
Abstract
Some aspects of acoustical communication of eight species of Tettigoniidae from Ucumarí Regional Natural Park (RNP) are presented for the first time. These species belong to three subfamilies: Pseudophyllinae (5 species): Trichottettix pilosula, Stetharasa exarmata, Chibchella nigrospecula, Scopioricus spatulatus, Typophyllum sp.; Copiphorinae (1 species): an unnamed genus and species, and Listroscelidinae (2 species): an unnamed genus and species. A brief diagnostic description and general ecological aspects of each species are provided. The physical structure of the five species found in Ucumarí is varied. The songs of the katydids described here are noisy, i.e. with many frequencies. The highest frequency encountered in this group was 17.5 kHz, produced by T. pilosula, and the lowest was 8 kHz, produced by Scopioricus spatulatus. Only one species presented energy in the ultrasonic range, and three species produced a band spectrum: Chibchella nigrospecula, Stetharasa exarmata and Scopioricus spatulatus; all of the others tend toward high-Q tones.
Introducción
La familia Tettigoniidae está conformada por insectos orthopteroides conocidos comúnmente con el nombre de saltamontes o chapulines. Representan un grupo bastante diverso, con cerca de 6300 especies descritas (Naskrecki y Otte 1999). Estos insectos muestran relación con los grillos, aunque al parecer los mecanismos de generación de canto en estos dos grupos han evolucionado en forma diferente (Gwynne 1995). Sin embargo, se ha establecido que el taxón hermano de la familia Tettigoniidae es la familia Haglidae (Naskrecki 2000).
Como en otros grupos de insectos, los saltamontes machos producen señales acústicas que tienen función tanto en la formación de parejas como en la expresión de rivalidad entre machos. Estas señales acústicas son generadas únicamente por adultos y se producen por frotación de una serie de dientes ubicados en el ala anterior izquierda (fila o vena estriduladora), sobre un borde esclerotizado (raspador) ubicado en el área anal del ala anterior derecha. En algunas especies (subfamilia Phaneropterinae), la hembra puede producir señales acústicas en respuesta al llamado del macho, pero el órgano generador es diferente al del macho (Nickle 1976).
Las señales sonoras han sido usadas en estudios de bioacústica enfocados a la formación de parejas (Morris 1980), fisiología (Bailey 1995) y en taxonomía (Walker y Gurney 1972; Morris y Walker 1976; Morris y Beier 1982; Walker y Greefield 1983; Morris et al. 1989). En muchos casos estos estudios bioacústicos han ayudado a la separación de nuevos taxa, sobre todo en especies hermanas (Walker et al. 1973). Otro interés en el estudio de estas señales sonoras está en investigar las causas de producción de ultrasonidos y los mecanismos de recepción que muchas especies han desarrollado (Belwood y Morris 1987; Mason et al. 1991; Morris et al. 1994).
Se pueden encontrar especies semiápteras, ápteras, braquípteras y macrópteras. En especies semiápteras los machos alcanzan a tener unas tegminas (alas anteriores) muy cortas, las cuales alcanzan únicamente el tamaño del campo estridulador y normalmente están cubiertas por el pronotum, de tal forma que a simple vista no se ven. Sin embargo, en las hembras de estas especies, casi siempre las tegminas están ausentes. Hay pocas especies ápteras en las cuales ambos sexos carecen de tegminas, de tal forma que los machos no poseen órgano estridulador y por lo tanto son completamente "mudos". La mayoría de especies de Tettigoniidae son nocturnas, permanecen ocultas durante el día en troncos de árboles caídos, cortezas, o en el follaje y en la noche salen a realizar sus actividades como oviposición, cópula y alimentación (Nickle 1992; Montealegre 1997).
Los Tettigoniidae son muy diversos en las regiones tropicales, especialmente en Colombia donde muchas especies son aún desconocidas. Solo existen algunos trabajos antiguos que fueron realizados en su mayoría con material colectado en la región andina (Hebard 1927, 1933); otros trabajos son los de Nickle (1985), Montealegre et al. (1993), Montealegre y González (1995) y Montealegre (1996). En ellos se describen aspectos taxonómicos de una gran cantidad de especies de Colombia, principalmente de la región andina y del Chocó biogeográfico (especialmente del Valle del Cauca) y se registra la existencia de aproximadamente 250 especies en todo Colombia. Actualmente, se tiene un buen conocimiento de la fauna del departamento del Valle del Cauca, en donde se han registrado más de 130 especies (Montealegre 1997; Montealegre y Morris 1999). Las especies señaladas para Colombia pertenecen a 6 subfamilias: Phaneropterinae, Pseudophyllinae, Copiphorinae, Agraeciinae, Conocephalinae y Listroscelidinae.
A pesar que la mayoría del material examinado en los trabajos de Hebard (op. cit.) provenía de diferentes departamentos de la región andina, no se conocía hasta el presente nada de la fauna del departamento de Risaralda (Montealegre 1997). El primer estudio realizado en este departamento comenzó con una corta visita al Parque Regional Natural Ucumarí en el mes de mayo de 1996, en donde se notó la existencia de posibles nuevos taxa; además se obtuvieron grabaciones del canto de algunas especies. Un año después y en la misma época, se realizaron búsquedas sistemáticas intensas en la noche por los diferentes senderos del parque con el fin de conseguir más información.
Materiales y Métodos
Área de estudio
El Parque Regional Natural (PRN) Ucumarí se encuentra en el departamento de Risaralda a unos 28 km al este del municipio de Pereira, entre las coordenadas planas (3° W de Bogotá):

El parque se encuentra entre los 1850 y 2600 m de elevación. Limita por el norte y por el oeste con el Parque Nacional Natural Los Nevados, por el sur con la divisoria de aguas de los ríos Otún y Quindío y por el oeste con la quebrada Paloblanco hasta su confluencia con el río Otún. La precipitación varía con el gradiente altitudinal entre 2000 y 4000 mm/año y de igual forma la temperatura entre 12 y 18°C. En esta área se encuentra representada la zona de vida Bosque muy Húmedo Montano (datos tomados de Londoño 1994).
Se incluyen adicionalmente algunos datos de distribución y de campo para algunas especies que se encuentran también en la Reserva Natural La Sirena. Esta reserva se encuentra ubicada en el departamento del Valle del Cauca y pertenece al municipio de Palmira. La Sirena es un bosque premontano de la cordillera central y se encuentra en el área de protección de la cuenca del río Nima (la principal fuente de agua del municipio de Palmira, entre los 3° 32′ N, 76° 7' E). Su rango altitudinal va desde los 2600-4100 m y su precipitación varía entre 1600-2800 mm/año.
Trabajo de campo
Los especímenes se colectaron en la noche con la ayuda de lámparas de cabeza por los senderos de ambas reservas. Se realizaron dos visitas al Parque Ucumarí entre los meses de abril y mayo de 1996 y 1997; los especímenes se colectaron por senderos cercanos al refugio turístico La Pastora. El trabajo de campo en la Reserva Natural La Sirena (Palmira) comprendió dos visitas entre los meses de febrero y abril de 1999.
El canto de algunos especímenes se grabó directamente en el campo con equipo limitado al rango audible; otros fueron transportados vivos para ser grabados en el laboratorio. Las grabaciones del campo se realizaron con una grabadora Sony WM D6C Professional y un micrófono ECM 909. Varias veces se usó el "zero-crossing meter" de un detector de murciélagos S-25 para localizar y grabar cantos. La temperatura se tomó con un termómetro digital Omega HH23.
Trabajo de laboratorio
El equipo usado en el laboratorio fue exitoso en la grabación de cantos, tanto ultrasónicos como en el rango audible, desde 1 a 100 kHz. Para esto se empleó equipo Brüel & Kjaer. Se conectó un micrófono condensador 1/4 pulgadas (4135) a un amplificador o a un medidor de presión de sonido. La producción total de cualquiera de estos amplificadores se registró en una grabadora de cinta Racal de 4 canales a una velocidad de 30 pulgadas/s. Posteriormente, las señales fueron digitalizadas usando una tarjeta de sonidos Keithley DAS50 y analizados con software DADISP.
Una vez se transportaron los insectos al laboratorio, los machos se encerraron individualmente en jaulas pequeñas elaboradas con tela metálica, la cual facilita la grabación de sus cantos. El micrófono condensador se puso a una distancia aproximada de 10 cm del insecto enjaulado.
Taxonomía
Las descripciones morfológicas de Tettigoniidae se encuentran en Naskrecki (1996) y Montealegre (1996) entre otros. Cuando se dan los tamaños corporales para algunas especies, éstos son expresados en milímetros (mm) y los insectos medidos longitudinalmente desde el fastigium del vértice de la cabeza hasta el décimo tergito.
