
Abstract
Communication by means of substrate-borne vibrations is a fundamental aspect of the reproductive behavior of most homopterans. This phenomenon has not yet been studied in the family Cercopidae. The role of vibrational communication in the reproductive behavior of Zulia carbonaria (Lallemand) and Aeneolamia varia (F.) was determined. Calls were registered directly from the substrate (stems and leaves of the plant) by means of a phonographic cartridge connected to a computer with specialized software for sound analysis. Trials were performed with males and females, together with observations during the recordings, to determine the relation between calls and associated behavior. The two sexes produced calls, but it was the male that initiated the search for the female. It was found that this form of communication was fundamental for mating and reproductive success in this family. To call the female, males produced a characteristic song; Z. carbonaria had a frequency of 317.38 ± 23.25 Hz and duration of 9.5 ± 0.59 sec., statistically different from A. varia which presented a frequency of 425.16 ± 37.27 Hz and duration 3.43 ± 0.08 sec. Upon detecting the male caller, the female answered, establishing a "dialogue" with the male which oriented him to find her in order to initiate mating. Other different patterns not directly related with mating were found. This is new basic information on the family Cercopidae and especially for grassland spittlebugs, the most important pest of neotropical forages. Considering the differences found in the calls of the two species, communication patterns could be used as a taxonomic tool, as has been used in other families.
Introducción
Desde el siglo XVIII la comunicación entre los insectos ha sido objeto de estudio en los órdenes Homoptera (Cicadidae) y Orthoptera (Tettigoniidae y Gryllidae) por ser fácilmente audibles sin el uso de equipos especiales. El caso más conocido es el de las cigarras (Homoptera: Cicadidae), cuyos cantos a través del aire pueden escucharse a varios kilómetros de distancia (Kirby y Spence 1818, Alexander 1960, citados por Claridge 1985). La mayoría de los insectos, sin embargo, producen sonidos no audibles para el oído humano. Requieren equipos especiales para poder ser registrados porque en muchos casos se transmiten a través del sustrato como piedras, arena, hojas, tallos e incluso la superficie del agua (Ewing 1989). Esta forma de comunicación se presenta en órdenes como Orthoptera, Blatodea, Isoptera, Plecoptera, Hemiptera, Homoptera, Neuroptera, Coleoptera, Diptera, Trichoptera e Hymenoptera (Stewart 1997).
En el suborden Auchenorrhyncha (Homoptera) se presentan los dos tipos de transmisión: por aire y por el sustrato. Estos cantos son producidos por un órgano situado dorsolateralmente en el primer segmento abdominal, conocido como timbal (uno a cada lado). A diferencia de la familia Cicadidae, los pequeños homópteros no poseen sacos de resonancia. Por esta razón sus cantos son de baja intensidad (Claridge 1985). Ossiannilsson (1949), pionero en realizar estudios sobre comunicación en pequeños homópteros, sugirió que para su comunicación son más importantes las vibraciones a través del sustrato. Propuso un sistema de clasificación de seis tipos diferentes de cantos: 1) canto común ("common song") se realiza de manera espontanea independiente de la presencia de otros insectos, 2) cortejo o apareamiento ("courtship") invita al apareamiento, 3) invitación ("invitation") es producida solo por hembras, 4) agresión ("rivalry") se presenta en caso de competencia por pareja, 5) cópula C'pairing") se realiza durante el apareamiento, y 6) conmoción o alarma C'distress") se produce en situaciones de inconformidad o peligro. Los cantos a través del sustrato alcanzan hasta un metro de distancia (Hunt y Nault 1991).
La comunicación vibracional ha sido una herramienta útil en taxonomía. Claridge y Reynolds (1973) encontraron diferencias en los cantos de cuatro especies del género Oncopsis (Homoptera: Cicadellidae) demostrando que funcionan como un mecanismo de aislamiento reproductivo. Claridge et al. (1985) en trabajos realizados con dos poblaciones alopátricas de Nilaparvata lugens (Stal) encontraron diferencias en. la frecuencia de repetición de pulsos (FRP) de sus cantos. Heady et al. (1986) realizaron un cladograma con diez especies del género Dalbulus basándose en las diferencias de los cantos comunes. Los resultados mostraron tres grupos principales que coincidieron con los tres grupos monofiléticos de la taxonomía obtenidos por morfología.
