
Abstract
A bioassay method was developed to determine in artificial diet the insecticidal activity of Bacillus thuringiensis native strains against first instar larvae of Spodoptera frugiperda. Bioassays were conducted at 25°C of temperature, 70% of relative humidity and a photoperiod of 12 hours-light. Native and reference strains were analized by PCR technology and SDS-PAGE in order to select strains containing cry I insecticidad crystal family genes with toxicity against Lepidopteran insects. Bacteria concen-trations were expressed as total protein and were incorporated in the surface of the diet. The mortality percentage was evaluated daily during 120 hours. In preliminary studies reference strains HD-1 var. kurstaki and HD-137 var. aizaway evaluated as a unique concentration of 150 µg of total protein/ml permited choose reference strain HD-137 as positive control for bioassays. Using this reference strain, native strains Bt-127, Bt-147, Bt-292, Bt-444 were evaluated. Native strain Bt-127 exhibited higher insecticidad activity than all other strains. This strain showed in first instar larvae 88% mortality, a lethal time 50 (LT50) of 24 hours and a lethal concentration 50 (LC50) of 83 µg/ml. The reference strain HD-137 showed an insecticidad activity significantly lower than native strain Bt-127; results were represented by a mortality percentage of 79%, a LT50 of 48 hours and a LC50 of 138 µg/ml. The insecticidal effect of the reference strain was followed by native strain Bt-147. Native strains Bt-292 and Bt-444 showed insignificant mortality percentages.
Introducción
Dentro del mercado mundial de bioinsecticidas la bacteria Bacillus thuringiensis ha sido utilizada en varias formulaciones comerciales como una alternativa para el control biológico de plagas agrícolas. Esta bacteria se caracteriza fundamentalmente por su habilidad para producir cristales de manera simultánea con la producción de esporas. Dichos cristales contienen potentes toxinas denominadas d-endotoxinas que son altamente específicas contra insectos de los órdenes Lepidoptera, Coleoptera y Diptera (Orduz 1992). Las d-endotoxinas son proteolíticamente activadas en el ambiente alcalino del intestino medio de las larvas de especies susceptibles, en donde liberan uno o más péptidos tóxicos que causan la lísis de las células epiteliales, septicemia y la posterior muerte (Bravo y Quintero 1993).
A nivel nacional el gusano cogollero, Spodoptera frugiperda, es una de las plagas más importantes debido a los daños que ocasiona en cultivos de importancia económica tales como algodón, maíz y sorgo. Durante 1990 se destinaron para el control de este insecto 5.8 millones de dolares en cultivos de algodón y 4.2 millones de dolares en cultivos de maíz y sorgo, con los cuales se trataron 430.000 ha y 440.000 ha respectivamente. (Murillo 1991).
El control químico de este insecto, mediante el uso indiscriminado de insecticidas, ha ocasionado altos costos económicos y ambientales, por lo cual el control biológico aparece como una alternativa promisoria, siendo Bacillus thuringiensis el microorganismo que mayor potencial ha demostrado para su uso en condiciones prácticas de campo. Dada la gran biodiversidad existente en Colombia, es importante la búsqueda y selección de nuevas cepas de este microorganismo que presenten alta especificidad contra plagas locales y un amplio espectro de acción. Por lo tanto, el objetivo del presente trabajo fue el de generar una metodología para la evaluación de la actividad insecticida de cepas nativas de Bacillus thuringiensis contra larvas de primer instar de Spodoptera frugiperda bajo condiciones de laboratorio.
Materiales y métodos
La presente investigación se llevó a cabo en los laboratorios del Programa Nacional de Manejo Integrado de Plagas de Corpoica, ubicados en el Centro de Investigaciones "Tibaitatá" Regional 1. (Mosquera-Cundinamarca). El pie de cría de S. frugiperda provino de material recolectado en estado de pupas a partir de un cultivo de maíz ubicado en el Centro de Investigaciones La Libertad, Regional 8 de Corpoica (Villavicencio-Meta). La cría fue renovada cada cuatro meses a partir de material proveniente de campo y las condiciones ambientales para su mantenimiento en laboratorio fueron: 25°C de temperatura, fotoperíodo de 12 horas luz y una humedad relativa de 70%.
