
Abstract
The biology of three Colombian species of the genus Zulia (Homoptera: Cercopidae) was studied to establish comparative methodologies for the study of the grassland spittlebug complex, obtain new basic information and describe variation at the level of genus. To facilitate the differentiation and recognition of the developmental stages, certain aspects of the external morphology were described and characterized for the four developmental stages of the egg, five instars and adult sexes. To quantify the duration of the developmental stages, eggs were observed under incubation in petri dishes, nymphs in pots on surface roots of Brachiaria ruziziensis Germain and Everard, and adults on pots planted with B. ruziziensis maintained under acetate cages. Finally, oviposition sites were determined by simulating field conditions in the screenhouse in pots planted with B. ruziziensis. In the eggs of each species there was an increase in size among stages with little variation in external characteristics during development. In the nymphs there was an increase in size between each instar for each of the four parameters evaluated, with the head capsule being the most diagnostic morphological character. The adults presented sexual dimorphism expressed by the greater size of females in the majority of the six parameters evaluated in Z. carbonaria (Lallemand), Z. pubescens (Fabricius) and Zulia sp nov. The duration of the egg stage was 17.4, 14.3 and 14.6 days, respectively; the duration of the nymph stage 42.4, 38.0 and 42.7 days and adult longevity 19.6, 18.4 and 14.2 days, for a total life cycle of 69.6, 61.5 and 64.4 days, respectively. There were significant differences among species in developmental stages and total duration. Z. carbonaria and Zulia sp nov. presented a preference for the soil as oviposition substrate with 99.4 and 100% of recovered eggs, in contrast to Z. pubescens with 59.2% of eggs recovered from the stem surface and 40.4% from the soil. The established methodologies are adequate for describing the biology of Zulia and other spittlebug genera. This new biological information is important for improving the management of these species in forage grasses of Colombia. In addition, the degree of variation in the genus is relevant to the integrated management of spittlebugs and strengthens our understanding of the patterns of variation exhibited by this insect complex.
Introducción
El complejo del salivazo de los pastos (Homoptera: Cercopidae) es considerada la plaga limitante más importante en la producción ganadera de América Tropical (Lapointe y Miles, 1992), debido a su amplia distribución, a los altos niveles de población que alcanza en épocas de lluvia y a los daños que ocasiona en las pasturas susceptibles. En los últimos 10-20 años se ha convertido en plaga dañina en Colombia en los departamentos de Bolívar, Caquetá, Cesar, Córdoba, Magdalena, Meta, Santander, Sucre, Tolima y Valle del Cauca. Actualmente es una plaga creciente en gramíneas forrajeras en las laderas de Colombia al igual que en caña de azúcar en Centroamérica, Ecuador y Brasil (D. P., observación personal).
Los avances en el manejo del insecto se encuentran limitados por razones que incluyen (1) una persistencia a generalizar sobre las especies del complejo a pesar de que existe una alta diversidad taxonómica que implica una variación biológica, y (2) la falta de información básica sobre la biología y comportamiento de la mayoría de las especies económicamente importantes (Peck 2000a). Por ejemplo, de las 15 especies halladas en Colombia (Peck 2001) solo Aeneolamia varia (F.), Zulia carbonaria (Lallemand) (anteriormente Z. colombiana) y Prosapia simulans (Walker) han sido estudiadas biológica o ecológicamente. Se han iniciado estudios para caracterizar Aeneolamia reducta (Lallemand) y Aeneolamia lepidior (Fowler) en las sabanas colinadas de la Costa Caribe y Mahanarva sp nov. en las márgenes del bosque del Piedemonte amazónico de Colombia (CIAT 1998).
Para Colombia actualmente se encuentran referenciadas cuatro especies del género Zulia: Z. birubromaculata (Lallemand), Z. carbonaria, Z. pubescens (F.) y Zulia sp. nov. No se conoce Z. birubromaculata fuera de Colombia y ha sido señalada solamente en la costa Pacífica de Nariño y Valle del Cauca. Z. carbonaria se conoce en Bolivia y cinco departamentos colombianos, principalmente en los valles interandinos. Z. pubescens es frecuente en Perú, y Ecuador hasta Surinam, incluyendo 10 departamentos colombianos. Zulia sp. nov. es una nueva especie hallada en la Costa Pacífica de Ecuador y Nariño (Peck 2001). Fuera de un estudio biológico sobre Z. carbonaria (Arango y Calderón 1981) no existe información biológica en la literatura que haga referencia a estas especies.