La comunicación vibracional nunca ha sido estudiada en la superfamilia Cercopoidea (Homoptera). Los estudios de Ossiannilsson (1949) y Moore (1961) probaron que las familias Cercopidae, Aphrophoridae y Clastopteridae son capaces de emitir sonidos. Sin embargo, no describieron en detalle las diferentes clases de cantos y no probaron que éstos se transmitieran a través del sus trato. Se desconoce completamente su ímportancía en el comportamiento reproductivo en la superfamilia Cercopoidea aunque para otras familias como CicadeIlidae y Delphacidae son una parte fundamental tanto en el apareamiento, como en el mecanismo de aislamiento reproductivo (lchikawa 1974, 1976, Heady et al. 1986, Hunt y Nault 1991).
En la familia Cercopidae existe un subgrupo formado por algunos géneros que son de importancia económica en cultivos de gramíneas como forrajes y caña de azúcar (Fewkes 1969, Lapointe et al. 1994). Este subgrupo se conoce como salivazo o mión de los pastos. La biología de estas especies es bastante desconocida y su taxonomía es muy confusa en la actualidad. El estudio de la comunicación vibracional y el comportamiento relacionado con esta forma de comunicación es importante para ampliar el conocimiento básico de su biología. Además, se abre la posibilidad de utilizar las características de los cantos como herramienta taxonómica.
El objetivo de este trabajo fue comprobar si se presentaba la comunicación a través del sustrato en la familia Cercopidae y determinar su importancia en el comportamiento reproductivo. Se desarrolló una metodología que permitiera grabar los cantos de los adultos directamente del sus trato bajo condiciones que facilitaran la observación al momento de realizar los registros, sin perturbar su comportamiento natural. Esta metodología se usó para obtener evidencia para diferentes clases de cantos en dos especies de salivazo de los pastos: Zulia carbonaria (Lallemand) y Aeneolamia varia (F). Se caracterizó en detalle el canto de apareamiento del macho por su relación directa con el comportamiento reproductivo. Finalmente, se comparó el canto y el comportamiento relacionado entre las dos especies con el fin de encontrar si habían diferencias suficientes para justificar el uso de la comunicación vibracional como herramienta taxonómica en esta familia.
Materiales y Métodos
Se utilizaron dos especies para estudiar la comunicación vibracional en la familia Cercopidae. Inicialmente se eligió a Z. carbonaria para el establecimiento de la metodología debido a que es común en los departamentos de Cauca y Valle, es muy dócil y además era la más grande de las especies disponibles. Esto facilitó su manejo bajo condiciones experimentales, tanto en el establecimiento de una cría, como en la realización de los registros. Para establecer la cría se utilizaron adultos traídos de Santander de Quilichao (Cauca), siguiendo la metodología de cría de A. varia descrita por Lapointe et al. (1994). Z. carbonaria fue erróneamente identificada y conocida en Colombia como Zulia colombiana, la cual es un sinónimo menor de Zulia pubescens (F.), otra especie presente en la misma zona geográfica (CIAT 1999).
Una vez que se estandarizó la metodología de adquisición se utilizó A. varia para registrar y comparar sus cantos con los de Z. carbonaria. En Colombia A. varia se presenta en los Llanos Orientales y del Piedemonte Amazóníco de Colombia. Para estos estudios los adultos se obtuvieron de una cría masiva establecida desde hace diez años en el CIAT,con renovaciones periódicas con insectos traídos del campo desde Meta y Caquetá.
Para los ensayos se utilizaron adultos de 1 - 2 días de edad para reducir fuentes de variación entre individuos como la edad, gravidez de las hembras y el período de precópula. Los adultos tenerales sexados fueron separados en potes con plantas de Brachiaria ruziziensis (Germain y Evrard) dentro de cilindros de acetato hasta el momento de realizar los registros.

Montaje para la adquisición de los registros: a) plano general del montaje, b) detalle de la posición de los insectos con relación al cartucho fonográfico.
Para todos los ensayos se utilizaron como sustrato tallos de B. ruziziensis de 25 - 32 cm de alto, con tres hojas. Este arreglo facilitó al insecto espacio suficiente para caminar, alimentarse e interactuar con otros individuos al tiempo que permitió una distancia máxima de 30 cm con el cartucho (Fig. 1). Este rango aseguró una buena transmisión de las vibraciones del insecto al cartucho. La base del tallo se mantuvo en un recipiente plástico (40 mi capacidad) con agua, tapado con un cilíndro de espuma. El tallo de la planta se introdujo por un corte en el centro de la espuma.
