y plásmidico de los genes cry en B. thuringiensis no es tema de discusión, aunque cabe mencionar que de acuerdo con lo señalado en la literatu-ra existe una distribución mayor de genes cry a nivel plásmidico que a nivel cromosomal (Ben-Dov et al. 1996; Lereclus et al. 1993; Lereclus et al. 1989; Donovan et al. 1988). Sin embargo, dada la impor-tancia de la cepa Bt aizawai HD137 como referencia por su actividad biopesticida contra el insecto blanco S. frugiperda (Ce-rón et al. 1994; Aranda et al. 1996; Bravo et al. 1998), cualquier caracterización de su genoma asociado a la actividad biopes-ticida de la cepa es de interés para el desa-rrollo de trabajos posteriores.
Bioensayos de mortalidad en larvas de primer ínstar de S. frugiperda enfrentadas a las cepas silvestres HD137 e IBUN28.5 y sus respectivas colonias curadas de plásmidos
Con él ánimo de darle un significado esta-dístico a los resultados de los bioensayos y de esta forma identificar diferencias en-tre los tratamientos, se llevó a cabo el aná-lisis de los datos de acuerdo con el criterio de la prueba de Duncan (Tabla 4). Para este análisis se tomaron en consideración cada una de las repeticiones realizadas en las dos dosis evaluadas, la prueba de Duncan agrupó las cepas bajo estudio como se indica en la tabla 4, confirmándo-se la superioridad en la actividad insectici-da sobre las larvas de S. frugiperda de la cepa nativa IBUN28.5 respecto a la cepa de referencia HD137. Así mismo, se observa una disminución en términos de la activi-dad biopesticida en las colonias que pier-den uno o más genes, lo que permite asumir que todos los genes cry curados (cry1Ab, cry1C y cry1D) tienen un efecto en la actividad biopesticida de ambas cepas.
Prueba de Duncan para el porcentaje de mortalidad de las diferentes colonias empleando un límite de confianza de 0.05
La relación directa entre la actividad biopesticida sobre S. frugiperda de los genes cry1C y cry1D no tienen discusión de acuerdo con diferentes anotaciones en este sentido en la literatura (Bravo et al. 1998; Smith et al. 1996; Aranda et al. 1996). Sin embargo, el efecto de la proteí-na Cry1Ab no es muy claro ya que de acuer-do con Aranda et al. (1996), dicha proteína únicamente se une de manera inespecífica a proteínas apicales de alto peso molecular (150 kDa) de las microvellosidades del in-testino medio, sin que esta unión esté re-lacionada con algún nivel de toxicidad en el hospedero.
En este contexto es posible que el efecto de la proteína Cry1Ab esté más relaciona-do con la posibilidad de desarrollar efec-tos sinérgicos entre proteínas Cry, como los descritos para las proteínas Cry1Aa y Cry1Ac contra Lymantria dispar, los cuales muestran que al estar juntas en una pro-porción 1:1 aumentan casi 4 veces más en relación con la actividad esperada debido a la acción de cada proteína por separado (Lee et al. 1996). Este tipo de efectos sinérgicos se pueden manifestar gracias a la formación de heterodímeros entre las toxinas, los cuales favorecen más estric-tamente la formación de poros en las cé-lulas del intestino medio de acuerdo con lo sugerido por Lee et al. (1996); y/o un incremento en la solubilidad de las otras proteínas que si son tóxicas, como sugie-ren Aronson et al. (1991), para el caso de la acción de la proteína Cry1Ab en la cера HD133. Se deben llevar a cabo estudios posteriores para dilucidar la importancia del gen cry1Ab en la actividad biopes-ticida de las cepas empleadas en este es-tudio contra larvas de S. frugiperda.
En la tabla 4 se observan diferencias en términos de la actividad biopesticida de la сера Сс1-28.5 y Cc2-28.5, a pesar de que estas cepas no presentan diferencias en su perfil plasmídico, ni de proteínas asocia-das al cristal, ni en el perfil de PCR. Este resultado sugiere que el proceso de cura-do de plásmidos posiblemente afectó los niveles de expresión de los genes cry que permanecieron en las cepas luego del pro-ceso de curado (crylAa, cry1B, cry1C o cry1D), o algún proceso involucrado en el mecanismo de infección de las mismas hacia el hospedero. Se deben realizar es-tudios posteriores para sacar afirmaciones más concluyentes. Cabe mencionar que gracias a que la mayoría de estudios coin-ciden en una pobre actividad biopesticida de la proteína Cry1Ab sobre S. frugiperda (Bravo et al. 1998, Aranda et al. 1996) el valor más cercano a la realidad debe ser el obtenido con la cepa Cc1-28.5.
La diferencia en términos de la actividad biopesticida entre las cepas silvestres de IBUN28.5 y HD137 y las colonias que pier-den el gen crylAb de ambas cepas (Cc1-28.5 y Cc1-137) se mantienen estables después del experimento de curado (de 12.2% y 9.4% respectivamente, Tabla 4). Sin embargo, entre las cepas que pierden los tres genes (cry1Ab, cry1C y cry1D) no se presentan diferencias en términos de la actividad biopesticida (Tabla 4).
Estos resultados permiten sugerir que las diferencias entre ambas cepas residen en características intrínsecas al nivel de se-cuencia de los genes cry1C o cry1D, que afecten su expresión y/o actividad biopes-ticida. Un hallazgo de esta naturaleza sería de gran interés por las posibilidades de corresponder a nuevos genes cry, o a dife-rencias en los niveles de expresión de es-tos dos genes, lo cual tendría implicaciones importantes para el desarrollo y manipu-lación de nuevos productos a base de Bt.
Conclusiones
Los resultados obtenidos en este estu-dio permiten sugerir que el SDS, la tem-peratura y/o el repique continuo de las cepas, deben tenerse en cuenta al mo-mento de diseñar un procedimiento para el curado de plásmidos en B. thuringiensis. En este contexto, las condiciones propues-tas para este diseño son: el empleo de SDS a una concentración entre 0.002 y 0.02% a una temperatura de incubación de 42°C ó la aplicación de subcultivos o repiques continuos cada 24 horas en agar nutritivo.
El plásmido de 80 MDa en la cepa nativa de Bt IBUN28.5 y el plásmido de 77 MDa de la cepa de referencia HD137 son porta-dores del gen cry1Ab.
Los genes cry1C y cry1D están asocia-dos a los plásmidos de 95 MDa y/o 17 MDa para la cepa nativa IBUN28.5 y a los plásmidos de 99 MDa y/o 19 MDa para la cepa de referencia HD137. Sin embargo, se propone que estos genes deben estar asociados al plásmido de mayor peso molecular (95 MDa y 99 MDa), por ser ésta una característica más generalizada en las cepas de Bt.
La mayor actividad biopesticida de la cepa nativa IBUN28.5, respecto a la cepa de referencia HD137, debe residir en caracterís-ticas intrínsecas de la secuencia de los genes cry1C y/o cry1D.
Footnotes
Agradecimientos
Esta investigación recibió apoyo de COLCIENCIAS y de la Universidad Nacional de Colombia, los autores desean agrade-cer a estas instituciones sin cuyo apoyo no hubiese podido lograrse esta investiga-ción.