Dentro de la poca información publicada sobre el género Zulia, Arango y Calderón (1981) determinaron que el macho adulto de Z. carbonaria mide 10.0 mm de longitud por 4.8 mm de ancho; la hembra es un poco más grande y mide 11.0 mm de longitud por 5.0 mm de ancho. Los huevos miden 0.97-1.17 mm de largo y 0.26-0.33 mm de ancho. Para las ninfas determinaron para cada uno de los ínstares su longitud, ancho del tórax, amplitud de la cápsula cefálica y amplitud del clipeo. Además, indicaron para Z. carbonaria un ciclo de vida de 72.0 días, dividido en 15 días para el huevo, 45 días para la ninfa y 12 días para el adulto.
Con el presente estudio se busca suministrar información básica sobre la biología de Z. carbonaria, Z. pubescens y Zulia sp. nov. para lo cual se emplearon metodologías utilizadas en estudios de otras especies (CIAT 1998; Peck 1998; Ballesteros y Gallego 1999). Esta información incluye (1) caracterización morfológica de los estados de desarrollo de los huevos, ninfas y adultos, (2) cuantificación de la duración de los estados de vida, y (3) descripción del uso y preferencia por los sitios de oviposición. Además, se esperó confirmar la aplicabilidad de estas metodologías para el estudio de otras especies de salivazo presentes en Colombia y en general en América Tropical. Finalmente, se pretendió describir la variación presente a nivel del género Zulia para poder comparar con otras especies colombianas y con otros géneros estudiados anteriormente. Se discute como esta nueva información fortalece patrones de variación dentro del complejo salivazo.
Materiales y Métodos
El presente trabajo se realizó en las instalaciones del Centro Internacional de Agricultura Tropical (CIAT), Palmira, Valle del Cauca, localizado a 3° 31′ N y 76° 22' W, altitud de 965 msnm y precipitación anual de 1000 mm. La zona de vida corresponde a Bosque Seco Tropical de Holdrige, con una temperatura promedia de 24°C.
Origen y manejo de insectos.
Z. carbonaria se recolectó en el departamento del Cauca a 1060 msnm (Mpio. Santander de Quilichao), Z. pubescens en el departamento del Cauca a 1650 msnm (Mpio. Popayán, estación CIAT) y Zulia sp nov. en el departamento de Nariño a 16 msnm (Mpio. Tumaco, C. I. El Mira de CORPOICA), único sitio del país donde se ha registrado esta especie. En cada una de estas regiones se recolectaron adultos para establecer una colonia en casa de malla superando la estacionalidad del insecto en el campo y contar con todos los estados de desarrollo del insecto requeridos por los ensayos.
Los insectos se mantuvieron en casa de malla según la metodología de Lapointe et al. (1989). La cámara de oviposición consistió de una jaula de madera (40 × 40 × 80 cm) con piso removible para introducir una capa de barro de 1.0 cm de espesor como sustrato de oviposición y aberturas laterales para el suministro del alimento constituido por plantas de Brachiaria ruziziensis Germain y Everard CIAT 654. Se obtuvieron los huevos del sustrato por una serie de tamizados, lavados y flotación en agua saturada con sal. Los huevos se desinfectaron con hipoclorito de sodio al 0.05% y se almacenaron en cajas de petri sobre papel filtro húmedo en condiciones controladas de incubación (27°C, 100% HR, oscuridad total). Ninfas de un día de eclosionadas se infestaron, en número de 20 por matera plantada, previamente con macollas de B. ruziziensis. Las materas se cubrieron con una tapa de aluminio pintada de blanco provista de un orificio para sacar el material vegetal, proporcionando la buena formación de raíces secundarias y una alta humedad relativa, condiciones necesarias para el desarrollo de las ninfas.
Caracterización morfológica.
Para facilitar la diferenciación y reconocimiento de los estados de desarrollo se describieron y caracterizaron aspectos morfológicos externos. Se midió el largo y ancho de los estados de desarrollo del huevo: S1, S2, S3 y S4 (Fewkes 1969; Wiedijk 1982). En las ninfas se midió el ancho de la cápsula cefálica, largo del estilete (punto distal del labro al punto distal del labio), largo del cuerpo y largo del muñón alar anterior en los cinco ínstares. Para aumentar la resolución se distinguió entre el instar Va y el Vb (próximo a transformarse en adulto por aparición de la corona de espinas negras del adulto en la metatibia), además del sexo del ínstar Vb. Para los adultos, se adicionaron los parámetros ancho del cuerpo, largo del cuerpo con o sin ala, y largo del ala anterior (Fig. 1). Para tomar las medidas se contó con un microscopio estereoscópico acondicionado con reglilla ocular. Se dispuso de huevos (n= 68-100) de hembras provenientes de la colonia, ninfas (n= 40) de la colonia o el campo, y adultos (n= 40) recolectados directamente del campo. Debido a la escasez de especímenes de Zulia sp nov. no se incluyeron las ninfas.