La determinación taxonómica de los especímenes se realizó por comparación con referencias publicadas y con material existente en museos, especialmente del Museo de Zoología de la Universidad de Michigan (MZUM) y el Museo Naturhistoka Riksmuseet, Stockholm (MNRS). Se contó también con la opinión de otros expertos: Dr. Piotr Naskrecki, de la Universidad de Connecticut y el Dr. Theodore Cohn, MZUM.
Los nuevos taxa, pertenecientes a la subfamilia Pseudophyllinae, han sido recientemente descritos (Montealegre y Morris 1999), mientras que la descripción de las especies nuevas de otras subfamilias (Copiphorinae y Listroscelidinae) se encuentra en progreso.
Terminología
La siguiente terminología es usada para describir las señales acústicas (según Morris y Walker 1976; Morris et al. 1989; Morris 1999).
Longitud de onda (λ). En toda ondulación periódica, la perturbación se repite a intervalos iguales de distancia λ, que corresponden a la llamada longitud de onda. La longitud de onda de una ondulación sinusoidal es la distancia entre dos crestas adyacentes.
Amplitud de onda (A). Es la altura de una cresta o la profundidad de un valle, medidas a partir de la posición de equilibrio de la onda.
Periodo (T). Es el tiempo para un ciclo completo de una oscilación u otro movimiento regularmente repetido.
Frecuencias de las ondas acústicas. Hay vibraciones mecánicas de frecuencias muy variadas. La unidad de frecuencia es el hertz (Hz), pero cuando se manejan frecuencias altas es común expresar la frecuencia en múltiplos de mil (kHz). El oído humano solo percibe vibraciones cuyas frecuencias se encuentran entre 20 hertz y 20 x 103 hertz (20 kHz). Las oscilaciones cuyas frecuencias se encuentran entre estos límites se denominan sonidos, pero la palabra acústica es usada por los físicos para referirse también a las ondulaciones mecánicas cuyas frecuencias se encuentran fuera del espectro audible. Si la frecuencia es superior a la sonora, las ondulaciones se llaman ultrasónicas.
Intensidad. La intensidad del sonido se refiere a la sensación de fuerza o volumen, aunque esto depende también de la frecuencia. Es inversamente proporcional al cuadrado de la distancia de la fuente. El nivel de intensidad de un sonido es la intensidad respecto de un patrón convenido. La unidad fundamental es el bel, pero generalmente se usa el decibel (dB) (1 dB = 0.1 bel). El umbral de la audición se toma como 0 dB en la medida del sonido. Diez veces este nivel de potencia es 10 dB.
Armónico. Los sobretonos cuyas frecuencias son múltiplos enteros de la frecuencia fundamental se denominan armónicos. La frecuencia fundamental se denomina primer armónico, el primer sobretono, el segundo armónico y así sucesivamente.
Llamada (=canto). Es el patrón de amplitud de tiempo más repetitivo durante la emisión del canto del insecto.
Periodo de llamado (o de canto). Es el intervalo de tiempo entre dos cantos sucesivos que incorpora el intervalo del silencio entre cantos (Fig. 1).
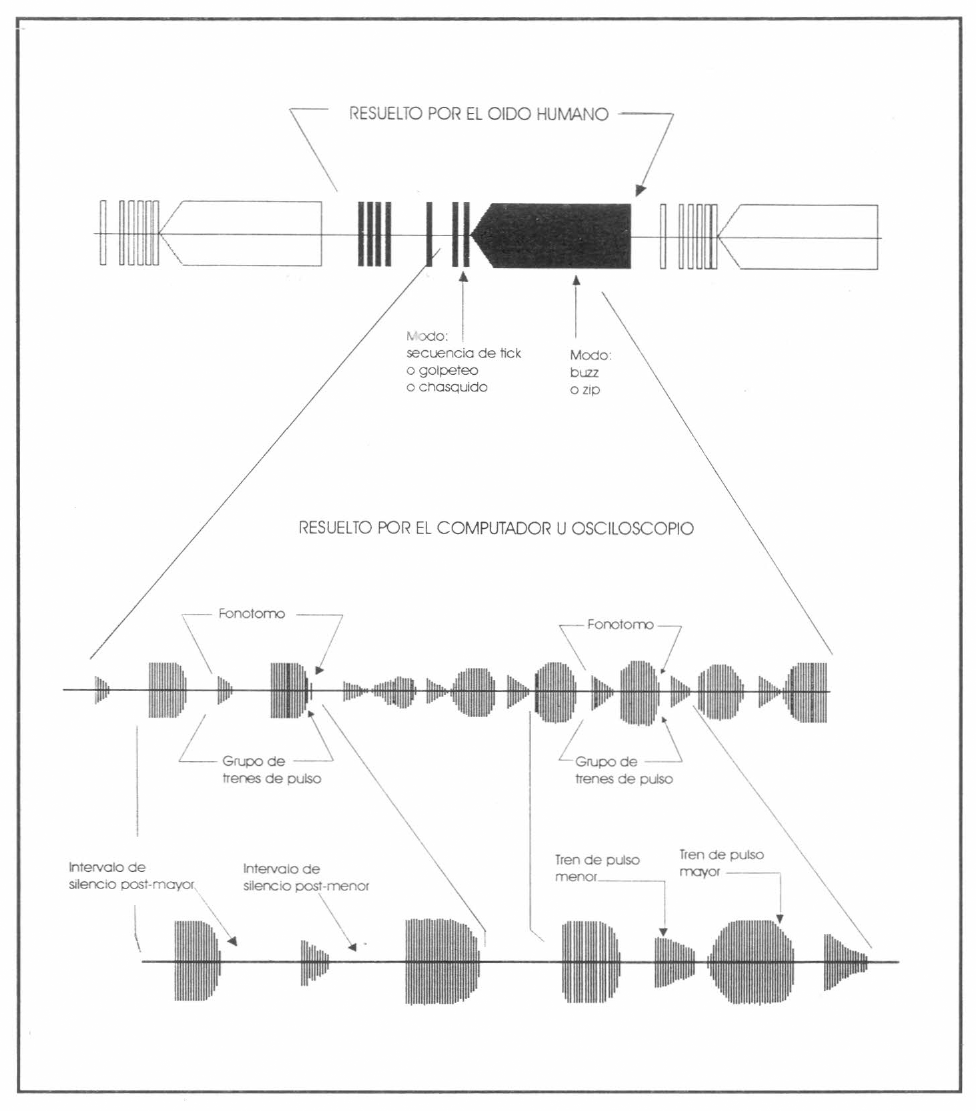
Representación hipotética de un sonograma para explicar la terminología empleada en el texto (modificado de Morris y Walker 1976; Morris et al. 1989).
Duración del ciclo. Es la proporción del periodo de canto gastado emitiendo sonido.
Pulso. Un tren de onda aislado en tiempo por una modulación de amplitud (AM) que declina a nivel de ruido del fondo.
Periodo de pulso. Es el espacio de tiempo entre un pulso y el pulso que le sigue.
Pulso de decadencia rápida. Un tren de onda caracterizado por rondas transitorias, una forma de onda compleja y decadencia rápida después de uno o pocos ciclos de máxima amplitud.
Pulso sinuosoidal sostenido. Un tren de onda caracterizado por rondas graduales (en vez de rondas transitorias) y una forma de onda sinuosoidal. sostenido por varios o muchos ciclos en su más alta amplitud o cerca de ella.
Tren de pulso (TP). Un primer orden de agrupamiento de dos o más pulsos, generalmente con alguna regularidad y su velocidad de repetición precedida y seguida por un periodo de silencio substancialmente más grande en duración que cualquiera de los intervalos de tiempo entre los pulsos (Fig.1).
Grupo de trenes de pulsos (GTP). Un segundo orden de agrupamiento de pulsos. Una secuencia de dos o más trenes de pulsos mutuamente distintos. TPs y GTPs nunca son idénticos y es materia de opinión y sentido común de cuán diferentes pueden ser éstos antes que sean considerados distintos (Fig. 1).
Tren de pulso mayor (TPMa). Es el TP con más duración y con mayor intensidad dentro de un GTP (Fig. 1).
Tren de pulso menor (TPMe). Es el TP con duración más corta e intensidad menor dentro de un GTP (Fig. 1). Los pulsos de un TPMe por lo general se traslapan mientras que los de un TPMa son discretos y separados por silencios. La velocidad de los pulsos de un TPMe es siempre notablemente más alta que la velocidad de los pulsos de su TPMa asociado.