Cría de Spodoptera frugiperda
Los insectos durante todo su estado larval fueron mantenidos dentro de vasos plásticos de 14 ml de capacidad, los cuales contenían previamente una dieta artificial estandarizada por López (1981); cuando las larvas alcanzaron el estado de pupa, se sexaron y se colocaron parejas de S. frugiperda en relación de cuatro hembras y tres machos por cada porrón de vidrio de boca ancha (1 galón de capacidad), que contenía en la base papel absorbente. El frasco poseía en su interior cuatro tiras de papel para recibir la oviposición de los adultos y era tapado con una tela (velo) sujetada con una banda de caucho, siguiendo la comunicación personal de Espitia (1995). Para la alimentación de los adultos emergidos se colocó una solución azucarada al 20% en vasos plásticos de 14 ml de capacidad que contenían algodón en el fondo del recipiente.
Las tiras de papel en las cuales los adultos ovipositaban se colocaron en frascos que contenían la dieta artificial. Aproximadadente a los dos ó tres días se presentaba la eclosión de los huevos y cuando las larvas alcanzaban su segundo o tercer ínstar, se depositaban individualmente en un vaso plástico transparente de 14 ml de capacidad, con 2 cm de diámetro y 2.5 cm de profundidad que contenía previamente 7-9 gramos aproximadamente de dieta fresca; de esta manera se evitaba el canibalismo propio de la especie. Cada vaso era tapado con un cartón estéril e incubado durante 23 días hasta cuando el insecto llegara de nuevo al estado de pupa.
Selección de las cepas de Bacillus thuringiensis para bioensayos
Las cepas utilizadas en el presente trabajo provenían del banco de cepas del laboratorio de Control Biológico de Corpoica. Estas fueron seleccionadas con base en los siguientes criterios: procedencia de los aislamientos nativos con orígenes geográficos diferentes, que presentaran por caracterización microscópica abundante esporulación y producción de cristales, que en su caracterización bioquímica mediante la técnica de la SDS-PAGE y molecular mediante la técnica de la reacción en cadena de la polimerasa (PCR) demostraran la presencia de los genes cry IC y cry ID los cuales codifican para proteínas con actividad contra Spodoptera spp. (Tabla 1). Esta caracterización bioquímica y molecular de las cepas fue realizada por el Programa Nacional de Biotecnología Agrícola de Corpoica.
Bioensayos
Para cada uno de los ensayos biológicos correspondientes a:
Selección de una cepa de referencia
Evaluación de la actividad insecticida de las cepas nativas y
Determinación de la concentración letal media (C150) de las cepas, de referencia y nativas seleccionadas.
Se utilizaron las condiciones metodológicas descritas a continuación:
Preparación del inóculo microbial para las cepas nativas y de referencia
Del contenido microbial de un cultivo de Bacillus thuringiensis crecido durante veinte días a 28° C en agar L, se preparó una suspensión de la bacteria en 10 ml de tampón fosfato a pH 7.4. A partir de esta suspensión se realizaron diluciones seriadas en el mismo tampón y se determinó la concentración de proteína total (esporas y cristales) de la bacteria, a una longitud de onda de 595 nm mediante la técnica espectrofotométrica de Bradford (1976). Para ello, se utilizó una curva de calibración con albúmina sérica bovina, la cual permitió correlacionar las lecturas de absorbancia obtenidas a 595 nm para las distintas concentraciones de albúmina, con respecto a las concentraciones de proteína total de cada una de las cepas evaluadas. La ecuación de predicción de la regresión lineal por el método de mínimos cuadrados corresponde a:
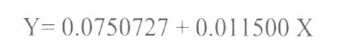
A partir del tubo que contenía la concentración deseada de la bacteria, expresada como microgramos de proteína total /ml en forma estéril, se tomaron 200 µl y se inocularon completamente sobre la superficie de la dieta contenida en cada vaso. Este inóculo se dejó secar durante dos horas en ambiente estéril para luego infestar cada vaso con una larva de primer instar de S. frugiperda y taparlo con un cartón estéril. El material biológico provino de la cría mantenida en condiciones de laboratorio.