Puntos de referencia para la caracterización morfológica. (1) largo cuerpo con ala, (2) ancho cápsula cefálica, (3) ancho del cuerpo, (4) largo ala anterior, (5) largo cuerpo sin ala y (6) largo estilete. Foto corresponde a Prosapia simulans.
Ciclo de vida.
La determinación de la duración de la fase de huevo se realizó en el laboratorio bajo condiciones de incubación (27°C, 100% HR, oscuridad total). Se realizaron evaluaciones diarias para determinar el paso de una fase a otra. Basados en 100 individuos iniciales se determinó la duración promedio de cada una de las fases de desarrollo y duración total de la fase de nuevo.
Para establecer la duración de las ninfas y los adultos se replicaron condiciones de campo en casa de mallas empleando como hospedero plantas de B. ruziziensis (Fig. 2). En el caso de las ninfas cada matera se infestó con una sola ninfa recién eclosionada (<12 horas de edad). Para determinar el paso de un ínstar a otro se utilizó la observación directa de la exuvia. Se colectaron adultos tenerales (<12 horas de edad) de la colonia procurando manipular lo menos posible los insectos. Se colocaron cuatro individuos en cada matera previamente preparada con la planta huésped bajo un cilindro de acetato transparente, cubierto en la parte superior con un trozo de muselina sujeta mediante un caucho para evitar el escape de los individuos. Para facilitar la evaluación se adicionó una capa de arena blanca en la superficie del sustrato para localizar fácilmente los individuos.

Montaje empleado para determinar la duración de la fase ninfal (A), longevidad de adultos (B) y preferencia por sitio de oviposición con relación al tipo de substrato (C): tapa invertida para sostener suelo para oviposición (1) y superficie con suelo y 2.0 g de hojarasca (2) (Foto: F. López).
Basados en 40 individuos y observaciones diarias se estableció la longevidad promedio para ninfas y la longevidad media para sexos de los adultos.
Sitios de oviposición.
Se determinaron los sitios de oviposición simulando condiciones de campo en casa de malla en materas plantadas con B. ruziziensis. En la parte superior de las materas se colocó una tapa de aluminio invertida para sostener una capa de suelo mullido de 2.0 cm de espesor. La mitad de esta superficie de suelo preparado se cubrió con 2.0 g de hojarasca seca (porciones de 1-2 cm de largo de hojas y tallos de B. ruziziensis) (Fig. 2). A los 10 días se terminó el ensayo cuantificando el número de huevos colocados en cada uno de los cuatro sustratos: suelo bajo hojarasca, suelo descubierto, hojarasca y sobre la planta. Para recuperar los huevos depositados en el suelo se empleó la metodología de Lapointe et al. (1989). Los huevos depositados en la hojarasca se cuantificaron mediante observación directa, al igual que los depositados en o sobre la superficie del tallo.
Los datos registrados durante el ensayo se agruparon en valores mínimos, máximos y promedios con el error estándar como medida de variación. Todos los análisis estadísticos se realizaron utilizando el programa SAS y la prueba de comparación múltiple Tukey-Kramer HSD al P>0.05. Para el caso específico de la longevidad de los adultos se empleó la distribución de Weibull (Pinder et al. 1978) para establecer la longevidad media de la población y de los sexos.
Resultados
Biología de Z. carbonaria
Huevo.
Los huevos de Z. carbonaria pasaron por las cuatro fases de desarrollo plenamente distinguibles por su apariencia externa y tamaño. Los huevos recién ovipositados en la fase S1 fueron de color amarillo crema y no mostraron ninguna evidencia de desarrollo. Se determinó el paso a la fase S2 con la aparición de una mancha roja localizada debajo de la línea de eclosión ubicada aproximadamente a un cuarto de distancia del polo anterior al posterior. La fase S3 se inició con la apertura de la línea de eclosión, exponiendo la superficie negra del opérculo. Se evidenció un punto rojo debajo del opérculo. En la fase S4 finalmente se evidenció la presencia de un par de puntos rojos aproximadamente a un tercio de distancia del polo anterior al posterior, a los costados del corión, y otra pareja al lado de la línea de eclosión representando los ojos. El tamaño de los huevos, tanto ancho como largo, se incrementó significativamente entre cada una de las cuatro fases con la excepción del largo del huevo entre la fase S1 y S2 (Tabla 1).
Tamaño (mm) de los huevos del salivazo del género Zulia según fase de desarrollo (promedio ± error estándar y rango)
Para cada fila, promedios seguidos por letras diferentes son estadísticamente diferentes al P< 0.05. Las diferencias se determinaron con la prueba Tukey-Kramer HSD para comparaciones múltiples.