El cartucho se conectó mediante un cable blindado a un pre-amplificador (especificaciones no disponibles) construido especialmente para aumentar la señal antes de ser procesada. El cartucho se rodeó con una malla de acero y el resto del montaje con una caja de Faraday fabricada en malla de aluminio conectada a tierra en el amplificador para descargar las interferencias electromagnéticas (Hunt y Nault 1991). Para reducir vibraciones externas, el montaje se instaló sobre una lámina de polipropileno (Icopor) y ésta a su vez sobre una lámina de espuma. Posteriormente, la señal se almacenó en una computadora (Gateway GP6 450) con una tarjeta de sonido (Sound Blaster PCI 64V) que convertía la señal análoga en una digital para poder realizar los análisis correspondientes (Fig. 1).
Para conseguir evidencia para las diferentes clases de cantos, se utilizaron cinco combinaciones de individuos simulando un rango de situaciones naturales: un macho, dos machos, una hembra, dos hembras, y una pareja. Bajo estas circunstancias se hicieron observaciones visuales, junto con los registros, para obtener evidencia de las distintas clases de cantos. Los cantos se distinguieron por los patrones (sonogramas) distintos al ser graficados y por el comportamiento asociado del insecto. No se pretendió cuantificar la actividad bioacústica según especie, sexo u otras condiciones. Se incluyeron todos los registros interpretables para realizar los análisis. Se sometieron aproximadamente 30 adultos de Z. carbonaria y 15 de A. varia, representando 40 horas de grabación aproximadamente.
Se comparó el canto de apareamiento de los machos de Z. carbonaria y A. varia. Se tuvieron en cuenta los tres parámetros anteriormente descritos: duración, frecuencia y FRP. Las diferencias en los promedios de cada parámetro se detectaron mediante una prueba Tukey con α = 0.05.
Resultados
Se obtuvieron registros a través del sustrato para machos y hembras de Z. carbonaria y A. varia. Con base en los diferentes patrones asociados con su respectivo comportamiento, se obtuvo evidencia para cinco clases de cantos: alarma, común, apareamiento, invitación y otro desconocido.
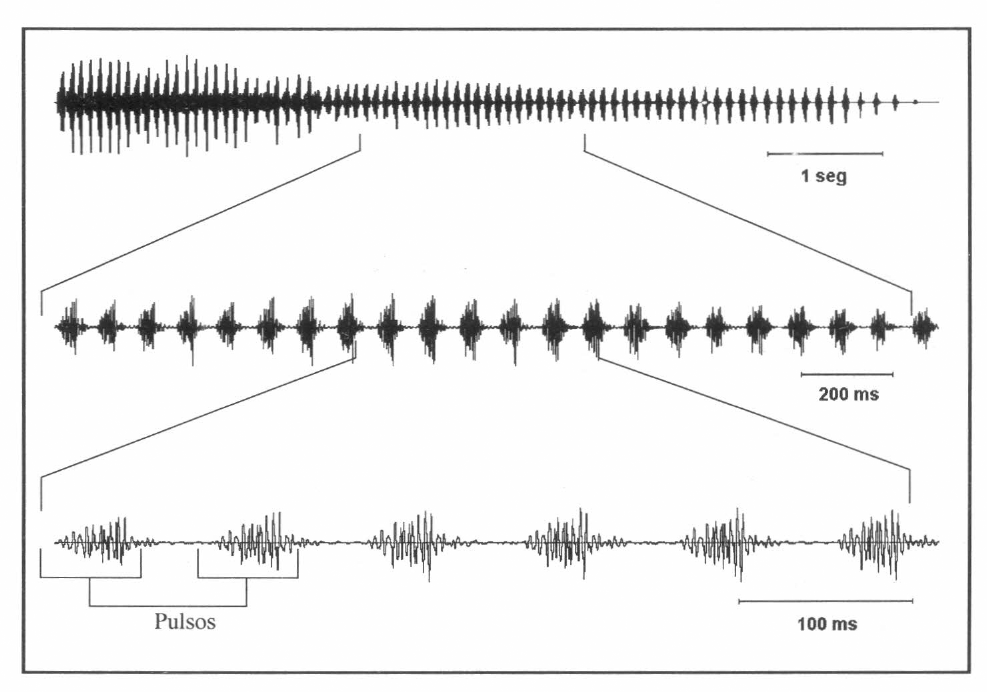
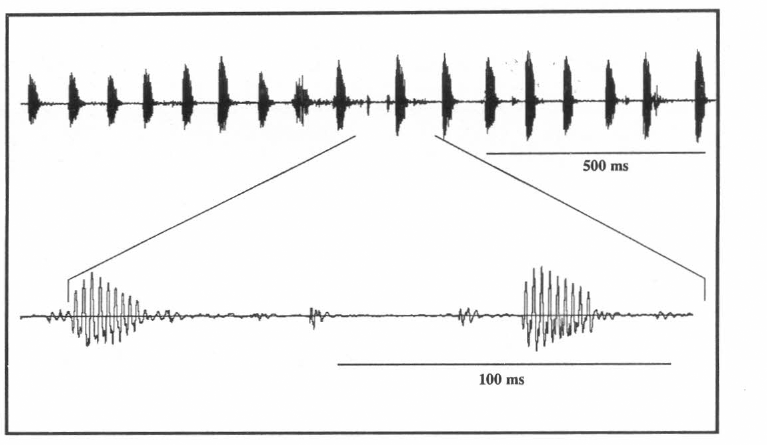
Oscilograma en tres escalas diferentes del canto de apareamiento o cortejo, realizado por el macho de Z. carbonaria para llamar a la hembra.