Interva'o de silencio post-mayor (ISPMa). Es el intervalo de silencio que sigue después de un TPMa (Fig. 1).
Intervalo de silencio post-menor (ISPMe). Es el intervalo de silencio que sigue después de un TPMe (Fig. 1).
Fonótomo. El término fonótomo fue usado por (Leroy 1966), y puede definirse como todo el canto producido durante un ciclo del movimiento de las tegminas. Un fonótomo es un componente del sonido definido con base en el comportamiento de las tegminas. Como tal, puede ser difícil de aplicar cuando la base del movimIen to generador del sonido no se conoce con exactitud. La velocidad de la estridulación impide por lo general. asociar el movimiento generador a los elementos de señales por simple observación. Por lo tanto en ausencia de técnicas especiales, es posible predecir el fonótomo de una especie en particular. Sin embargo muchas especies alternan dos tipos de pulso o trenes de pulso, uno corresponde presumiblemente al movimiento del raspador en la clausura (o cierre) y el otro en la apertura de las tegminas. AsÍ, el termino fonótomo es usado aquí como una probable unidad repetitiva, aun cuando la correspondencia de los elementos sonoros al movimiento generador sea desconocida.
Q (el factor de calidad). El término Q describe las propiedades de resonancia de un sistema, es decir es una medida del rango de frecuencias en que un sistema opera exitosamente (Bennet-Clark 1999; Prestwich et al. 2000). Cuando se tienen valores Q grandes (en comparación a valores previamente conocidos), es un indicio que el sistema está sintonizado (hecho) para resonar simultáneamente con el mecanismo que lo estimula. En el caso de los saltamontes, el mecanismo implica un plectro o raspador que es frotado sobre una hilera de dientes. En especies con factor Q alto, cada vez que el raspador contacta un diente se produce una vibración de la membrana (o membranas) amplificadoras del espejo. O sea que la velocidad de contacto del raspador sobre los dientes se convierte en función de la frecuencia generada (a la cual la membrana amplificadora vibra) y la relación se dice que es 1: 1; de tal forma que se produce un ciclo (u onda) por cada diente tocado y antes que esta onda sonora decaiga, se produce el siguiente contacto y en general el ciclo se sostiene. Esto indica que mediante un oscilograma podemos predecir el número de dientes que el raspador contacta para generar el sonido. El espectro producido por este tipo de mecanismo posee una sola frecuencia y en acústica se conoce como tono puro (también musical de alto Q o resonante). En especies en donde la membrana amplificadora genera diferentes vibraciones (frecuencias) por cada diente tocado, no existe una relación 1:1 en el sistema, las oscilaciones producidas van a decaer antes de que el raspador contacte el próximo diente de la fila y el sonido va a presentar un espectro de banda ancha. Este tipo de sonido se conoce como ruidoso, no resonante o de Q bajo.
El factor Q se puede calcular de varias formas, pero una forma sencilla es usando un oscilograma y midiendo la amplitud de cada ciclo en la decadencia del pulso (no en el incremento [Bennet-Clark 1999]). Se calcula el valor absoluto de la pendiente de la recta que resulta de graficar las ondas con el número asignado (desde la onda cero a la onda n, esto depende de la cantidad de ondas de interés) en el eje X contra el logaritmo natural de las amplitudes para cada una de esas ondas en el eje Y. Mediante la ecuación:
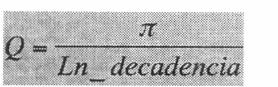
se calcula el valor. Normalmente Q superiores a 15 se consideran altos (Prestwich et al. 2000; Montealegre y Morris 2001).
Muchos saltamontes y grillos mueven sus tegminas tan rápidamente (para estridular) en un movimiento de "ida y regreso" (clausura o apertura) que se requiere de técnicas muy sofisticadas de fotografía para saber si un determinado TP coincide con la apertura o clausura de las tegminas durante la estridulación. Se ha encontrado que los TPMa coinciden con la clausura de la tegmina mientras que los TPMe coinciden con la apertura (Thomas y Alexander 1962). Sin excepción, los componentes del canto más intensos y más prolongados son producidos en la clausura o cierre de las tegminas.
Algunos componentes del canto de ciertos saltamontes pueden ser percibidos por los humanos. Es conveniente usar términos descriptivos basados sólo en la percepción de cantos que el oído humano puede efectuar. El componente de canto temporal más corto que puede ser resuelto completamente por un receptor humano podría componerse de otros sonidos más cortos. Aunque nuestro oído no puede separar claramente esta infraestructura, podemos percibir su presencia como la impresión que da un sonido "particular" o "serrado". La tabla 1 resume la definición de ocho componentes de sonido diferentes de acuerdo a si el componente dura más de un segundo o menos, si el componente es ruidoso (si tiene muchas frecuencias portadoras diferentes) o si es musical dominado por una sola frecuencia y si su calidad indica una infraestructura.
Terminología usada para describir los componentes del canto con base en la percepción del oído humano (las palabras empleadas para describir cada sonido se presentan con su equivalente en inglés entre paréntesis). La duración se expresa en segundos (s). Modificado de Morris et al. 1989
Zumbidos (buzz), golpeteos y secuencias de tictac (tick) (Fig. 1). Las distinciones de estos términos se basan en las tasas de repetición de fonótomos. Los zumbidos son el resultado de las tasas altas de fonótomos; el fonótomo individual no puede ser resuelto por nuestro oído y el sonido no da la impresión de tener TPs y GTPs (Fig. 1). Si la velocidad del fonótomo es muy baja, haciendo posible contar y distinguir fonótomos individuales, el componente del sonido se define como una secuencia de tictac. En fonótomos de velocidades intermedias se puede detectar como un zumbido con calidad tremulante o secuencia de tictacs en las cuales las unidades ocurren muy rápidamente para ser contadas; en tal caso se describe el componente de canto como un golpeteo. Se debe tener presente el efecto substancial de la temperatura sobre la velocidad de fonótomos; el mismo componente de canto identificado como un golpeteo a una temperatura alta puede sonar como una secuencia de tictac a una temperatura más baja.
Otros términos útiles referentes a la resolución del oído humano son silbido (zip) y chasquido (smack). Un silbido es un zumbido corto con menos de 1 segundo en duración, el cual tolera un incremento notable en la amplitud desde el primer tercio hasta un medio de su producción. Un chasquido es un tictac particularmente ruidoso y cargado de sonido (Fig. 1).
Resultados
La fauna de Tettigoniidae del PRN Ucumarí comprende hasta el presente ocho especies en las subfamilias Pseudophyllinae (5 especies), Copiphorinae (1 especie) y Listroscelidinae (2 especies). A continuación se presenta una breve descripción morfológica (basada en la descripción original) y algunos aspectos generales de su biología acompañados por una amplia descripción de los componentes acústicos de cada una. Algunos géneros y especies no han sido descritos todavía y por ello sólo se presenta una diagnosis general.