La duración promedio del estado de huevos fue de 17.4 días dividido en 5.9, 1.1, 3.7 y 6.8 días para cada una de las fases de desarrollo, que corresponden a una proporción de tiempo de 34.1, 6.1, 21.2 y 38.9%, respectivamente, de la duración total del huevo (Tabla 2). No se observaron huevos diapáusicos durante el ensayo ya que todos eclosionaron dentro de los 20 días, la duración máxima de la fase de huevo.
Duración (días) de los huevos del salivazo del género Zulia según fase de desarrollo (promedio ± error estándar y rango)
Para cada fila, promedios seguidos por letras diferentes son estadísticamente diferentes al P< 0.05. Las diferencias se determinaron con la prueba Tukey-Kramer HSD para comparaciones múltiples.
Ninfa.
En cada uno de los parámetros morfológicos evaluados hubo un incremento significativo en el tamaño de un ínstar a otro (Tabla 3). Además, el ínstar Vb superó al Va en cada uno de los parámetros, y las hembras del ínstar Vb superaron a los machos del mismo ínstar en el largo del cuerpo y del estilete. La duración promedio para la fase ninfal fue 42.4 días divididos en 7.5, 7.2, 6.4, 8.3 y 13.0 días para cada uno de los ínstares, respectivamente, correspondiendo a una proporción de 17.7, 17.0, 15.1, 19.6 y 30.7% de la duración total de la ninfa. La duración del V ínstar fue estadísticamente diferente a los demás ínstares, además el IV ínstar presentó una duración mayor que los ínstares I, II y III (Tabla 4).
Tamaño (mm) de los estados de vida ninfal y adulto de Z. carbonaria (promedio ± error estándar y rango) (n= 40)
Para cada columna, promedios seguidos por letras diferentes son estadísticamente diferentes al P< 0.05. Las diferencias se determinaron con la prueba Tukey-Kramer HSD para comparaciones múltiples.
Duración (días) de la fase ninfal del salivazo del género Zulia según instar (promedio ± error estándar y rango)
Para cada fila, promedios seguidos por letras diferentes son estadísticamente diferentes al P< 0.05. Las diferencias se determinaron con la prueba Tukey-Kramer HSD para comparaciones múltiples.
Adulto.
Con excepción del largo del estilete, las hembras superaron a los machos en cada uno de los parámetros morfológicos evaluados (Tabla 3). Con respecto a los sexos del ínstar Vb, los adultos presentaron un incremento en el ancho de la cápsula cefálica y largo del ala anterior pero una disminución en el largo del cuerpo y estilete. La longevidad media para adultos de Z. carbonaria fue 19.6 días, con una longevidad media para los machos de 18.4 días y para las hembras de 20.4 días, superando a los machos en 2.0 días.
Las hembras de Z. carbonaria presentaron poca flexibilidad por el tipo de sustrato para ovipositar, con una preferencia marcada por el suelo. El 72.7 y 26.7% de los huevos se recuperaron en el suelo descubierto y suelo cubierto, respectivamente. Sólo el 0.64% se recuperó de la hojarasca y no se recuperaron huevos del tejido (Fig. 3).
Biología de Z. pubescens
Huevo.
Con poca excepción, los huevos de Z. pubescens experimentaron los mismos cambios externos exhibidos por Z. carbonaria al pasar por las cuatro fases de desarrollo. A diferencia de Z. carbonaria, en la fase S2 los huevos de Z. pubescens no se evidenció la mancha roja localizada debajo de la línea de eclosión. El tamaño se incrementó significativamente entre cada una de las cuatro fases con la excepción del largo del huevo entre la fase S1 y S2 de igual manera que ocurrió en Z. carbonaria (Tabla 1).
La duración promedio de los huevos fue 14.3 días divididos en 4.1, 1.1, 4.7 y 4.4 días para cada una de las fases de desarrollo, que corresponden a una proporción de tiempo del 28.9, 7.3, 33.0 y 31.0%, respectivamente de la duración total del huevo (Tabla 2). No se observaron huevos diapáusicos durante el ensayo ya que todos eclosionaron dentro de los 16 días, la duración máxima para la especie.
Ninfa.
Para cada uno de los parámetros morfológicos evaluados, hubo un incremento significativo en el tamaño de los ínstares (Tabla 5). Además, el ínstar Vb superó al Va en la mayoría de los parámetros excepto en el largo del estilete, y las hembras del Vb superaron a los machos del mismo ínstar en el ancho de la cápsula cefálica y largo del cuerpo sin ala. La duración promedio para la fase ninfal fue 38.3 días divididos en 6.7, 6.3, 5.9, 7.1 y 12.3 días para cada uno de los instares, respectivamente, correspondiendo a una proporción de 17.5, 16.4, 15.4, 18.5 y 32.2% de la duración total de la ninfa. La duración del ínstar V fue significativamente más prolongada que la de los demás ínstares (Tabla 4).