El canto de apareamiento de A. varia presentó una frecuencia de 425.16 ± 37.27 Hz (387.89 − 462.43), con una mayor intensidad entre 320 y 460 Hz, una duración promedio de 3.43 ± 0.08 seg, con una FRP de 42.16 ± 0.56 pulsos por seg (n = 5). La amplitud y el espacio entre pulsos fue constante desde el inicio hasta el fin (Figs. 3 y 4b).
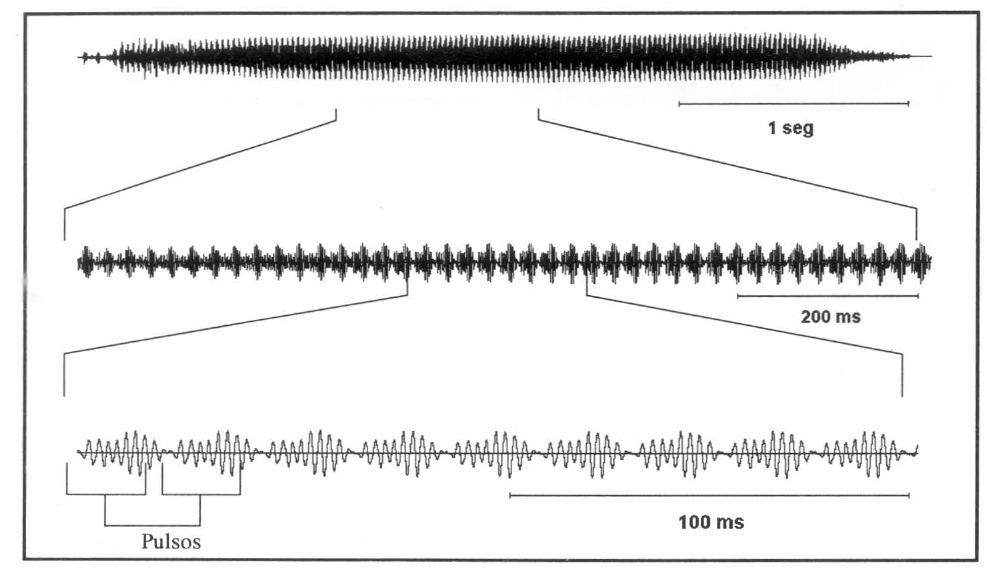
Oscilograma en tres escalas diferentes del canto de apareamiento o cortejo, realizado por el macho de A. varia para llamar a la hembra.
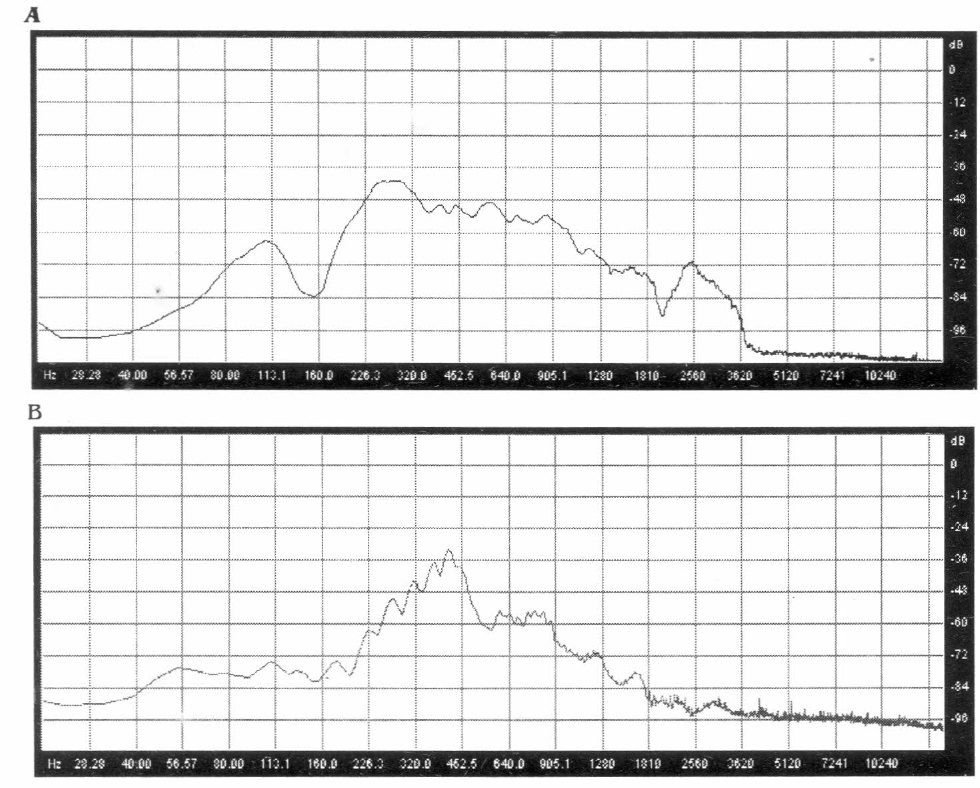
Análisis de espectro (frecuencia vs intensidad) del canto de apareamiento de los machos de las dos especies estudiadas.