Subfamilia Pseudophyllinae
Trichotettix pilosula Stål 1873
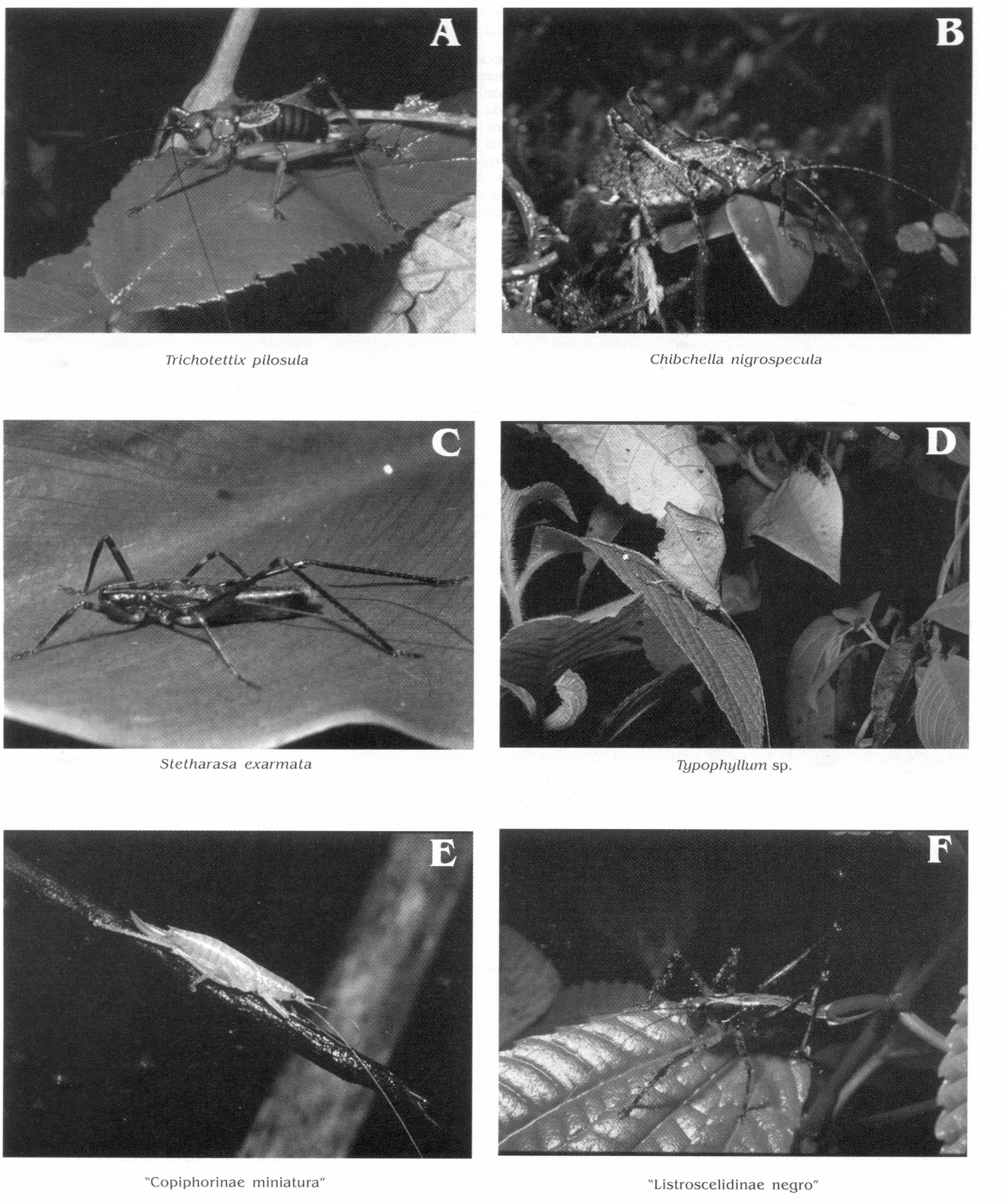
Algunas especies de Tettigoniidae del parque. A. Trichotettix pilosula (hembra), B. Chibchella nigrospecula (macho), C. Stetharasa exarmata (macho), D. Typophyllum sp. (macho), E. "Copiphorinae miniatura" (hembra), F. "Listroscelidinae negro" (hembra). B, C, E, cortesía de F. Vargas; A, D, F, fotografías de G.K. Morris.
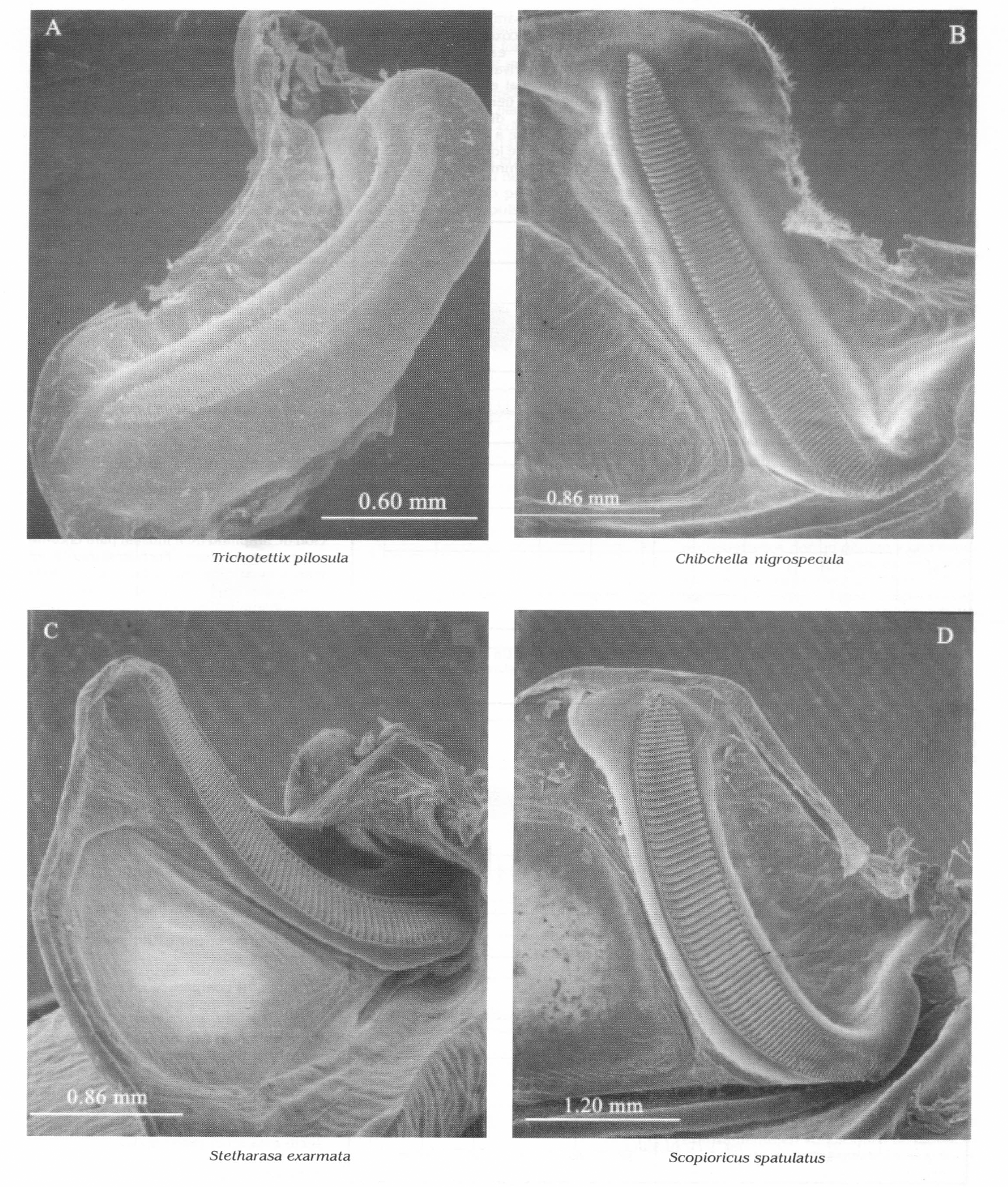
Detalle de la fila estriduladora de algunas especies mostrando forma y número de dientes. A. Trichotettix pilosula, B. Chibchella nigrospecula, C. Stetharasa exarmata, D. Scopioricus spatulatus. Después de Montealegre y Morris 1999.
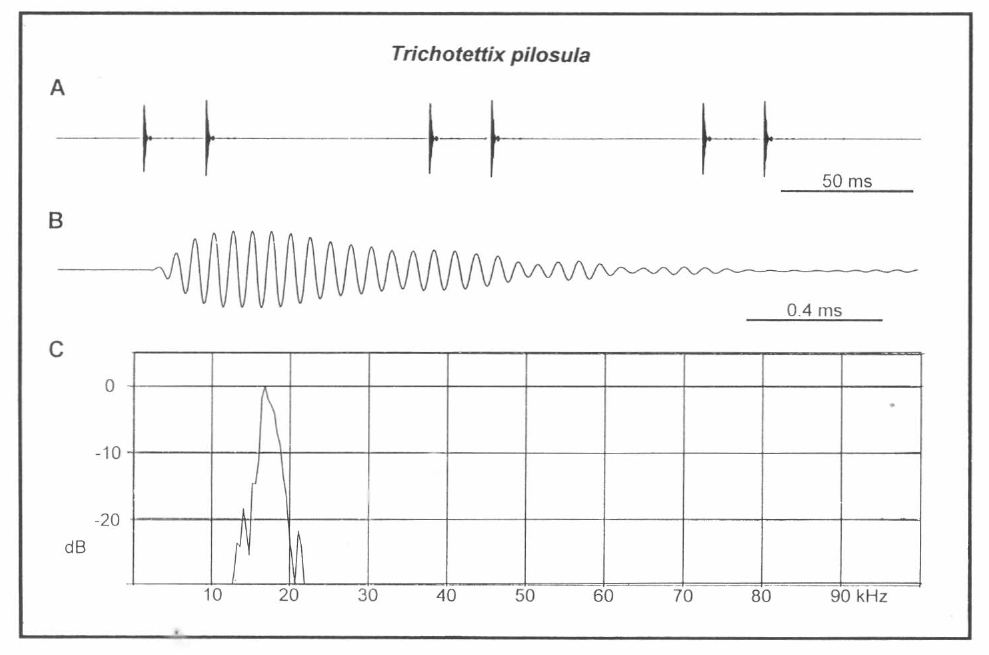
Análisis del canto de T. pilosula. A. Tres llamadas, B. Un simple pulso a alta resolución, C. Espectro de potencia del pulso visto en B.