Diseño experimental
Las condiciones de evaluación para cada uno de los ensayos biológicos fueron de 25°C de temperatura, 70% de humedad relativa y un fotoperíodo de 12 horas-luz. Las dietas se inocularon con la concentración respectiva para cada ensayo y se colocó una larva de primer instar por vaso, para un total de 20 larvas por cada tratamiento (20 unidades experimentales).
Para cada ensayo, se incluyó: un tratamiento correspondiente a un control tratado con la solución tampón y sin el inóculo bacteriano (testigo tratado), un tratamiento control sin la solución tampón y sin el inóculo bacteriano (testigo absoluto) y un tratamiento con la concentración correspondiente de la cepa de referencia, suspendida en la solución tampón (control positivo).
El diseño experimental para la realización de cada uno de los ensayos biológicos fue completamente aleatorizado; cada ensayo fue repetido en tres días diferentes y las lecturas de mortalidad, teniendo en cuenta el número de larvas vivas y muertas, se realizaron cada 24 horas durante un total de 120 horas. Para determinar el porcentaje de mortalidad corregido en cada ensayo biológico se utilizó la fórmula de Abbott (Ciba-Geigy 1978).
Cepas de Bacillus thuringiensis seleccionadas y caracterizadas por microscopia, PCR y SDS-PAGE
Análisis de resultados
Para las interacciones de la variable mortalidad, evaluada entre los diferentes tratamientos en cada uno de los ensayos biológicos por separado con S. frugiperda, se realizó una prueba de comparación de promedios Duncan con un nivel a del 0.05% de significancia. Para la deteminación de los valores de la concentración letal media (C150) se utilizó el análisis estadístico de regresión logarítmico Probit de POLO-PC (LeOra Software 1987).
Descripción de los diferentes ensayos biológicos
Selección de una cepa de referencia de B. thuringiensis como control positivo
Se evaluaron las cepas de referencia HD-1 var. Kurstaki y HD-137 var. aizaway para seleccionar aquella que sirviera como control positivo en los posteriores bioensayos. Siguiendo las condiciones metodológicas anteriormente descritas, estas cepas fueron cultivadas y para cada una de ellas se obtuvieron dos concentraciones de proteína total correspondientes a 50 µg/ml y 150 µg/ml, las cuales fueron inoculadas en las dietas para luego realizar el diseño experimental y la evaluación del bioensayo.
Evaluación preliminar de la actividad insecticida de las cepas nativas
Según las condiciones metodológicas descritas anteriormente, se montaron los bioensayos utilizando la cepa HD-137 como control positivo y las cepas nativas Bt-127, Bt-147, Bt-292, Bt-444. A partir del contenido microbial de cada una de estas cepas se ajustó una suspensión bacteriana expresada para cada una de ellas en una concentración única de proteína total correspondiente a 200 µg/ml. Se inocularon 200 µl sobre la superficie de cada dieta dispensada por vaso, en la que posteriormente se infestó cada vaso con dieta, con una larva de primer ínstar, y finalmente se realizó el diseño experimental previamente descrito.
Determinación de la concentración letal media (C150) de las cepas seleccionadas
A partir de un cultivo de la cepa HD-137 y de la cepa nativa Bt-127 se realizaron cinco diluciones bacterianas, las cuales fueron expresadas como concentración de proteína total/ml. Estas concentraciones para cada una de las cepas correspondieron a 50, 100, 150, 200 у 250 µg/ml de proteína total. Cada concentración se utilizó para inocular las dietas, en las cuales posteriormente se colocó una larva de primer instar por vaso con dieta. Las condiciones metodológicas y el diseño experimental fueron similares al descrito inicialmente.