El canto de apareamiento produjo como respuesta en las hembras de las dos especies el canto de invitación (Fig. 5). Las hembras solo hicieron el canto de invitación como respuesta al canto de apareamiento de los machos. Nunca se observó este canto con hembras solas. En el caso del canto de apareamiento siempre fue el macho quien empezaba a buscar a la hembra luego de que ésta contestara al canto de apareamiento con el canto de invitación. El macho se orientaba con el canto de invitación de la hembra hasta encontrarla; cuando los dos entraban en contacto físico, se iniciaba la cópula y terminaban los cantos. Los valores de frecuencia para este canto no se midieron dado que se registró mezclado con el canto de apareamiento de los machos.
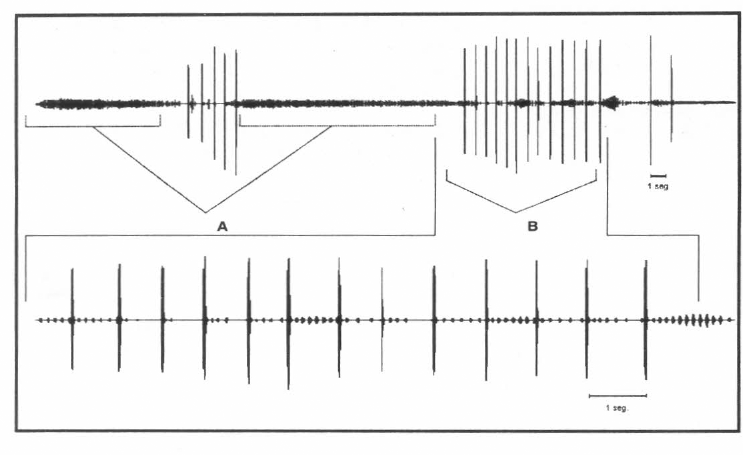
Oscilograma del intercambio de cantos entre un macho (canto de apareamiento A) y una hembra (canto de invitación B) de Z. carbonaria.
Oscilograma del canto de alarma de una hembra de Z. carbonaria.
El canto común era realizado espontáneamente tanto por hembras como por machos al caminar sobre los tallos y hojas de la planta hospedera, o cuando se alimentaba de ésta. En Z. carbonaria presentó una frecuencia de 322.64 ± 21.36 Hz (301.28 - 344.00) (n = 10 pulsos). En A. varia presentó una frecuencia de 405.33 ± 41.14 Hz (364.19 - 446.47) (n = 14 pulsos). No se observó que este canto condicionara una respuesta en otros insectos sobre la misma planta (Fig. 7).