La amplitud de un pulso se incrementa rápidamente para formar entre 7 y 14 ondas intensas, con poca o ninguna altiplanicie y luego existe alguna variación de amplitud en larga decadencia (Fig. 4B). Posiblemente cada par de pulsos constituye un fonótomo, es decir, los dos corresponden a un ciclo del movimiento de las tegminas. Pero esto es difícil de considerar cuando la fila estriduladora tiene un gran número de dientes (110, n=1) (Fig. 3A). Alternativamente se pueden producir varios pares de pulsos durante un recorrido de la fila estriduladora.
El espectro está dominado por un simple pico moderadamente estrecho, cercano a los 17 kHz (Fig. 4C). Se encuentran también armónicos débiles, pero estos están 20 dB por debajo del tope de energía del pico principal. Uno de los machos produjo su máximo pico a 17.5 kHz (rango 17.0-17.7). El otro macho produjo picos levemente más bajos, a 16.3 kHz (rango 16.3-16.4). El pico principal medio obtenido para el segundo macho en condiciones naturales fue de 16.3 kHz: exactamente el mismo conseguido a 11 y 21°C.
Chibchella nigrospecula Montealegre & Morris 1999
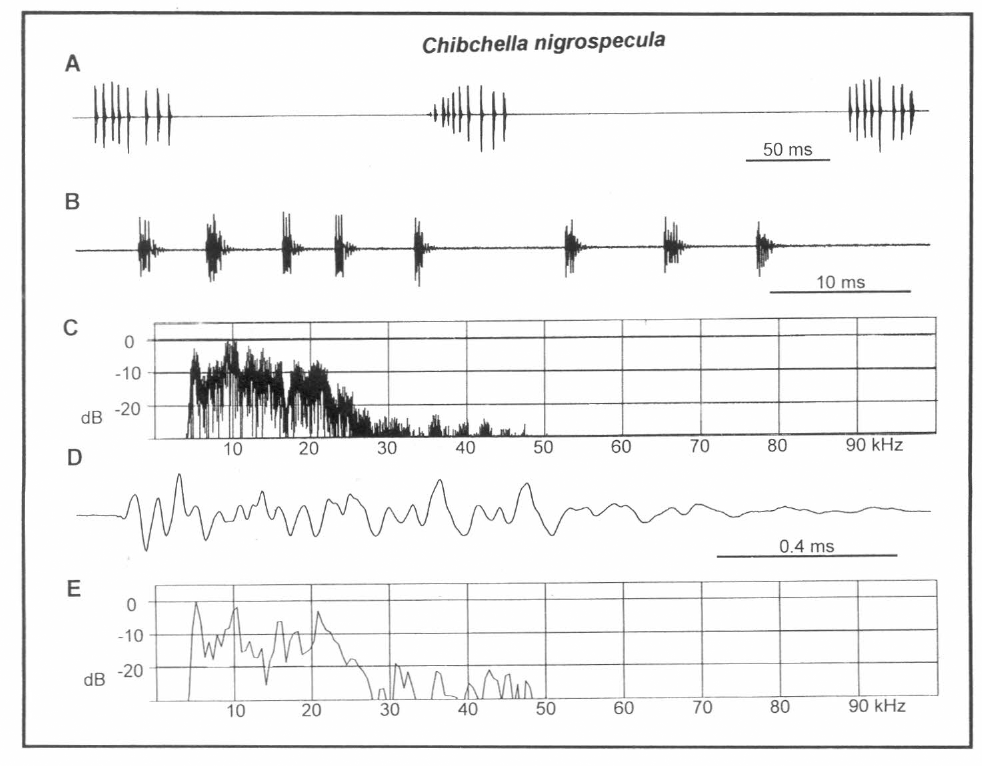
Análisis del canto de Chibchella nigrospecula. A. Una llamada compuesta de tres zips, B. Un zip a alta resolución, C. Espectro de potencia de B., D. Forma de onda compleja de un pulso simple de B., E. Espectro de potencia de D.
El canto es fácilmente audible y de acuerdo con esto, la mayoría de la energía espectral se encuentra en el rango audible de los humanos (Fig. 5C). La forma de la onda de pulso (Fig. 5D) es compleja, creando una banda de espectro que va desde 5 a 25 kHz (Fig. 5E). No parece haber algún pico predominante, aunque hay subpicos a 5 y 10 kHz.
Este insecto se observó produciendo también series de llamadas vibratorias mientras se encontraba enjaulado en el laboratorio. Las series de llamadas vibratorias eran alternadas con llamadas acústicas.
Stetharasa exarmata Montealegre & Morris 1999
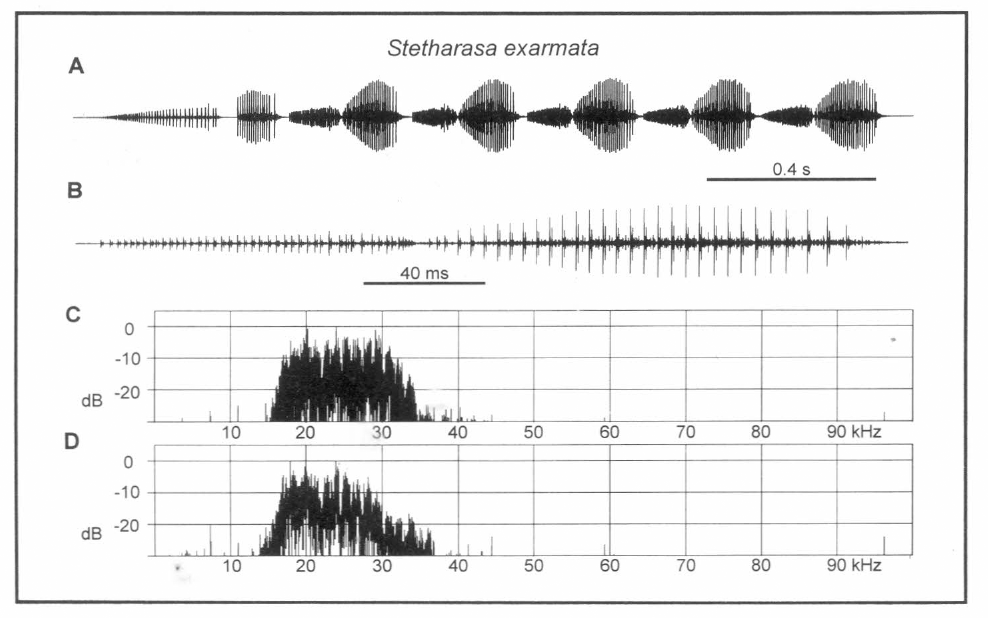
Análisis del canto de Stetharasa sp. A. Porción de secuencia, B. Un fonótomo, C. Espectro de potencia de A., D. Espectro de potencia de B.
Cada fonótomo (Fig. 6B) se compone de un tren de más baja amplitud y de pulsos de decadencia rápida (TPMe) combinado con un tren mayor más intenso (TPMa). El espectro resultante del TPMe contiene una banda coherente de frecuencias transportadoras entre 16 y 34 kHz (Fig. 6C). La energía está distribuida más bien uniformemente sobre esta banda de tal forma que no hay algún pico que identifique la frecuencia y no hay una diferencia apreciable entre el espectro de los dos tipos de trenes (TPMe y TPMa).
El periodo de fonótomo de esta especie es 272 ms. La velocidad del fonótomo a una temperatura de 20°C fue 3.2 por segundo. Hay 42 pulsos en el tren menor producidos a una velocidad de 420 por segundo, mientras en el tren mayor hay 32 pulsos que ocurren a una velocidad de 220 por segundo (Fig. 6B). Es notable en esta especie la coherencia inusual de los pulsos del tren menor (TPMe) y su larga duración, casi 100 ms. Los dientes de la fila estriduladora son conspicuamente uniformes en anchura y espacio (Fig. 3C). Posiblemente la morfología de esta fila está relacionada con la separación de pulsos más clara en tiempo. La duración del tren es muy larga comparada con otros Tettigoniidae que no producen resonancia, como Conocephalus spp., por ejemplo.