Resultados y discusión
Selección de una cepa de referencia como control positivo
El análisis de Duncan no mostró diferencias significativas en la actividad insecticida de las cepas HD-1 y HD-137 contra larvas de primer instar de S. frugiperda. Sin embargo, se observó una tendencia ligeramente mayor de la cepa HD-137 para controlar las larvas del insecto. Este análisis mostró diferencias significativas entre los tratamientos en los que se utilizaron concentraciones de 150 µg/ml de proteína total con aquellos en los que se utilizó una concentración de 50 µg/ml de proteína total y con los dos controles (tratado y absoluto) (Fig. 1).
Se estableció para las dos cepas una relación directamente proporcional entre la concentración de la bacteria inoculada a la dieta y la actividad insecticida. El mayor porcentaje de mortalidad acumulada durante 120 horas obtenido al aplicar en la dieta 150 µg/ml de proteína total fue de 50% para la cepa HD-137 con respecto a un 44% para la cepa HD-1, mientras que al aplicar en la dieta 50 µg/ml de proteína total se obtuvo un 21% de mortalidad para la cepa HD-137 con respecto a un 19% de mortalidad para la cepa HD-1. Este comportamiento fue similar al registrado por Hofte et al. (1988) y Van Rie et al. (1990), quienes encontraron que la cepa HD-137 con respecto a la cepa HD-1 presentaba una mayor actividad insecticida contra larvas de primer instar de Spodoptera littoralis. Este mismo resultado fue registrado por Ceron et al. (1994) para el caso de larvas de primer instar de S. frugiperda. En este último ensayo se registró una concentración letal media (C150) de 969 ng de proteína de cristal/cmβ para la cepa HD-1 y una concentración letal media de 43 ng/cmβ para la cера HD-137.
Las diferencias en los rangos de mortalidad hallados en la presente investigación para las dos cepas pudieron deberse a la mayor susceptibilidad de las larvas de S. frugiperda a las proteínas codificadas por los genes cry IAa, IB, IC, ID de la cepa HD-137, mientras que la cepa HD-1 presentó los genes cry IAa, IAb, IAC, IIA, IIB, V, los cuales podrían codificar para proteínas menos activas contra el insecto. Dada la tendencia mostrada por la cepa HD-137 respecto al porcentaje de mortalidad acumulado producido, ésta fue escogida como control positivo para los siguientes bioensayos.
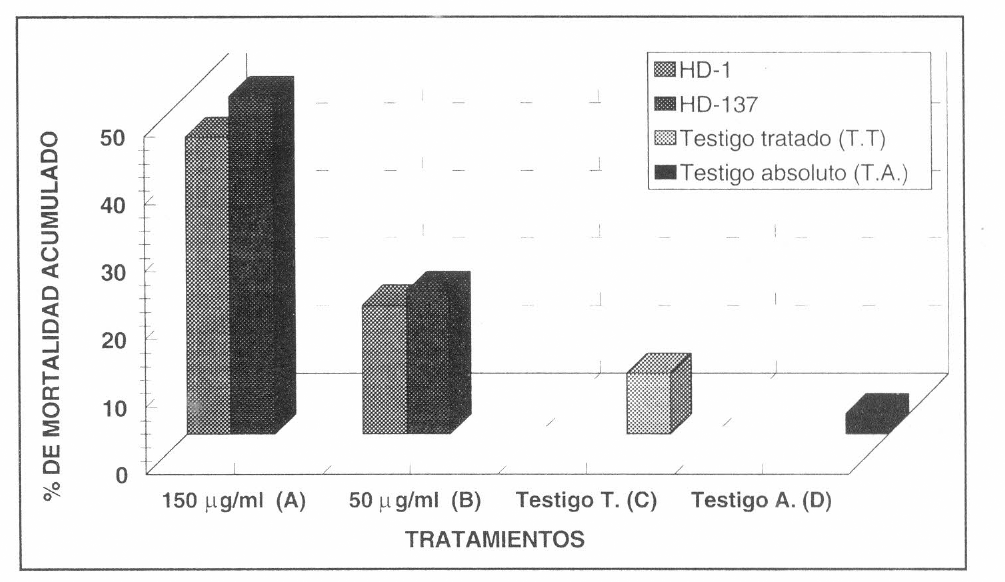
Evaluación de la actividad insecticida de dos cepas de referencia de B. thuringiensis contra larvas de primer instar de S. frugiperda durante 120 horas. Las barras que comparten la misma letra no son significativamente diferentes según la prueba Duncan con un a del 0.05% de significancia.