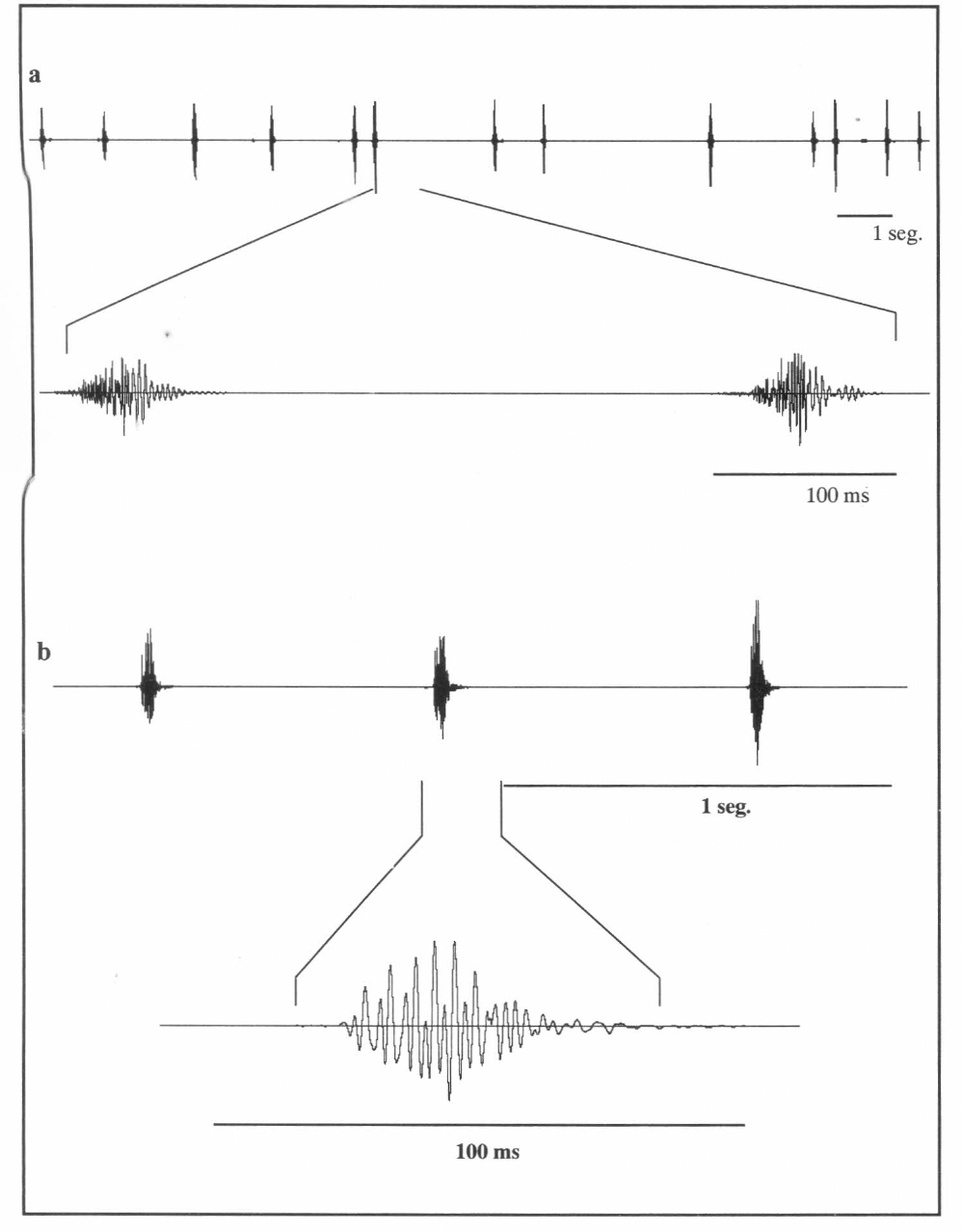
Oscilograma del canto común de: a) A. varia y b) Z. carbonaria.
Un canto distinto se observó cuando había dos machos de Z. carbonaria; éste se obtuvo dos veces durante 15 y 25 minutos continuos. Este canto consistió en pulsos de 461.50 ± 4.17 Hz (457.33 − 455.67) para el primero (n = 20 pulsos) y 472.05 ± 5.32 Hz (466.73 − 477.37) para el segundo. Este patrón se denominó comunicación entre machos (Fig. 8).