Scopioricus spatulatus Montealegre & Morris 1999
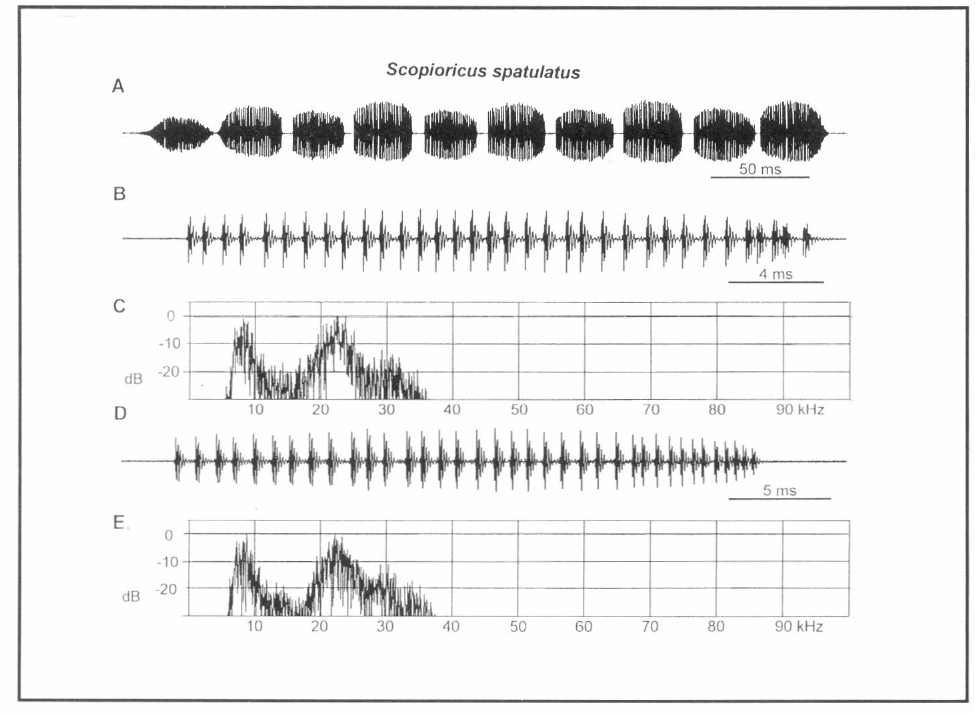
Análisis de canto de Scopioricus sp. A. Una llamada, B. Tren de pulso del movimiento de ida, C. Espectro de potencia del tren en B., D. Tren de pulso del movimiento de venida, E. Espectro de potencia del tren en D.
La igualdad aproximada en amplitud y duración de dos trenes de pulso sucesivamente distintos es un patrón de modulación de amplitud (AM) inusual para el canto de un saltamontes. Frecuentemente un tren con intensidad notablemente más corta y más baja, será producido entre trenes con intensidad más larga y más intensa. Pero aquí los dos tipos son casi iguales (Fig. 7A). El receptor recibe la impresión que el emisor cambia de orientación con cada secuencia sucesiva, tal como ocurre con el sonido hacia adelante y hacia atrás (ida y venida) producidos cuando se asierra madera con un serrucho. Los despliegues de la tegmina durante el silbido pueden ser seguidos a simple vista y de esta forma se pudo confirmar la existencia de golpes efectivos con sonidos aproximadamente iguales en direcciones opuestas. Hay alguna similitud en la estructura de la fila estriduladora de esta especie y la de Stetharasa exarmata, también habitante de esta zona (comparar Figs. 3C y 3D). Sin embargo los dientes de la fila estriduladora de S. spatulatus están separados entre sí a una distancia de casi el doble de lo que están los de la fila estriduladora de St. exarmata.
La figura 7B comprende un tren hecho en una dirección (apertura) a alta resolución mientras la figura 7D es un tren hecho en dirección opuesta (clausura), los espectros de cada uno son idénticos (Fig. 7C, E). Hay dos picos en un espectro de banda; uno es de casi 8 kHz y el otro, aproximadamente a la misma intensidad, es casi de 23 kHz. El volumen de energía espectral de este insecto está en el rango audible.
Typlophyllum sp
El género Typophyllum pertenece a la tribu Pterochrozini en la cual se agrupan especies que mimetizan hojas secas (Fig. 2D). Se encontró una sola especie perteneciente a este género pero la sistemática de este grupo es complicada y aún no se tiene certeza de su identidad.
Esta especie exhibe un despliegue aposemático inusual que consiste en la exposición de manchas que simulan ojos en la base de los miembros. La membrana trocanto-coxal de los dos pares de patas anteriores es de color naranja fluorescente, bordeado por pigmentación negra que contrasta en la parte distal de la cоха у proximal del trocánter. La extensión de fémur y trocánter relativa a la coxa expone esta membrana (de forma semilunar) dando la apariencia de un par de "ojos".
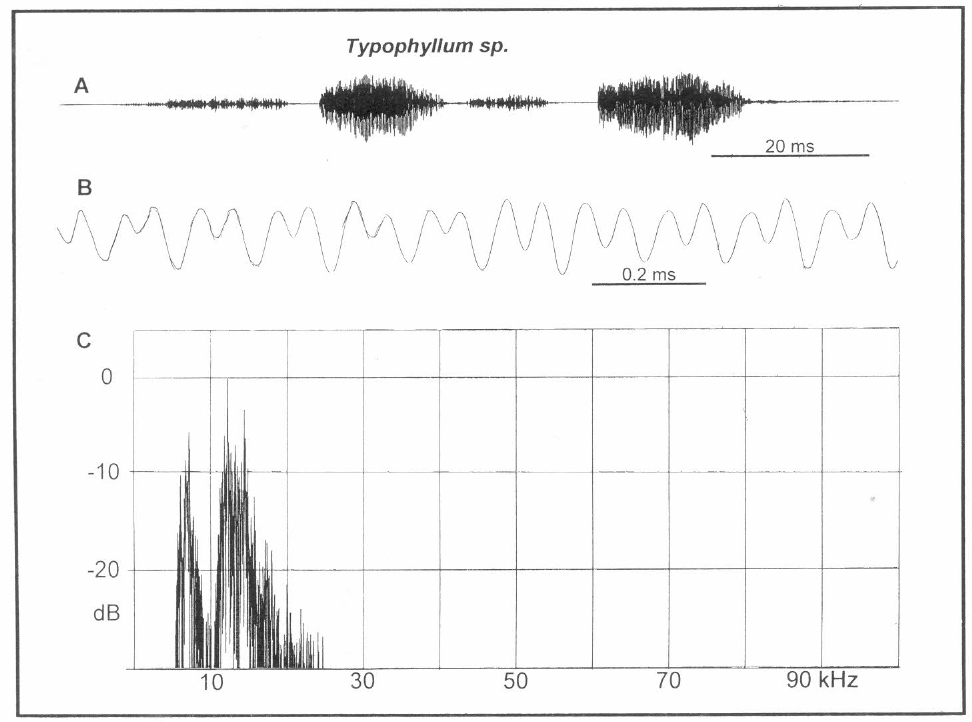
Análisis de canto de Typophylum sp. A. Una llamada compuesta de dos fonótomos, B. Onda compleja del pulso mayor en A., C. Espectro de potencia de A.
El canto es fácilmente percibido por el oído humano, ubicándose casi completamente en el rango audible (Fig. 8C). Hay dos picos espectrales de intensidad comparable armónicamente relacionados. El más intenso es casi 12.1 kHz mientras su fundamental aparente, 6.8 kHz, está levemente suprimido (valor medio para n=13 llamadas, FFT calculada sobre el total de cada canto). La forma de onda (Fig. 8B) muestra el patrón característico de dos frecuencias transportadoras dominantes relacionadas armónicamente. En vista de contar con un solo espécimen no fue posible disecar la vena estriduladura para los estudios detallados de ésta.
Copiphorinae
Los Copiphorinae del PRN Ucumarí están representados por un género y una especie. Este género (de especies pequeñas) es nuevo para la ciencia (Morris y Montealegre en prepar.). Algunos especímenes se encontraron perchados en hojas de arbustos a unos pocos metros por encima del suelo cerca al campamento en PRN Ucumarí. En un comienzo se creyó que esta especie pertenecía a un género descubierto recientemente en Costa Rica (Naskrecki, com. pers.), el cual parece estar relacionado con el género Eriolus Bolívar (Bolívar 1888) pero presenta más similitud con el género Borinquenula (Walker y Gurney 1972). Estudios más detallados en sistemática y acústica, referentes a esta especie, se encuentran en progreso; mientras tanto, a esta especie se le da el nombre asignado en el campo "Copiphorinae miniatura".