Tamaño (mm) de los estados de vida ninfal y adulto de Z. pubescens (promedio ± error estándar y rango) (n= 40)
Para cada columna, promedios seguidos por letras diferentes son estadísticamente diferentes al P< 0.05. Las diferencias se determinaron con la prueba Tukey-Kramer HSD para comparaciones múltiples.
Adulto.
Las hembras superaron a los machos en el ancho de la cápsula cefálica, largo del cuerpo sin ala y largo del estilete mientras que los machos superaron a las hembras en el largo del ala anterior (Tabla 5). Con respecto a los sexos del ínstar Vb, los adultos exhibieron un incremento en el ancho de la cápsula cefálica y largo del ala anterior y una disminución en la longitud del cuerpo y estilete. La longevidad media para los adultos de Z. pubescens fue 18.4 días, con una longevidad media para los machos de 17.5 días y para las hembras de 19.4 días, superando a los machos en 1.9 días.
Las hembras de Z. pubescens presentaron flexibilidad por el tipo de sustrato para ovipositar, con una preferencia similar por el tejido y suelo con el 59.2 y 40.4% del total de los huevos recuperados, respectivamente. Los huevos sobre la planta se asociaron a la superficie del tallo, particularmente en el tercio inferior cubiertos con las porciones secas del tallo. El 7.26, 33.13 y 0.41% de los huevos se recuperaron del suelo bajo hojarasca, suelo descubierto y hojarasca, respectivamente (Fig. 3).
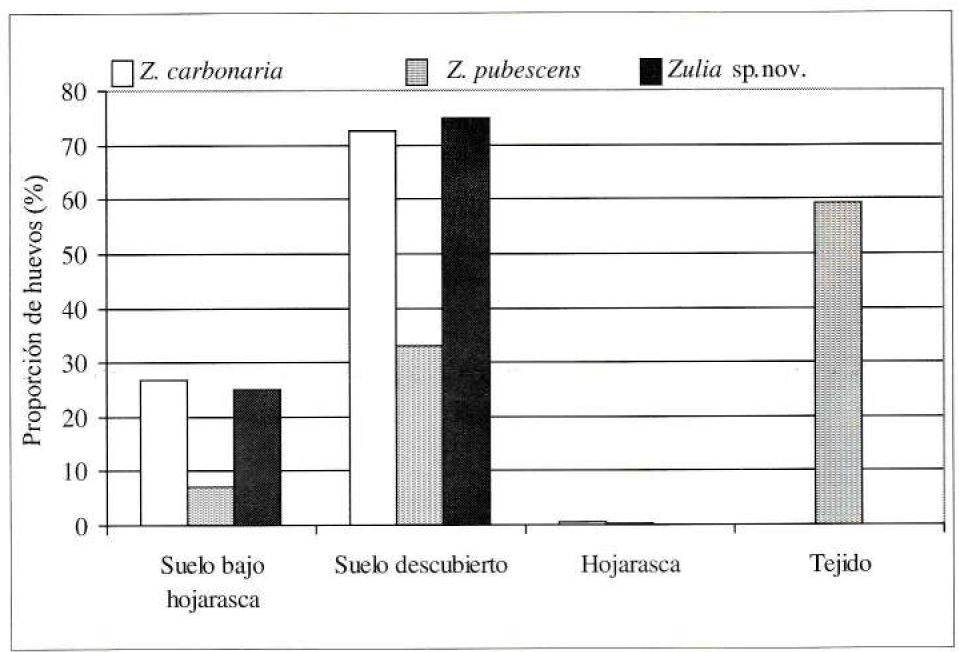
Preferencia y usos de sitios de oviposición por el género Zulia según cuatro sustratos. Un total de 472, 978 y 403 huevos se recuperaron para Z. carbonaria, Z. pubescens y Zulia sp. nov., respectivamente.
Biología de Zulia sp nov.
Huevo.
Los huevos de Zulia sp nov. experimentaron los mismos cambios externos exhibidos por Z. carbonaria y Z. pubescens al pasar por las cuatro fases de desarrollo. A diferencia de Z. carbonaria y Z. pubescens el huevo de Zulia sp nov. durante la fase S1 evidenció una mancha roja tenue en el extremo de la línea de eclosión. El tamaño de los huevos, tanto ancho como largo, se incrementó significativamente entre cada una de las cuatro fases con la excepción del largo del huevo entre la fase S2 y S3 (Tabla 1).
La duración promedio para los huevos fue 14.6 días divididos en 6.1, 1.1, 2.9 y 4.7 días para cada una de las fases de desarrollo, que corresponden a una proporción de tiempo del 41.9, 7.5, 20.0 y 32.2%, respectivamente, de la duración total del huevo (Tabla 2). No se observaron huevos diapáusicos durante el ensayo ya que todos eclosionaron dentro de 16 días, la duración máxima para la especie.
Ninfa.