Actividad insecticida de las cepas nativas
Para la evaluación preliminar de la actividad insecticida de las cepas nativas de B. thuringiensis se utilizó la cepa HD-137 como control positivo y cuatro cepas nativas según los criterios de selección de las cepas anteriormente mencionados; todas estas cepas se evaluaron a una concentración única correspondiente a 200 µg/ml de proteína total. Los resultados de este bioensayo se expresaron como porcentajes diarios de mortalidad después de expuestas las larvas a la dieta que había sido inoculada con las diferentes cepas.
Se observó durante un tiempo de 120 horas de estar expuestas las larvas a la bacteria, que la mortalidad fue máxima y permaneció estable. El análisis estadístico de Duncan determinó diferencias significativas en los porcentajes de mortalidad producidos por las cepas HD-137 y Bt-127 con respecto a las demás cepas evaluadas (Fig. 2).
Los análisis de mortalidad observados mostraron que dentro de las cepas nativas, la cepa Bt-127 fue la que presentó la mayor actividad insecticida. Según el análisis de Duncan, esta actividad fue significativamente diferente a la de las demás cepas evaluadas (Fig. 2). La cepa Bt-127 presentó un tiempo letal medio (T150) de aproximadamente 24 horas en el experimento y un porcentaje acumulado de mortalidad de 88% a las 120 horas. El tiempo letal medio (T150) para las otras cepas fue de 48 horas para la cера HD-137 у de 72 horas para la cepa Bt-147. Las cepas Bt-292 y Bt-444 no alcanzaron un T150 al termino de la evaluación. De otra parte, los porcentajes acumulados de mortalidad fueron de 79% para la cepa HD-137, de 65% para la cepa Bt-147, de 43% para la cepa Bt-292 y de 33% para la cepa Bt-444. Se observó que los parámetros de mortalidad y de T150 de la cepa Bt-127 no fueron superados por los correspondientes a las demás cepas (Fig. 3).
La alta actividad insecticida de las cepas HD-137 y Bt-127 podría estar relacionada con la expresión de los genes IAa, IB, IC, ID y IAa, IC, ID, respectivamente. Algunos autores registraron que varias especies de Spodoptera, tales como, Spodoptera exigua y Spodoptera exempta eran susceptibles a las toxinas codificadas por los genes cry IC y cry IE (Visser et al. 1990; Bai et al. 1993). De otra parte, Van Rie et al. (1990) encontraron una elevada actividad insecticida contra Spodoptera exigua y una baja actividad contra Spodoptera exempta en cepas que poseían los genes cry ID. Esto permitiría sugerir que diferentes genes podrían codificar toxinas con actividad contra varias especies de este insecto.