"Copiphorinae miniatura"
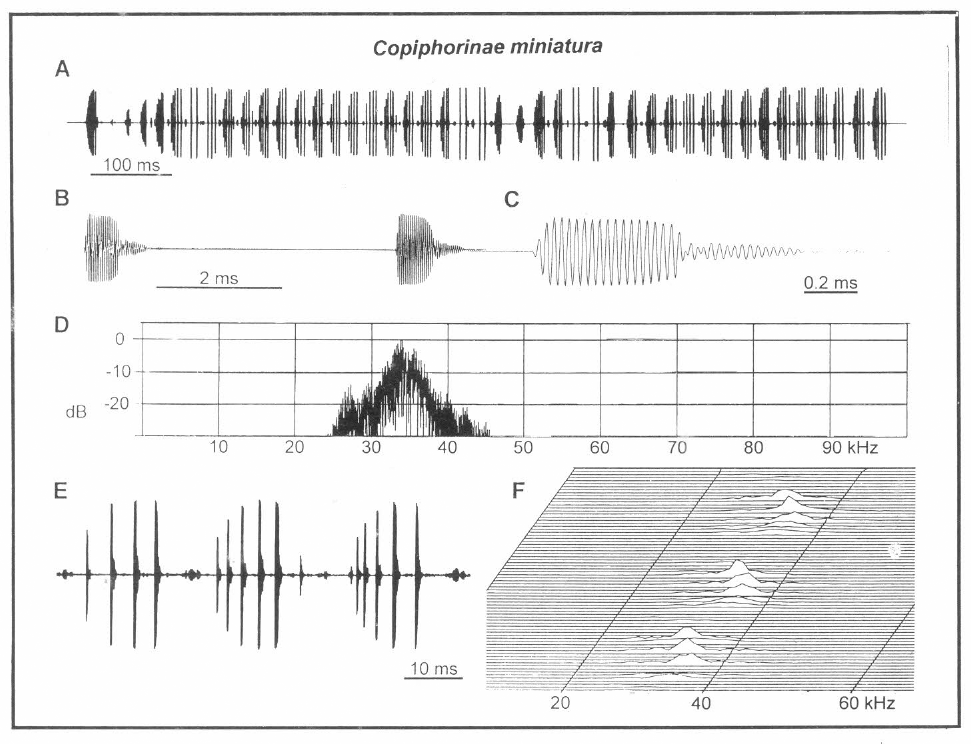
Análisis de canto de "Copiphorinae miniatura". A. Una llamada (canto), B. Dos pulsos sucesivos, C. Ondas de sonido de un pulso, D. Espectro de uno de los pulsos visto en B., E. Tres fonótomos, F. Diagrama en tres dimensiones mostrando el espectro de un fonótomo.
Listroscelidinae
Los Listroscelidinae conforman un grupo pequeño pero de amplia distribución mundial. Este grupo está representado en Ucumarí por un género y dos especies (referenciadas aquí como "Listroscelidinae ámbar" y "Listroscelidinae negro"). El género no ha sido descrito, la descripción oficial se encuentra actualmente en progreso (Montealegre y Morris, en preparación).
"Listroscelidinae ámbar"
"Listroscelidinae negro"
Diagnóstico. Son insectos pequeños (machos 19, hembras 20 mm) de color marrón oscuro, casi negro. Machos braquípteros y hembras ápteras. La coloración de las hembras es más oscura que la de los machos, pero ambos sexos poseen una franja dorsal de color crema (Fig. 2F).
La figura 10A muestra los tres fonótomos promedio de una ronda. Cada uno consiste en un sonido precedente de baja amplitud y un sonido más intenso y sostenido. Una característica de este canto es la completa ausencia de una estructura de pulsos de decadencia rápida discreta dentro de la emisión de alta amplitud (Fig. 10B); no hay evidencia de interacciones diente-raspador. Este componente intenso es, hasta ahora un simple pulso de 68 ms de duración, con una forma de onda completa y extremadamente variable (Fig. 10C). Su cubierta de amplitud es más bien errática y variable, pero es consistente en forma sobre sucesivos pulsos del macho. Un sonido de baja amplitud que precede por casi 35 ms, está presente en los dos fonótomos finales de la figura 10A, pero efectivamente ausente del primero. En sus rondas de tictacs, este macho regularmente omitió este componente de baja amplitud.
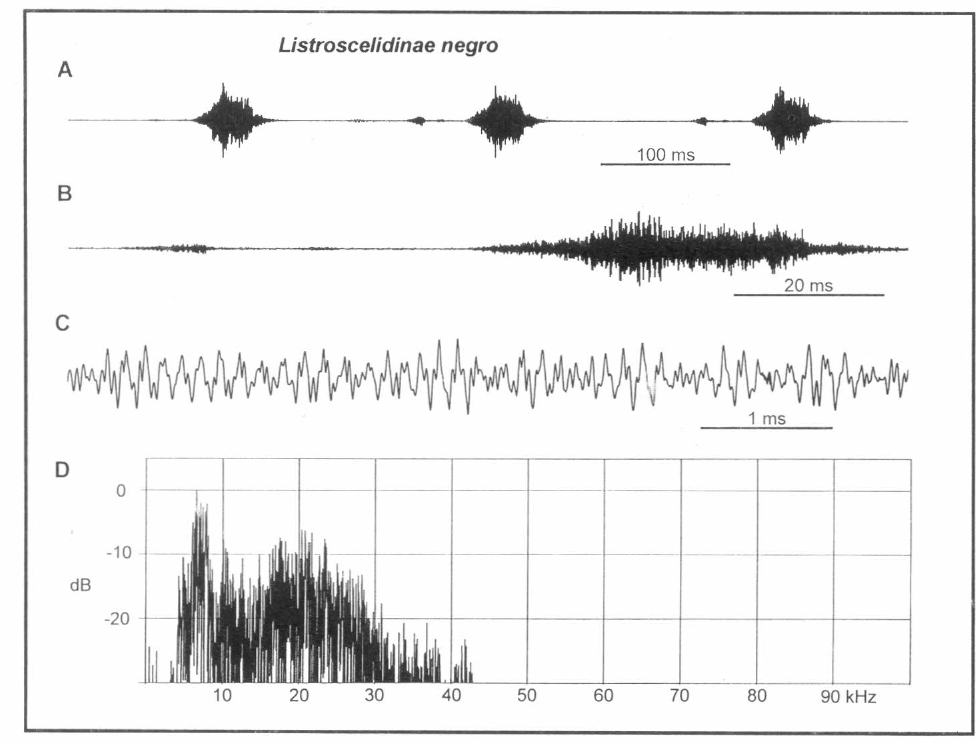
Análisis de canto de "Listroscelidinae negro". A. Tres fonótomos producidos durante un evento o ronda de emisión de sonido, B. Un fonótomo, C. Onda compleja del pulso mayor en A, D. Espectro de B.
De acuerdo con el ruido de mayor y menor pulso, la energía espectral está distribuida en una banda amplia. La parte más intensa es un pico coherente cerca a 7 kHz (6.53 kHz, Fig. 10D), pero hay también energía apreciable en una banda entre 4 y 29 kHz (usando <20 dB bajo el pico como criterio para energía biológicamente relevante).
Discusión
Los saltamontes machos que generan cantos lo hacen mediante el frotamiento de las alas anteriores o tegminas. El borde de una de estas alas (raspador) se mueve sobre una hilera de protuberancias o dientes (fila estriduladora) en el lado ventral de la otra. Las venas adyacentes de las membranas de la tegmina vibran cuando cada diente es frotado y como el cono de un parlante, estas membranas mueven el aire que las rodea para crear el sonido. El raspador viaja en un sentido a lo largo de la fila estriduladora cuando las tegminas deslizan entre sí, entonces el insecto debe cambiar la dirección y regresar al comienzo. De esta forma, para todas las especies, cada canto es una sucesión de movimientos de apertura y cierre de las alas anteriores.
Los cantos de saltamontes descritos aquí (excepto por 2 especies) son ruidosos, es decir, con muchas notas (frecuencias) diferentes. Esta diferencia en calidad surge de la velocidad a la cual el raspador hace contacto con los dientes de la fila estriduladora (Elsner y Popov 1978). Esto es análogo a empujar un niño en un columpio. Si se efectúan empujones con exacta regularidad, el niño se moverá hacia atrás y hacia delante a una sola frecuencia uniforme, pero si esta regularidad falla, dicha frecuencia uniforme decae y se pierde la sincronía del sistema. Algunos saltamontes producen cantos musicales sincronizando el raspador para contactar cada diente de la fila estriduladora con el fin de mantener las membranas de la tegmina vibrando suavemente hacia atrás y hacia adelante, es decir, a una frecuencia constante. Pero otras especies (especies ruidosas) hacen el contacto con los dientes de la fila mucho más lento y las membranas del ala pueden descansar en cada choque. En estas especies las membranas nunca se mueven en una forma sostenida, sino que en ellas el movimiento está constante y gradualmente iniciando y desapareciendo.