La duración promedio para la fase ninfal fue de 42.7 días divididos en 8.0, 7.1, 6.7, 7.8 y 13.1 días para los ínstares I, II, III, IV y V, respectivamente, correspondiendo a una proporción del 18.7, 16.6, 15.7, 18.3 y 30.7% de la duración total de la ninfa. La duración del ínstar V fue significativamente más prolongada que la de los otros ínstares (Tabla 4).
Adulto.
Con excepción del largo del estilete las hembras superaron a los machos en cada uno de los parámetros morfológicos (Tabla 6). La longevidad media para los adultos de Z. pubescens fue 14.1 días, con una longevidad media para los machos de 12.9 días y para las hembras de 14.9 días, superando a los machos en 2.0 días.
Tamaño (mm) de los adultos de Zulia sp nov. según sexo (promedio ± error estándar y rango) (n= 40)
Para cada columna, promedios seguidos por letras diferentes son estadísticamente diferentes al P< 0.05. Las diferencias se determinaron con la prueba Tukey.
Las hembras de Zulia sp nov. no presentaron flexibilidad por el tipo de sustrato para ovipositar, con una preferencia por el suelo donde se recuperaron el 100% de los huevos. El 74.9 y 25.1% de los huevos se recuperaron del suelo descubierto y bajo hojarasca, respectivamente (Fig. 3).
Biología comparada de las especies
Huevo.
Desde el punto de vista de los cambios físicos externos que acompañan el desarrollo, hubo muy poca variación entre las tres especies. Todas presentaron un desarrollo que correspondió a las cuatro fases generalizadas de desarrollo. En términos de tamaño, todas exhibieron un incremento significativo de una fase a otra. Para las tres especies el rango del incremento entre S1 y S4 fue 21.4-26.7% para el ancho y 5.3-5.7% para el largo. En términos de la duración, la fase S2 fue la más corta para las tres especies, pero hubo diferencias en la fase más prolongada: S4, S3 y S1 para Z. carbonaria, Z. pubescens y Zulia sp nov., respectivamente. La duración total fue estadísticamente diferente entre cada una de las especies siendo Z. carbonaria la de mayor duración y Z. pubescens la de menor duración, variando 3.1 días (P<0.05, Tukey-Kramer HSD para comparaciones múltiples). Los huevos de Z. carbonaria duraron 21.6% más que los huevos de Z. pubescens.
Ninfa.
En términos de tamaño, sin excepción, todos los parámetros morfológicos evaluados para Z. carbonaria y Z. pubescens incrementaron entre los ínstares I al V. Además, el Vb superó al Va en cada parámetro con la excepción del largo del estilete en Z. pubescens (Tablas 3 y 5). Con excepción del largo del estilete, los sexos del ínstar Vb de Z. carbonaria superaron a los sexos del mismo ínstar de Z. pubescens en cada uno de los parámetros evaluados (P<0.05, Prueba Tukey).
En términos de duración, el ínstar V fue el más prolongado para las tres especies. Hubo diferencias entre todas las especies para la duración total del estado ninfal (P<0.05, Tukey-Kramer HSD para comparaciones múltiples). La duración total varió 4.7 días, con una duración de las ninfas de Zulia sp. nov. 12.4% más que las ninfas de Z. pubescens.
Adulto.
En términos de tamaño, para los adultos de Z. carbonaria y Z. pubescens el ancho de la cápsula cefálica superó el tamaño del ínstar V pero el largo del estilete fue menor. Hubo un polimorfismo sexual en las tres especies expresado por el tamaño mayor de las hembras en la mayoría de los parámetros evaluados. Los machos superaron a las hembras solamente para el caso del ala anterior en Z. pubescens. Para cada uno de los parámetros evaluados, los sexos de Z. carbonaria superaron en tamaño los sexos de las otras dos especies (P<0.05, Tukey-Kramer HSD para comparaciones múltiples). Además, las hembras de Zulia sp. nov. superaron a los sexos de Z. pubescens en cada uno de los parámetros estudiados. Las hembras de Z. carbonaria (más grande) superaron al estado más pequeño por 29.3, 26.8, 29.7, 45.7, 28.3 y 39.6% para el ancho de la cápsula cefálica, largo del estilete, largo del cuerpo con ala, el largo del cuerpo sin ala, ancho del cuerpo y el largo del ala, respectivamente (Tablas 3, 5 y 6).
En términos de longevidad, las hembras superaron a los machos de las tres especies evaluadas pero sin mostrar diferencia estadística; esta fue solo de 1.9-2.0 días. No se detectaron diferencias significativas entre los seis grupos (sexo x especie) con la excepción de los machos de Zulia sp nov. que presentaron una longevidad estadísticamente menor. La longevidad varió 7.5 días con las hembras de Z. carbonaria superando los machos de Zulia sp. nov. por 58.2%.