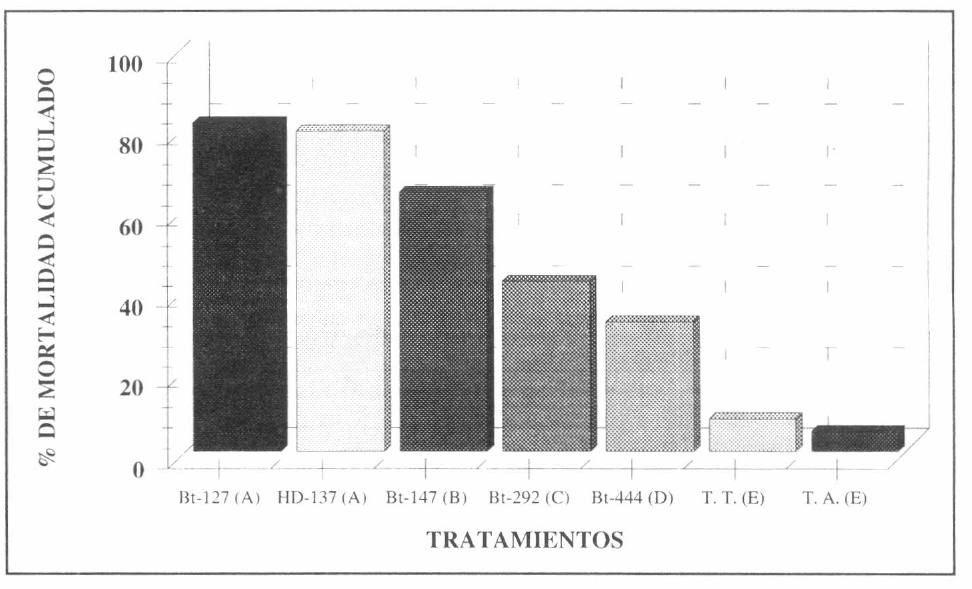
Evaluación de la actividad insecticida de las cepas nativas de B. thuringiensis contra larvas de primer instar de S. frugiperda durante 120 horas. Las barras que comparten la misma letra no son significativamente diferentes según la prueba Duncan con un nivel a del 0.05% de significancia.
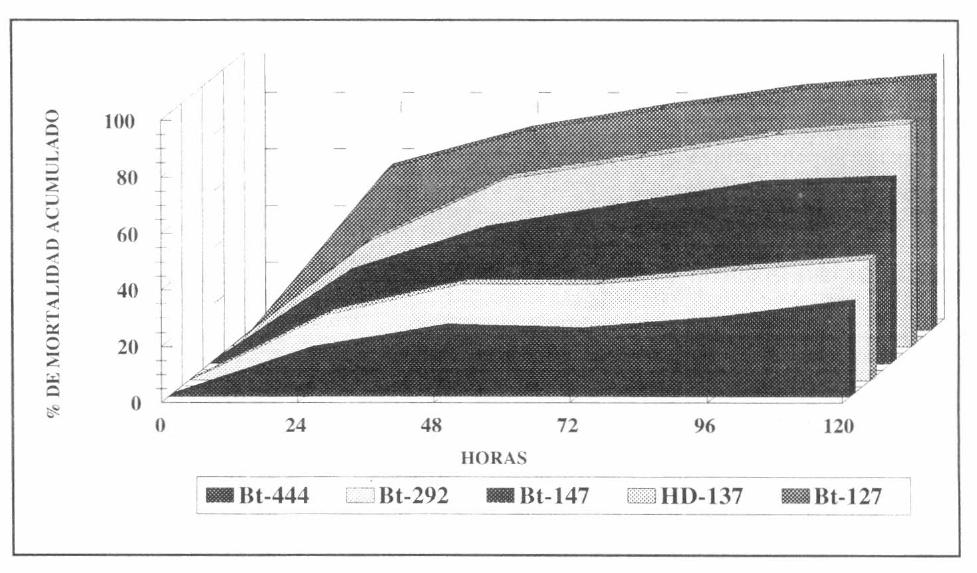
Evaluación de la actividad insecticida de las cepas nativas de B. thuringiensis contra larvas de primer instar de S. frugiperda.