La estructura física del canto de las cinco especies de Pseudophyllinae encontradas en Ucumarí es amplia. El pico de frecuencias más alto lo presenta T. pilosula con 17.5 kHz, el más bajo Scopioricus spatulatus con 8 kHz. Sólo una especie tiene energía en el rango ultrasónico y tres especies producen espectros de banda: Chibchella nigrospecula, Stetharasa exarmata, y Scopioricus spatulatus; las demás tienden hacia espectros de tono puro altos, por ejemplo Trichotettix pilosula.
Se conocen los espectros de cuatro especies de Pterochrozini (saltamontes con forma de hoja seca) de estudios previos: Mimetica incisa Stål (Morris y Beier 1982) y tres especies del género Typlophylum (Morris et al. 1989; Montealegre y Morris 1999). Toda ellas tienen espectros de Q altos en combinación con pulsos sostenidos. La mayor duración de pulsos oscila entre 26 y 48 ms, y las frecuencias de 2.3 a 9.4 kHz. La especie de Typophyllum encontrada en Ucumarí se encuentra en este rango, el mayor pulso tardó 34.3 ms; la calidad del sonido generado es bastante musical (tono puro).
Hay una tendencia consistente en la morfología de la fila estriduladora asociada con la producción de pulsos sostenidos (tono puro) por estos insectos (Figs. 3A y 4). La vena de soporte Cu2 está fuertemente dilatada, elevando los dientes en un refuerzo alto y curvado. Los dientes son relativamente bajos, varias veces más anchos que altos y en gran número. En el extremo más cercano al cuerpo, la fila disminuye para formar una barra protectora separada de la tegmina encrustante (tegmina derecha), la cual puede servir para darle al raspador un alineamiento más recto a lo largo de la fila. La fila estriduladora de Trichotettix pilosula es un buen ejemplo. La morfología de esta fila estriduladora está relacionada aquí con la producción de pulsos sostenidos (alto-Q), esto ocurre también en otros Pseudophyllinae (por ejemplo, Scopiorinus, Morris y Beier 1982). Hay una cuarta uniformidad en las características físicas del canto de los Pterochrozini, los cantos son de tonos puros y se encuentran en el rango audible o escasamente alcanzan el rango ultrasónico.
Chibchella nigrospecula tiene un buen ejemplo de una típica vena estriduladora en los Tettigoniidae. La morfología de la fila estriduladora (Fig. 3B) comprende dientes relativamente delgados dentro de un refuerzo sencillo sin ninguna protrusión basal, es decir, la fila es menos curva. Este tipo de fila estriduladora está relacionado con la producción de cantos ruidosos (o espectros de banda ancha). Una característica inusual que resulta aquí es la tendencia de un movimiento (adoptivo) de ida y de venida, de duración comparable; Scopioricus spatulatus y Stetharasa exarmata muestran esta característica.
Se puede considerar que T. pilosula tiene un espectro de alto-Q, pero éste muestra sorprendentemente el rango de duración de pulso que con regularidad puede acompañar "cantores" con frecuencias simples. El canto de T. pilosula produce dos pulsos breves, igualados en tiempo. Pero al asumir que cada par de pulsos en T. pilosula es un fonótomo, el número de dientes es excesivamente grande (110) con relación al número total de ondas de amplitud sostenida o creciente, sumada sobre los dos pulsos. Probablemente dos pulsos no constituyen un fonótomo en esta especie aunque no se ha detectado algún ciclo en la estructura del canto, lo cual puede reflejar una transición en el ciclo de la tegmina.
Un macho de Chibchella nigrospecula fue observado alternando vibraciones corporales con llamadas. Se ha demostrado que la producción de ultrasonidos permite disminuir el rango en el cual un saltamontes macho es detectado por murciélagos que explotan la fuente sonora para localizar sus presas. La disminución de ultrasonidos llega a ser otra ventaja. Hay evidencia que algunas especies de murciélagos depredan sobre varias especies de saltamontes neotropicales (Belwood y Morris 1987) у muchas características estructurales y etológicas de estos insectos indican una historia de coevolución basada en una fuerte presión de depredación por parte de los murciélagos.
Las señales vibratorias son comunes en muchos saltamontes que exhiben ciclos de canto de duración reducida y altas frecuencias ultrasónicas. Un canto que es muy corto como para atraer a un murciélago sería muy corto también para atraer una hembra. De esta forma, la vibración de un substrato aporta un canal alterno de comunicación sexual en los saltamontes. Aunque C. nigrospecula no tiene cantos exclusivamente ultrasónicos, podría estar usando esta estrategia. Sin embargo, se puede especular muy poco respecto a las interacciones ecológicas entre estos animales ya que las especies de murciélagos insectívoros registradas en Ucumarí (Alonso y Cadena 1994), no corresponden a las que comúnmente se conocen como depredadores potenciales de saltamontes (Belwood 1988ab). Por lo tanto es recomendable realizar estudios más detallados en estos grupos.
Otro tipo de interacción ecológica interesante es el que exhiben las especies de Pterochrozini, las cuales mimetizan hojas secas. Poco se conoce de estas especies ya que es bastante difícil encontrar individuos en la vegetación (Fig. 2D). La especie encontrada en Ucumarí pertenece al género Typophyllum y además de su mimetismo crítico, exhibe un despliegue en el cual deja a la vista la coloración fluorescente en la membrana trocanto-coxal (en la base de las patas anteriores y medias) a manera de ojos (ver datos sobre la especie). Presumiblemente un par de ojos pueden ahuyentar a un depredador que se acerque al insecto por cualquier lado. Despliegues aposemáticos han sido observados en alas posteriores de otras especies de Pseudophyllinae (Robinson 1969, Castner y Nickle 1995), pero a nivel de membrana trocanto-coxal no se había registrado hasta ahora.
Es de esperar que el número de especies disminuya a medida que se incrementa la altitud y por supuesto ya se había notado este gradiente altitudinal en Tettigoniidae (Montealegre 1997). Sin embargo, no se realizó un estudio de este tipo en Ucumarí para investigar la gradación altitudinal referente a la abundancia de poblaciones.
A pesar que ocho especies es una riqueza bastante baja cuando se compara con cifras arrojadas en otros trabajos enfocados a otros grupos de insectos en Ucumarí (Andrade 1994; Schneider y Fernández 1994) es interesante que cerca del 90% de la fauna de saltamontes encontrada en el parque es nueva para la ciencia (la mayoría de especies de Pseudophyllinae fueron descritas recientemente [Montealegre y Morris 1999]).
Footnotes
Agradecimientos
Este estudio fue apoyado por Natural Science and Engineering Research Council of Canada, beca de investigación No. 4946 asignada a Glenn K. Morris. Los autores desean expresar sus más sinceros agradecimientos a la Corporación Autónoma Regional de Risaralda (CARDER), especialmente a Eduardo Londoño, por proporcionar alojamiento y permisos para trabajar en el PRN Ucumarí. La investigación en la Reserva Natural La Sirena (Palmira) fue apoyada completamente por la Alcaldía Municipal de Palmira, con agradecimientos al Dr. José Antonio Calle F. (exalcalde de la ciudad) y al Dr. Diego Arango (ex-asesor del gobierno municipal). La Corporación Autónoma Regional del Valle del Cauca, CVC, apoyó esta investigación proporcionando alojamiento y alimentación en las instalaciones de La Sirena y permiso de investigación, gracias a Jairo Arias. Fernando Vargas Salinas y Leonardo Rocha participaron en todas las jornadas de campo en La Sirena. Sinceros agradecimientos a Fernando Vargas por sus valiosas fotografias. Dita Klimas, Adrianne Rigler, Paul DeLuca y Liliana Castaño participaron en el trabajo de campo en mayo de 1996 en Ucumarí. Liliana Castaño ayudó en las jornadas de colecta en abril de 1997. Theodore Cohn y Piotr Naskrecki corroboraron la existencia de nuevos taxa. Se agradece inmensamente la ayuda de Wolfram Schulze y Voker Runkel (Universitat Erlangen, Alemania), por la edición de la figura 2 y a Maria Nakata (University of Toronto) por la edición de las micrografías electrónicas.