Ciclo de vida.
Las especies del género Zulia estudiadas presentaron un ciclo de vida que osciló entre 61.5 y 69.6 días, tomándose para este cálculo la mitad del valor de la longevidad de los adultos (Tabla 7). Varió 7.9 días con Z. carbonaria superando a Zulia sp. nov. por 38.5%. Para las tres especies la mayor proporción de la duración del ciclo de vida se expresó en la fase ninfal con un 60.9, 61.8 y 66.3% para Z. carbonaria, Z. pubescens y Zulia sp. nov., respectivamente.
Ciclo de vida de algunas especies colombianas de salivazo
Calculado por la mitad de la longevidad media de los adultos.
Sitios de oviposición.
Hubo diferencias marcadas entre las especies por el uso y preferencia por los sitios de oviposición. Z. carbonaria y Zulia sp. nov. exhibieron muy poca flexibilidad en el tipo de sustrato ya que solamente el 0.6% de los huevos se recuperaron fuera del suelo. Z. pubescens mostró mayor flexibilidad en el uso de los sustratos utilizando suelo (40.4%) al igual que la superficie del tallo (59.2%). Ninguna de las especies exhibió preferencia por la hojarasca.
Discusión
Los parámetros morfológicos evaluados en este estudio son adecuados para la determinación precisa de los estados de desarrollo de los huevos, ninfas y adultos del salivazo de los pastos. Esto permite una alta resolución para estudios de campo sobre fenología y un aporte para estudios biológicos. La mayoría de los estudios poblacionales, por ejemplo, no ha evaluado el ínstar de las ninfas, perdiendo así el detalle útil para la interpretación de la fluctuación poblacional y pronóstico en campo (Peck 1999).
Las metodologías empleadas para establecer la duración de los estados de vida son adecuadas para estas tres especies al igual que en otras especies de Aeneolamia y Mahanarva donde ya fueron empleadas (Tabla 7). Permiten la cuantificación de la duración de las fases de desarrollo de huevo e ínstar ninfal, aspectos que no se determinaron con metodologías empleadas en estudios anteriores con Z. carbonaria (Arango y Calderón 1981).
Los resultados obtenidos en este trabajo no incluyeron datos sobre aspectos de la biología reproductiva como períodos de precópula y preoviposición, tasa de oviposición y fecundidad, aspectos importantes en la descripción de la biología de la especie y su interpretación como plaga. Las metodologías probadas hasta la fecha por el CIAT no se adecuaron para las especies del género Zulia estudiadas en el trabajo.
Los resultados obtenidos para estas tres especies del género Zulia fortalecen ciertos patrones morfológicos comunes a otras especies del salivazo estudiadas hasta la fecha. Primero, los huevos pasan por las cuatro fases de desarrollo, incrementando en tamaño junto con el desarrollo, especialmente entre las fases S2 y S3 cuando abre el corión y el huevo absorbe agua. El mismo patrón está exhibido por las especies A. lepidior, A. varia, Mahanarva sp. nov., P. simulans, y Prosapia sp. nov. (CIAT 1998; Peck 1998; Ballesteros y Gallego 1999; Rodríguez et al. 2000). Segundo, las ninfas pasan por cinco ínstares plenamente distinguibles por el incremento significativo en el tamaño de parámetros físicos que acompañan la muda. El ancho de la cápsula cefálica es la característica morfológica más diagnóstica para separar los instares ya que no presentó traslape entre los rangos obtenidos para cada instar, lo cual es diferente a los demás parámetros evaluados. Se confirmó que se puede distinguir entre el ínstar Va y el Vb (próximo a transformar en adulto). En el ínstar Vb, además de la espina lateral y la corona de espinas en la metatibia de la ninfa, asociadas a éstas se evidencian las espinas y coronas oscuras del adulto próximo a salir. Tercero, se exhibe un polimorfismo sexual expresado en el tamaño mayor de las hembras adultas. Este patrón es aportado por otros géneros como Aeneolamia (CIAT 1998), Mahanarva (Ballesteros y Gallego 1999) y Prosapia (Peck 1998; Rodríguez et al. 2000). Cuarto, hay una reducción en el largo del estilete al pasar del ínstar V al adulto, comparable con las especies A. lepidior, A. reducta, Mahanarva sp. nov. y Prosapia sp. nov. (CIAT 1998; Peck 1998; Ballesteros y Gallego 1999). Finalmente un patrón biológico que se pudo establecer, es la menor duración de la fase S2, aportado por las especies A. reducta, Mahanarva sp. nov. y P. simulans (CIAT 1998; Ballesteros y Gallego 1999; Rodríguez et al. 2000). S2 es la fase que se extiende en el caso de huevos diapaúsicos.