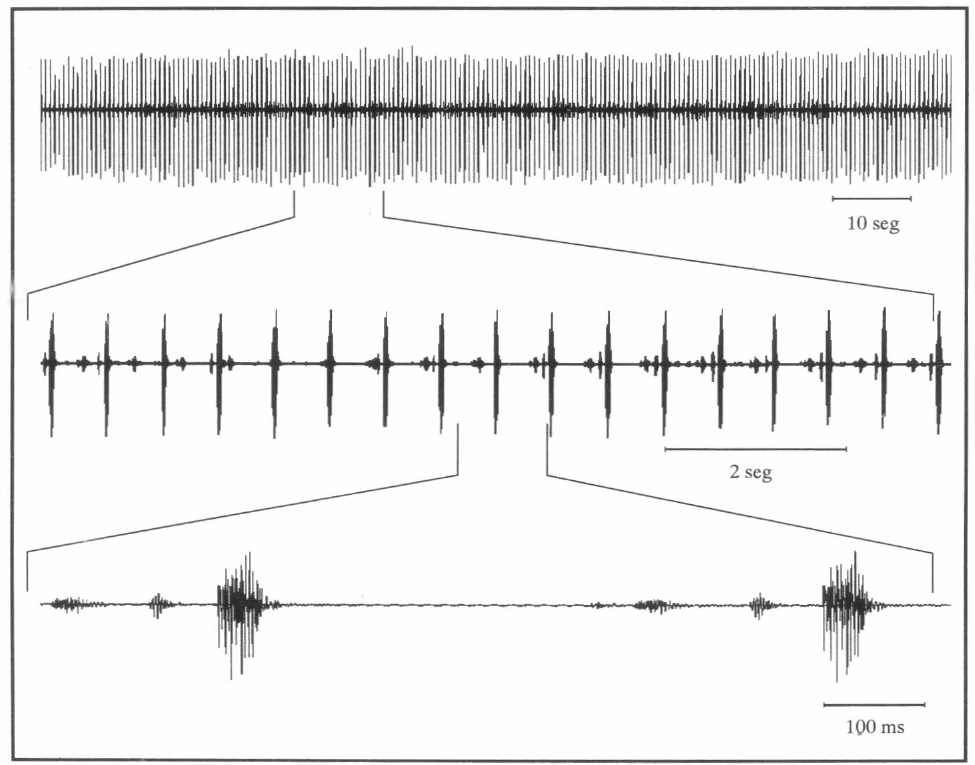
Oscilograma en tres escalas diferentes de la comunicación entre dos machos de Z. carbonaria. El de mayor amplitud corresponde al macho que estaba en el mismo tallo del cartucho; el de menor amplitud es de otro macho que estaba en un tallo adyacente. Las dos plantas tenían las hojas en contacto.
Diferencias entre especies. Al comparar los cantos de apareamiento de los machos de las dos especies, se encontraron diferencias significativas entre la frecuencia, duración del canto y la FRP (Tabla 1). Los oscilogramas mostraron dos cantos muy distintos en su forma. El canto de apareamiento de Z. carbonaria mostró una serie de pulsos espaciados que van perdiendo amplitud a medida que transcurre el canto (Fig. 2). El canto de apareamiento de A. varia presentó pulsos continuos con amplitud constante aminorando solo al inicio y al final del canto (Fig. 3). Las gráficas de análisis de espectro (intensidad vs frecuencia) de los cantos de las dos especies mostraron que la mayor intensidad (dB) se presenta en frecuencias diferentes para cada especie.
Diferentes parámetros evaluados (promedio ± error estándar*) a partir del canto de apareamiento registrado en los machos de Z. carbonaria y A. varia
Para cada parámetro valores seguidos con la misma letra no presentan diferencias estadísticas significativas según la prueba de Tukey α = 0.05.
Discusión
Se confirmó por primera vez que en la superfamilia Cercopoidea se presenta la comunicación por vibraciones a través del sustrato (tallos y hojas de las plantas hospederas). Igual que sucede en Cicadellidae (Claridge y Howse 1968, Heady et al. 1986), en Cercopidae los timbales, en hembras y machos, son los que producen las vibraciones que se transmiten por el sustrato. En Delphacidae sólo los machos poseen timbales funcionales; las hembras producen las vibraciones con movimientos rápidos del abdomen sin tocar el sustrato (Ichikawa 1974, 1976).
El menor tamaño de los insectos de estas familias sugiere que es más efectiva la comunicación por sólidos que por el aire. El sonido al transmitirse por el aire, un medio menos denso que los sólidos, pierde energía más rápidamente, limitando así el alcance de los cantos (Ewing 1989).
Para las dos especies bajo estudio, esta forma de comunicación es importante en el comportamiento reproductivo para el reconocimiento intraespecífico y para la ubicación de la pareja para el apareamiento. Siempre fue el macho el que inició la comunicación y búsqueda de la hembra. Esta se limita a responder los cantos del macho para orientarlo, permaneciendo inmóvil en su sitio de alimentación. Este comportamiento contrasta con el de otros homópteros como N. lugens (Homoptera: Delphacidae) donde la hembra es la que inicia la comunicación (Ichikawa 1974, 1976). Al ser cantos transmitidos por sólidos, los machos pueden determinar el origen y la dirección de donde provienen los cantos mas fácilmente que si el sonido fuera transmitido por el aire (Michelsen et al. 1982).