La actividad insecticida alta de estas dos cepas también podría estar relacionada con una mayor afinidad de sus d-endotoxinas con los receptores intestinales de las larvas de S. frugiperda. Sin embargo, si se analiza el hecho que a pesar de que la cepa Bt-444 presenta los mismos genes de la cepa HD-137, mostró una actividad significativamente menor de 33%, con respecto a un 79% de la cepa HD-137; se podría pensar que aunque estas cepas comparten los mismos genes, presentan niveles de expresión de d-endotoxina diferentes. De otra parte, es probable que aquellas cepas nativas que presentaron una actividad menor produjeron toxinas menos activas contra S. frugiperda, ó produjeron menores cantidades de d-endotoxina. Esta escasa actividad tóxica podría ser debida igualmente según Sanchis y Ellar (1993) a que los cristales tóxicos que las cepas nativas Bt-147, Bt-292 y Bt-444 produjeron, no encontraron receptores en la membrana del intestino medio de las larvas a los cuales se pudieran acoplar, ó a que los cristales se desnaturalizaron en el intestino larval. En todos los casos, tanto con la cepa de referencia como con las diferentes cepas nativas, se observó inhibición en el crecimiento de las larvas sobrevivientes. Este fenómeno es, según Murillo (1987), debido a que las subdosis consumidas por las larvas prolongan su ciclo de vida y producen un retardo en el metabolismo larval.
Las cepas HD-137 y Bt-127 fueron escogidas como las más activas contra S. frugiperda basados en los mayores porcentajes de mortalidad producidos en el menor tiempo.
Concentración letal media (C150) de las cepas seleccionadas
Se determinó la C150 de las dos cepas (Bt-127 y HD-137) probando cinco diluciones sucesivas expresadas como proteína total. Las concentraciones probadas fueron 50, 100, 150, 200 y 250 µg/ml.
El análisis estadístico Probit mostró, a las 120 horas de iniciado el bioensayo, una C150 menor para la cepa Bt-127 que la obtenida para la cepa HD-137, las cuales fueron de 83.51 µg/ml y de 138.65 µg/ml respectivamente. Se evidenció una relación directamente proporcional entre las concentraciones utilizadas en las dos cepas evaluadas y la mortalidad causada por ellas (Tabla 2).
Pérez y Rodríguez (1996) establecieron la relación entre la concentración letal y el porcentaje de mortalidad en diferentes especies de insectos Lepidópteros como Heliothis virescens (Fabricius) y Tecia solanivora (Povolny). Se podría asumir que al aumentar la concentración de proteína, la dosis ingerida por cada larva también aumenta, expresándose en una mayor mortalidad.
Se registró una tendencia mayor en los porcentajes de mortalidad obtenida durante 120 horas, tanto en las concentraciones bajas de la cepa nativa como en las altas, con respecto a los porcentajes de mortalidad producidos por la cepa HD-137, estos fueron de 25% y 98% de mortalidad para la cepa Bt-127 y de 14% y 81% de mortalidad para la cepa HD-137 en sus concentraciones baja y alta respectivamente. Estos resultados confirman la promisoria actividad insecticida de esta cepa nativa contra S. frugiperda.
Previo a la muerte de las larvas tratadas con las cepas Bt-127 y HD-137, se observaron signos similares de inapetencia, reducción de tamaño, flacidez, pérdida de movilidad y aparición de coloraciones oscuras debido a los tejidos necrosados por la invasión bacteriana. Estas observaciones coincidieron con lo descrito por Pérez y Rodríguez (1996).
Concentraciones letales 10, 50 y 90 para las cepas HD-137 y Bt-127
Concentración letal determinada por análisis Probit con un 95% de confiabilidad.
Conclusiones
La concentración de la bacteria expresada en microgramos de proteína total/ml permitió evidenciar el efecto insecticida de las cepas evaluadas.
La cepa de referencia HD-137 var. aizaway fue seleccionada por su actividad insecticida como control positivo en los bioensayos.
Dentro de las cepas nativas evaluadas se seleccionó como promisoria la cepa Bt-127 por su elevada actividad insecticida contra larvas de primer instar de S. frugiperda.
Recomendaciones
Evaluar la actividad insecticida de la cepa Bt-127 en ensayos de campo.
Seleccionar en laboratorio nuevas cepas nativas de Bacillus thuringiensis con una elevada actividad insecticida y un alto espectro de acción contra otras plagas locales.
Realizar estudios para la producción masiva y formulación de las cepas promisorias de Bacillus thuringiensis para su aplicación en campo.