Para la duración de los huevos, el rango exhibido por el género Zulia (14.3-17.4 días) es parecido a lo conocido por otros géneros como Aeneolamia (12.5-17.2) y Prosapia (12.0-18.7). Para las ninfas de Zulia, el rango exhibido (38.0-42.7 días) es intermedio a Aeneolamia (23.0-44.0) y Prosapia (23.0-62.0). Para los adultos, el rango exhibido (14.2-19.6 días) es también intermedio a Aeneolamia (6.4-24.5 días) y Prosapia (12.5-42.0 días) (Peck 2000b). Para la duración del ciclo de vida, el rango exhibido por las especies de Zulia (61.5-69.6 días) es superior al de otras especies de Aeneolamia (45.3-52.6) en Colombia, y con un rango similar al de Mahanarva sp. nov. y P. simulans (Tabla 7).
Aunque durante los ensayos ninguna de las tres especies del género Zulia exhibió evidencia de diapausa, se espera que en condiciones de campo ésta se exprese como un mecanismo del insecto para superar períodos secos poco favorables para su desarrollo.
Existe una variación intra e interespecífica con relación a los sitios de oviposición en los géneros de salivazo que se han estudiado (Peck 2000b). Z. carbonaria y Zulia sp. nov. mostraron preferencia por el suelo descubierto como sustrato de oviposición similar o lo referido en la literatura para el género Aeneolamia (CIAT 1998), y las especies P. simulans (Rodríguez et al. 2000) y Mahanarva sp. nov. (Ballesteros y Gallego 1999). Sólo Z. pubescens presentó preferencia por el tallo como sustrato de oviposición, como algunas especies del género Prosapia (Peck 2000b).
La nueva información obtenida para el género Zulia en comparación con otros géneros estudiados indica que existe más variación biológica dentro del género que entre géneros. Sin embargo, hay necesidad de realizar más estudios en otras especies del complejo para fortalecer el conocimiento de los patrones de variación en el salivazo de los pastos.
La determinación precisa de los estados de vida, variación en tamaño y duración del ciclo de vida es relevante en la determinación del impacto y manejo del insecto. Estos factores son importantes para el nivel de daño al hospedero y el período del insecto en el campo. Además, aumenta la resolución en la interpretación de muestreos y estudios sobre dinámica poblacional, tanto como la aplicación y efectividad de las tácticas de manejo. Por esta razón es necesario obtener una determinación taxonómica precisa para las especies presentes en cada región donde se requiera la implementación de un manejo. Es crítico realizar estudios biológicos de las especies de importancia económica que aún están desconocidas.
Conclusiones
La metodología evaluada es adecuada para describir la biología general de Zulia y otros géneros del salivazo; sin embargo, es necesario establecer una metodología para describir componentes particulares de la biología reproductiva como período de precópula y preoviposición, tasa de oviposición y fecundidad.
Es posible distinguir con precisión los estados de desarrollo (cuatro fases de desarrollo en los huevos, cinco ínstares ninfales y dos sexos en los adultos) basados en características morfológicas. En el caso de las ninfas el ancho de la cápsula cefálica es el carácter más diagnostico para diferenciar los ínstares.
Para las tres especies, el dimorfismo sexual se manifiesta en un tamaño mayor de las hembras para cada uno de los parámetros morfológicos evaluados.
Para el género Zulia, la duración total de la fase de huevo, ninfa y adulto es 14.3-17.4 días, 38.2-42.7 y 14.2-19.6 días, respectivamente, para un ciclo de vida 61.5-69.4 días.
Para el género Zulia, dos especies (Z. carbonaria y Zulia sp. nov.) exhibieron poca flexibilidad por el tipo de sustrato colocando el 99.4 y 100% de los huevos en el suelo, mientras que una especie (Z. pubescens) utilizó dos sustratos colocando el 40.4 y 59.2% de los huevos en el suelo y en la superficie del tallo, respectivamente. Ninguna especie mostró preferencia por la hojarasca.
La nueva información obtenida para el género Zulia sugiere que existe más variación biológica dentro del género que entre géneros.
Footnotes
Agradecimientos
Se agradece a todos los integrantes del grupo de Bioecología y MIP del Salivazo en el CIAT: Ulíses Castro, Francisco López, Anuar Morales y Rosalba Tobón por sus ideas y apoyo constante durante el desarrollo del trabajo. De igual manera se agradece a Reynaldo Pareja, Gilberto Córdoba y Guillermo Sotelo por su apoyo y colaboración para el desarrollo de este proyecto. Este trabajo se realizó gracias a las facilidades prestadas por el Proyecto IP-5 Gramíneas y Leguminosas Tropicales, CIAT y ayuda financiera parcial del Programa Nacional de Transferencia de Tecnología Agropecuaria (PRONATTA).