En algunas especies de importancia agrícola de las familias Cicadellidae y Delphacidae, se utilizaron los cantos para diferenciar especies e incluso híbridos resultantes de las mismas (Claridge y Howse 1968, Ichikawa 1974, 1976, Heady et al. 1986). En el caso de Z. carbonaria y A. varia las diferencias entre los cantos de apareamiento demuestran que eventualmente podría utilizarse como herramienta taxonómica, como por ejemplo para distinguir entre especies crípticas. Al igual que Claridge y Howse (1968), Ichikawa (1974, 1976) utilizó el canto de apareamiento para realizar las comparaciones entre las dos especies. Heady et al. (1986) utilizaron el canto común para realizar un cladograma con 11 especies del género Dalbulus. En el caso de los cercópidos el canto de apareamiento es mejor que el canto común para encontrar diferencias entre especies.
Los otros patrones encontrados demuestran que estos insectos tienen un repertorio variado de cantos para diferentes situaciones. Para su correcta descripción es necesario una investigación más detallada con situaciones diferentes a las utilizadas en este estudio. En el caso de la comunicación entre dos machos, no se observó un comportamiento que pudiera arrojar alguna información útil sobre la finalidad de esta canto. Siempre estuvieron quietos en su sitio de alimentación durante todo el tiempo que duró su comunicación.
Esta nueva información sobre la familia Cercopidae podría explicar algunos aspectos de su comportamiento, como parte del conocimiento básico que se debe tener de un insecto plaga. Hunt y Nault (1991) encontraron que los machos de Graminella nigrifrons (Forbes) (Homoptera: Cicadellidae) utilizan una estrategia llamada "call-fly" (llamar - volar) para buscar las hembras. El macho se posa sobre una hoja y llama de forma espontánea esperando la respuesta de una hembra cercana. Si no hay respuesta el macho vuela a otra planta. Con las observaciones realizadas en Z. carbonaria y A. varia se encontró un comportamiento similar por parte de los machos. Aunque ellos fueron puestos de forma manual sobre la planta, pasado un tiempo realizaban el canto de apareamiento esperando respuesta de alguna hembra. En el campo este comportamiento podría explicar porqué en algunos casos, en los muestreos realizados con pases de jama, se capturan más machos que hembras.
Conclusiones Y Recomendaciones
La metodología de adquisición de los registros directamente del sustrato demostró ser fácil y confiable; no es necesario someter los insectos a espacios reducidos que limiten o afecten su comportamiento natural. Esta metodología es fácilmente adaptable para otras familias de homópteros que tengan la misma forma de comunicación a través del sustrato.
Se comprobó que los insectos de la familia Cercopidae utilizan cantos transmitidos a través del sustrato para comunicación intraespecífica. Estos cantos se producen en el timbal, el cual es funcional tanto en machos como en hembras. Una función de este tipo de comunicación es la identificación y ubicación de pareja para el apareamiento. En Z. carbonaria y A. varia siempre fue el macho el que inició la comunicación y posterior búsqueda de la hembra.
Aunque las hembras contestaban el llamado del macho con el canto de invitación, nunca se movieron de su sitio de alimentación. Este canto de invitación es también el responsable de la orientación del macho hasta el sitio donde está situada la hembra.
Se encontraron otros patrones no relacionados directamente con el apareamiento, como el canto común, de alarma, lo que sugiere que existen otros cantos para diferentes situaciones. El canto denominado "comunicación entre machos" de Z. carbonaria se detectó solo entre machos. Lo realizaron independiente de la presencia o no de las hembras.
En el canto de apareamiento se encontraron diferencias significativas en términos de frecuencia, duración del canto y número de pulsos por segundo, entre las dos especies. Estas diferencias sugieren que podrían servir como herramienta taxonómica para identificación de especies.
La comunicación vibracional es un aspecto completamente nuevo para el estudio de la biología de la superfamilia Cercopoidea y abre un nuevo campo de investigación que podría explicar muchos aspectos desconocidos de la biología de este grupo de insectos. Dado que los cercópidos de importancia económica son insectos biológicamente desconocidos y difíciles de manejar, se justifica un estudio más detallado que permita caracterizar los cantos de otras especies de importancia económica para apoyar la taxonomía del grupo y comprender un poco su comportamiento en el campo.
Footnotes
Agradecimientos
Al Dr. Paul-Andre Calatayud por la colaboración brindada en la parte técnica, en la revisión de este artículo y del informe de tesis. Se agradece también a Ulises Castro, Guillermo Sotelo, Anuar Morales y Reynaldo Pareja por sus ideas, críticas y apoyo constante durante el desarrollo del trabajo. Este trabajo se realizó gracias a las facilidades del Proyecto de Gramíneas y Leguminosas Tropicales, CIAT.